Phylogeny and Density Dynamics of Wolbachia Infection of the Health Pest Paederus fuscipes Curtis (Coleoptera: Staphylinidae)


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and DNA Extraction
2.2. Wolbachia Screening and Multilocus Sequence Typing
2.3. qPCR and Statistical Analyses
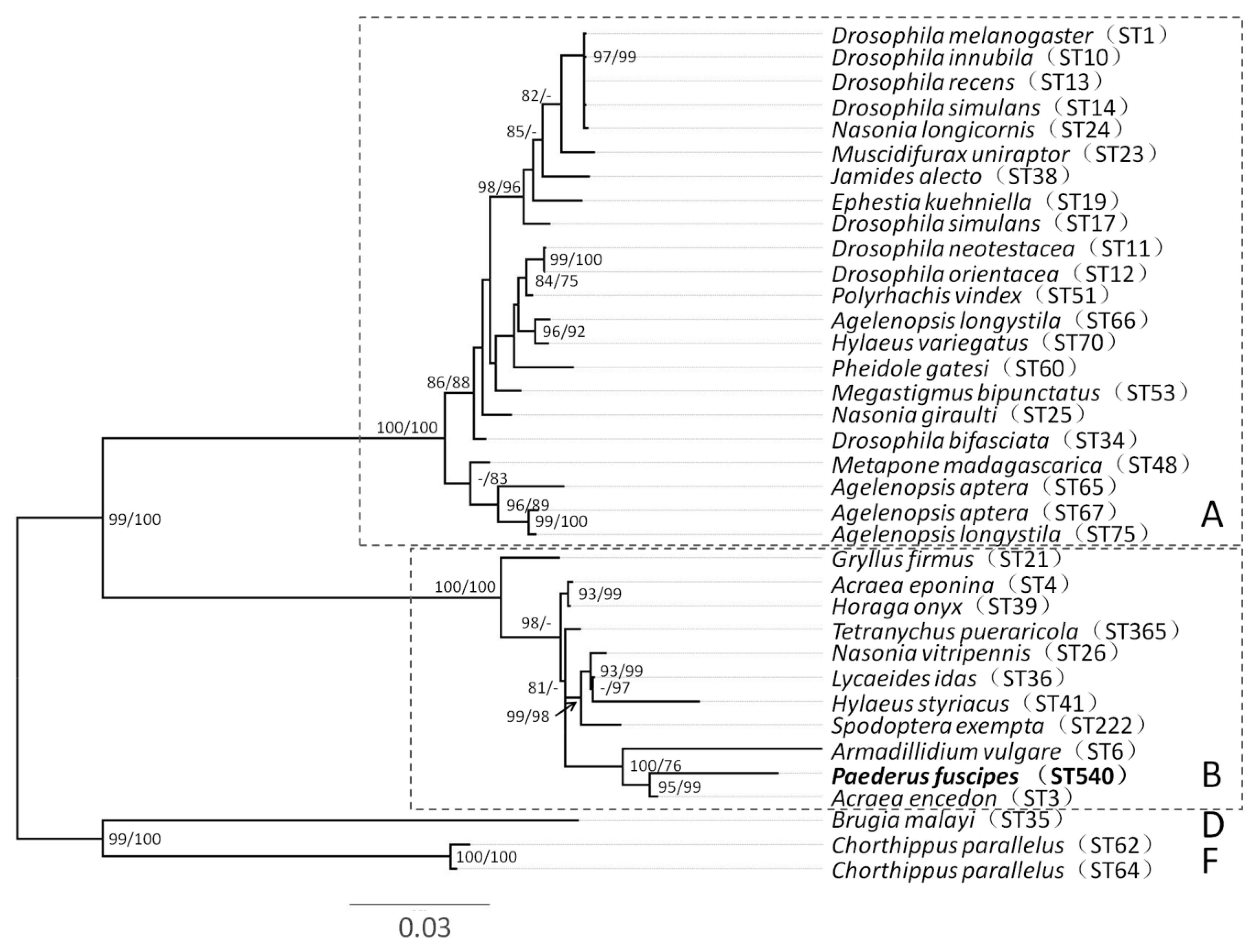
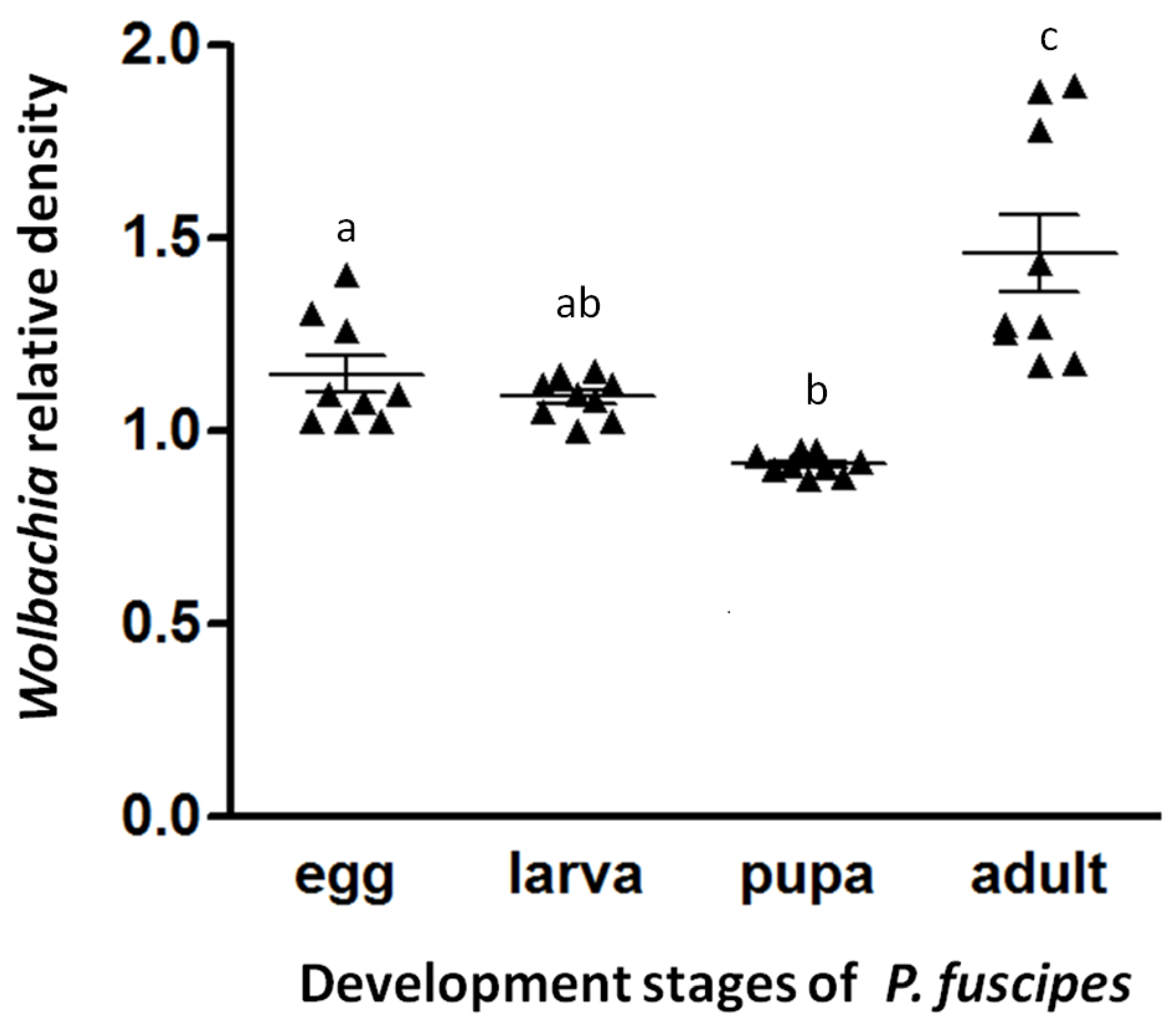
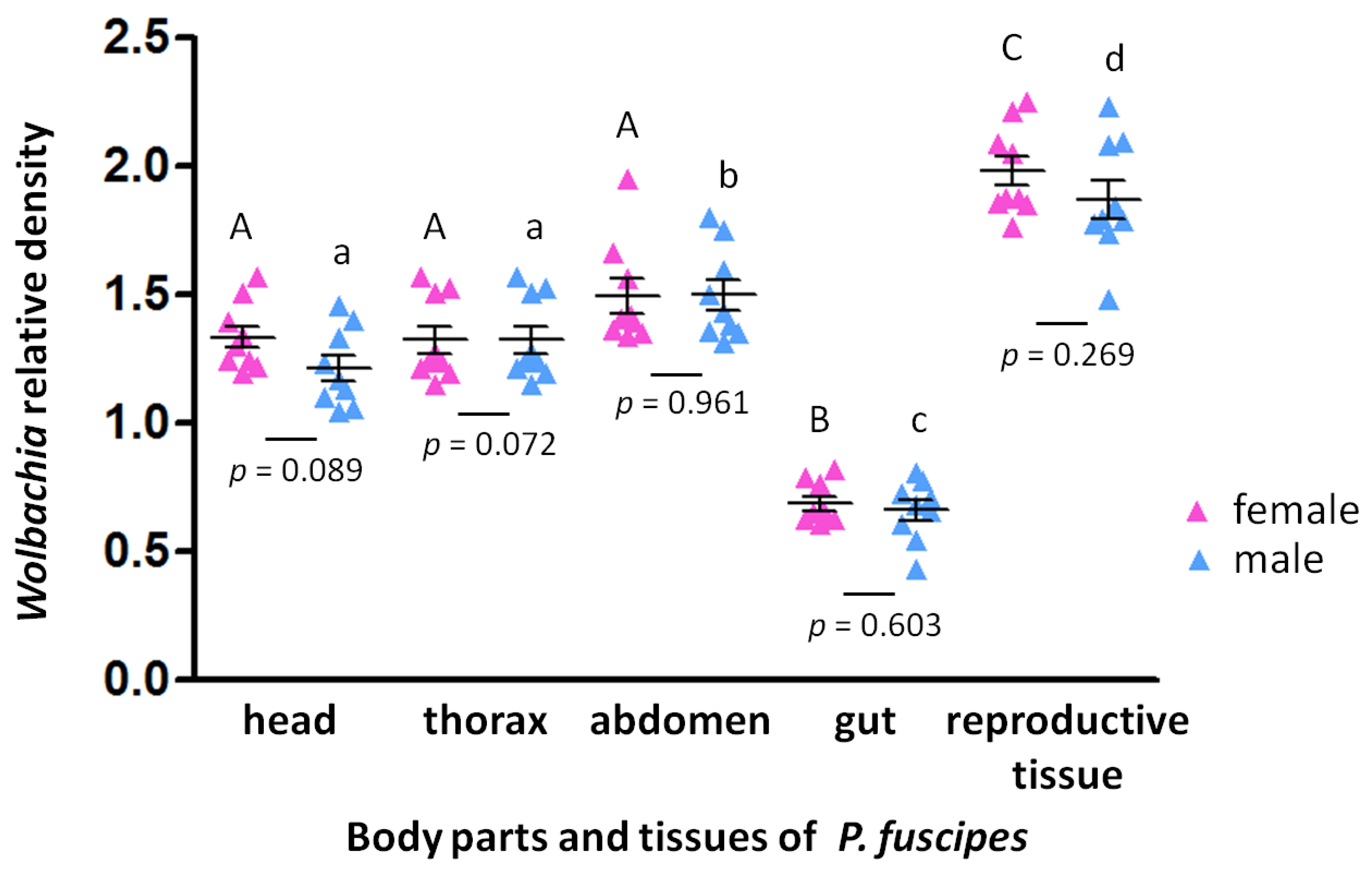
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef] [Green Version]
- Raychoudhury, R.; Baldo, L.; Oliveira, D.C.S.G.; Werren, J.H. Modes of acquisition of Wolbachia: Horizontal transfer, hybrid introgression, and codivergence in the Nasonia species complex. Evolution 2009, 63, 165–183. [Google Scholar] [CrossRef]
- Yen, J.H.; Barr, A.R. New hypothesis of the cause of cytoplasmic incompatibility in Culex pipiens L. Nature 1971, 232, 657–658. [Google Scholar] [CrossRef]
- Stouthamer, R.; Breeuwert, J.A.; Luck, R.F.; Werren, J.H. Molecular identification of microorganisms associated with parthenogenesis. Nature 1993, 361, 66–68. [Google Scholar] [CrossRef]
- Rigaud, T.; Souty-Grosset, C.; Raimond, R.; Mocquard, J.P.; Juchault, P. Feminizing endocytobiosis in the terrestrial crustacean Armadillidium vulgare Latr. (Isopoda): Recent acquisi-tions. Endocytobiosis Cell Res. 1991, 7, 259–273. [Google Scholar]
- Hurst, G.D.D.; Jiggins, F.M. Male-killing bacteria in insects: Mechanisms, incidence, and implications. Emerg. Infect. Dis. 2000, 6, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Darby, A.C.; Armstrong, S.D.; Bah, G.S.; Kaur, G.; Hughes, M.A.; Kay, S.M.; Koldkjær, P.; Rainbow, L.; Radford, A.D.; Blaxter, M.L.; et al. Analysis of gene expression from the Wolbachia genome of a filarial nematode supports both metabolic and defensive roles within the symbiosis. Genome Res. 2012, 22, 2467–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedeine, F.; Boulétreau, M.; Vavre, F. Wolbachia requirement for oogenesis: Occurrence within the genus Asobara (Hymenoptera, Braconidae) and evidence for intraspecific variation in A. tabida. Heredity 2005, 95, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Ryuichi, K.; Kikuchi, Y.; Meng, X.Y.; Fukatsu, T. Wolbachia as a bacteriocyte-associated nutritional mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriesner, P.; Hoffmann, A.A.; Lee, S.F.; Turelli, M.; Weeks, A.R. Rapid sequential spread of two Wolbachia variants in Drosophila simulans. PLoS Pathog. 2013, 9, e1003607. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.J.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, G.L.; Koga, R.; Xue, P.; Fukatsu, T.; Rasgon, J.L. Wolbachia infections are virulent and inhibit the human malaria parasite Plasmodium falciparum in Anopheles gambiae. PLoS Pathog. 2011, 7, e1002043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micieli, M.V.; Glaser, R.L. Somatic Wolbachia (Rickettsiales: Rickettsiaceae) levels in Culex quinquefasciatus and Culex pipiens (Diptera: Culicidae) and resistance to West Nile virus infection. J. Med. Entomol. 2014, 51, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Dutra, H.L.C.; Moreira, L.A. Exploiting intimate relationships: Controlling mosquito-transmitted disease with Wolbachia. Trends Parasitol. 2016, 32, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, H.A.; O’Neill, S.L. Controlling vector-borne diseases by releasing modified mosquitoes. Nat. Rev. Microbiol. 2018, 16, 508–518. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbe-Ormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–457. [Google Scholar] [CrossRef]
- Bian, G.; Joshi, D.; Dong, Y.; Lu, P.; Zhou, G.; Pan, X.; Xu, Y.; Dimopoulos, G.; Xi, Z. Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science 2013, 340, 748–751. [Google Scholar] [CrossRef]
- Ryan, P.A.; Turley, A.P.; Wilson, G.; Hurst, T.P.; Retzki, K.; Brown-Kenyon, J.; Hodgson, L.; Kenny, N.; Cook, H.; Montgomery, B.L. Establishment of wMel Wolbachia in Aedes aegypti mosquitoes and reduction of local dengue transmission in Cairns and surrounding locations in northern Queensland, Australia. Gates Open Res. 2019, 3, 1547. [Google Scholar] [CrossRef]
- Nazni, W.A.; Hoffmann, A.A.; Afizah, A.N.; Cheong, Y.L.; Mancini, M.V.; Golding, N.; Kamarul, G.M.R.; Arif, M.A.K.; Thohir, H.; Syamimi, H.N. Establishment of Wolbachia strain wAlbB in Malaysian populations of Aedes aegypti for dengue control. Curr. Biol. 2019, 29, 4241–4248. [Google Scholar] [CrossRef] [Green Version]
- Clark, T.L.; Meinke, L.J.; Skoda, S.R.; Foster, J.E. Occurrence of Wolbachia in selected diabroticite (Coleoptera: Chrysomelidae) beetles. Ann. Entomol. Soc. Am. 2001, 94, 877–885. [Google Scholar] [CrossRef] [Green Version]
- Jäckel, R.; Mora, D.; Dobler, S. Evidence for selective sweeps by Wolbachia infections: Phylogeny of Altica leaf beetles and their reproductive parasites. Mol. Ecol. 2013, 22, 4241–4255. [Google Scholar] [CrossRef] [PubMed]
- Kajtoch, Ł.; Kotásková, N. Current state of knowledge on Wolbachia infection among Coleoptera: A systematic review. PeerJ 2018, 6, e4471. [Google Scholar] [CrossRef] [Green Version]
- Lachowska, D.; Kajtoch, Ł.; Knutelski, S. Occurrence of Wolbachia in central European weevils: Correlations with host systematics, ecology and biology. Entomol. Exp. Appl. 2010, 135, 105–118. [Google Scholar] [CrossRef]
- Maleki-Ravasan, N.; Akhavan, N.; Raz, A.; Jafari, M.; Zakeri, S.; Djadid, N.D. Co-occurrence of pederin-producing and Wolbachia endobacteria in Paederus fuscipes Curtis, 1840 (Coleoptera: Staphilinidae) and its evolutionary consequences. MicrobiologyOpen 2018, 8, e777. [Google Scholar] [CrossRef] [Green Version]
- Sontowski, R.; Bernhard, D.; Bleidorn, C.; Schlegel, M.; Gerth, M. Wolbachia distribution in selected beetle taxa characterized by PCR screens and MLST data. Ecol. Evol. 2015, 5, 4345–4353. [Google Scholar] [CrossRef]
- Yun, Y.; Peng, Y.; Liu, F.X.; Lei, C. Wolbachia screening in spiders and assessment of horizontal transmission between predator and prey. Neotrop. Entomol. 2011, 40, 164–169. [Google Scholar]
- Zhang, Q.; Wu, X.; Liu, Z. Primary screening of plant essential oils as insecticides, fumigants, and repellents against the health pest Paederus fuscipes (Coleoptera: Staphylinidae). J. Econ. Entomol. 2016, 109, 2388–2396. [Google Scholar] [CrossRef]
- Frank, J.H.; Kanamitsu, K. Paederus, sensu lato (Coleoptera: Staphylinidae): Natural history and medical importance. J. Med. Entomol. 1987, 24, 155–191. [Google Scholar] [CrossRef]
- Veraldi, S.; Cuka, E.; Chiaratti, A.; Nazzaro, G.; Gianotti, R.; Süss, L. Paederus fuscipes dermatitis: A report of nine cases observed in Italy and review of the literature. Eur. J. Dermatol. 2013, 23, 387–391. [Google Scholar] [CrossRef]
- Schunkert, E.M.; Aschoff, N.S.; Grimmer, F.; Wiemann, C.; Zillikens, D. Paederus dermatitis—Touched by champion flies—Three clinical manifestations of pederin toxin-inflicted dermatitis. Int. J. Dermatol. 2018, 57, 989–991. [Google Scholar] [CrossRef]
- Ming, Q.L.; Shen, J.F.; Cheng, C.; Liu, C.M.; Feng, Z.J. Wolbachia infection dynamics in Tribolium confusum (Coleoptera: Tenebrionidae) and their effects on host mating behavior and reproduction. J. Econ. Entomol. 2015, 108, 1408–1415. [Google Scholar] [CrossRef]
- Dobson, S. Wolbachia infections are distributed throughout insect somatic and germ line tissues. Insect Biochem. Molec. 1999, 29, 153–160. [Google Scholar] [CrossRef]
- Binnington, K.C.; Hoffmann, A.A. Wolbachia-like organisms and cytoplasmic incompatibility in Drosophila simulans. J. Invertebr. Pathol. 1989, 54, 344–352. [Google Scholar] [CrossRef]
- Zouache, K.; Voronin, D.; Tran-Van, V. Persistent Wolbachia and cultivable bacteria infection in the reproductive and somatic tissues of the mosquito vector Aedes albopictus. PLoS ONE 2009, 4, e6388. [Google Scholar] [CrossRef] [Green Version]
- Frydman, H.M.; Li, J.M.; Robson, D.N.; Wieschaus, E. Somatic stem cell niche tropism in Wolbachia. Nature 2006, 441, 509–512. [Google Scholar] [CrossRef]
- Dittmer, J.; Beltran-bech, S.; Lesobre, J.; Raimond, M.; Johnson, M.; Bouchon, D. Host tissues as microhabitats for Wolbachia and quantitative insights into the bacterial community in terrestrial isopods. Mol. Ecol. 2014, 23, 2619–2635. [Google Scholar] [CrossRef]
- Espino, C.I.; Gomez, T.; Gonzalez, G. Detection of Wolbachia bacteria in multiple organs and feces of the triatomine insect Rhodnius pallescens (Hemiptera, Reduviidae). Appl. Environ. Microb. 2009, 75, 547–550. [Google Scholar] [CrossRef] [Green Version]
- Ijichi, N.; Kondo, N.; Matsumoto, R. Internal spatiotemporal population dynamics of infection with three Wolbachia strains in the adzuki bean beetle, Callosobruchus chinensis (Coleoptera: Bruchidae). Appl. Environ. Microb. 2002, 68, 4074–4080. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Martinez, J.; Rota-Stabelli, O.; Jiggins, F.M.; Miller, W.J. Age, tissue, genotype and virus infection regulate Wolbachia levels in Drosophila. Mol. Ecol. 2020, 29, 2063–2079. [Google Scholar] [CrossRef]
- Kellner, R.L.L.; Dettner, K. Allocation of pederin during lifetime of Paederus rove beetles (Coleoptera: Staphylinidae): Evidence for polymorphism of hemolymph toxin. J. Chem. Ecol. 1995, 21, 1719–1733. [Google Scholar] [CrossRef]
- Kador, M.; Horn, M.A.; Dettne, K. Novel oligonucleotide probes for in situ detection of pederin-producing endosymbionts of Paederus riparius rove beetles (Coleoptera: Staphylinidae). FEMS Microbiol. Lett. 2011, 319, 73–81. [Google Scholar] [CrossRef]
- Jiang, W.B.; Zhu, J.Q.; Chen, M.H.; Yang, Q.C.; Du, X.; Chen, S.Y.; Zhang, L.N.; Yu, Y.M.; Yu, W.D. Wolbachia infection status and genetic structure in natural populations of Polytremis nascens (Lepidoptera: Hesperiidae). Infect. Genet. Evol. 2014, 27, 202–211. [Google Scholar] [CrossRef]
- Werren, J.H.; Windsor, D.M. Wolbachia infection frequencies in insects: Evidence of a global equilibrium? Proc. Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.B.; Zhu, J.Q.; Wu, Y.J.; Li, L.Z.; Li, Y.Y.; Ge, C.; Wang, Y.; Endersby, N.M.; Hoffmann, A.A.; Yu, W.D. Influence of Wolbachia infection on mitochondrial DNA variation in the genus Polytremis (Lepidoptera: Hesperiidae). Mol. Phylogenet. Evol. 2018, 129, 158–170. [Google Scholar] [CrossRef]
- Baldo, L.; Lo, N.; Werren, J.H. Mosaic nature of Wolbachia surface protein. J. Bacteriol. 2005, 187, 5406–5418. [Google Scholar] [CrossRef] [Green Version]
- Baldo, L.; Dunning, H.J.C.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microb. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor an analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Lanfear, R.; Calcott, B.; Ho, S.Y.; Guindon, S. Partitionfinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.-A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Li, Y.Y.; Zhu, J.Q.; Ge, C.; Wang, Y.; Zhao, Z.M.; Ma, S.J.; Hoffmann, A.A.; Endersby, N.M.; Liu, Q.X.; Yu, W.D.; et al. Molecular Phylogeny and Historical Biogeography of the Butterfly Tribe Aeromachini Tutt (Lepidoptera: Hesperiidae) from China. Cells 2019, 8, 294. [Google Scholar] [CrossRef] [Green Version]
- Ali, H.; Muhammad, A.; Islam, S.U.; Islam, W.; Hou, Y.M. A novel bacterial symbiont association in the hispid beetle, Octodonta nipae (Coleoptera: Chrysomelidae), their dynamics and phylogeny. Microb. Pathog. 2018, 118, 378–386. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, B.; Lin, Y.; Chen, Z.; Zhang, X.; Ji, T.; Zhang, X.; Hou, Y. Identification of three prophenoloxidase-activating factors (PPAFs) from an invasive beetle Octodonta nipae Maulik (Coleoptera: Chrysomelidae) and their roles in the prophenoloxidase activation. Arch. Insect Biochem. Physiol. 2017, 96, e21425. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jiggins, F.M. Male-killing Wolbachia and mitochondrial DNA: Selective sweeps, hybrid introgression and parasite population dynamics. Genetics 2003, 164, 5–12. [Google Scholar]
- Augustinos, A.A.; Santos-Garcia, D.; Dionyssopoulou, E.; Moreira, M.; Papapanagiotou, A.; Scarvelakis, M.; Doudoumis, V.; Ramos, S.; Aguiar, A.F. Detection and characterization of Wolbachia infections in natural populations of Aphids: Is the hidden diversity fully unraveled? PLoS ONE 2011, 6, e28695. [Google Scholar] [CrossRef] [Green Version]
- Baldo, L.; Werren, J.H. Revisiting Wolbachia supergroup typing based on WSP: Spurious lineages and discordance with MLST. Curr. Microbiol. 2007, 55, 81–87. [Google Scholar] [CrossRef]
- Wang, G.H.; Jia, L.Y.; Xiao, J.H.; Huang, D.W. Discovery of a new Wolbachia supergroup in cave spider species and the lateral transfer of phage WO among distant hosts. Infect. Genet. Evol. 2016, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kondo, N.; Ijichi, N.; Shimada, M.; Fukatsu, T. Prevailing triple infection with Wolbachia in Callosobruchus chinensis (Coleoptera: Bruchidae). Mol. Ecol. 2002, 11, 167–180. [Google Scholar] [CrossRef]
- Zhang, X.; Luckhart, S.; Tu, Z.; Pfeiffer, D.G. Analysis of Wolbachia strains associated with Conotrachelus nenuphar (Coleoptera: Curculionidae) in the eastern United States. Environ. Entomol. 2010, 39, 396–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguero, M.S.; Confalonieri, V.A.; Guedes, J.V.; Lanteri, A.A. Wolbachia infection in the tribe Naupactini (Coleoptera, Curculionidae): Association between thelytokous parthenogenesis and infection status. Insect Mol. Biol. 2010, 19, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Majerus, M.E.N.; Hinrich, J.; Schulenburg, G.V.D.; Zakharov, I.A. Multiple causes of male-killing in a single sample of the two-spot ladybird, Adalia bipunctata (Coleoptera: Coccinellidae) from Moscow. Heredity 2010, 84, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Abrar, M.; Hou, Y. Infection density dynamics and phylogeny of Wolbachia associated with coconut hispine beetle, Brontispa longissima (Gestro) (Coleoptera: Chrysomelidae) by multilocus sequence type (MLST) genotyping. J. Microbiol. Biotechnol. 2018, 28, 796–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadfield, S.J.; Axton, J.M. Germ cells colonized by endosymbiotic bacteria. Nature 1999, 402, 482. [Google Scholar] [CrossRef] [PubMed]
- Serbus, L.R.; Sullivan, W. A cellular basis for Wolbachia recruitment to the host germline. PLoS Pathog. 2007, 3, e190. [Google Scholar] [CrossRef]
- Pavan, M.; Bo, G. Pederin, toxic principle obtained in the crystalline state from the beetle Paederus fuscipes Curt. Physiol. Comp. Oecol. 1953, 3, 307–312. [Google Scholar]
- Kellner, R.L.L. Suppression of pederin biosynthesis through antibiotic elimination of endosymbionts in Paederus sabaeus. J. Insect Physiol. 2001, 47, 475–483. [Google Scholar] [CrossRef]
- Piel, J. A polyketide synthase-peptide synthetase gene cluster from an uncultured bacterial symbiont of Paederus beetles. Proc. Natl. Acad. Sci. USA 2002, 99, 14002–14007. [Google Scholar] [CrossRef] [Green Version]
- Kellner, R.L.L.; Dettner, K. Differential efficacy of toxic pederin in deterring potential arthropod predators of Paederus (Coleoptera: Staphylinidae) offspring. Oecologia 1996, 107, 293–300. [Google Scholar] [CrossRef]
- Pankewitz, F.; Zollmer, A.; Hilker, M.; Graser, Y. Presence of Wolbachia in insect eggs containing antimicrobially active anthraquinones. Microb. Ecol. 2007, 54, 713–721. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene/Region | Primers | Sequence (5′–3′) | Amplicon Length | Annealing Temperature | Reference |
|---|---|---|---|---|---|
| 16S rRNA | 16S_F 16S_R | TTGTAGCCTGCTATGGTATAACT GAATAGGTATGATTTTCATGT | 870 bp | 55 °C | [43] |
| gatB | gatB_F1 gatB_R1 | GAKTTAAAYCGYGCAGGBGTT TGGYAAYTCRGGYAAAGATGA | 471 bp | 54 °C | [46] |
| coxA | coxA_F1 coxA_R1 | TTGGRGCRATYAACTTTATAG TCTAAAGACTTTKACRCCAGT | 487 bp | 54 °C | [46] |
| hcpA | coxA_F1 coxA_R1 | GAAATARCAGTTGCTGCAAA GAAAGTYRAGCAAGYTCTG | 515 bp | 54 °C | [46] |
| ftsZ | ftsZ_F1 ftsZ_R1 | ATYATGGARCATATAAARGATAGTCR AGYAATGGATTRGATAT | 524 bp | 54 °C | [46] |
| fbpA | fbpA_F1 fbpA_R1 | GCTGCTCCRCTTGGYWTGAT CCRCCAGARAAAAYYACTATTC | 509 bp | 59 °C | [46] |
| wsp | wsp1_F1 wsp1_R1 | TGGTATTGGTGTTGGTGCAG AACCGAAATAACGAGCTCCA | 158 bp | 50 °C | [53] |
| RPS3 | RPS3_F RPS3_R | CCCAGATAATCATTATCG CAGATTGAATGTGTGACAC | 191bp | 50 °C | [54] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, C.; Hu, J.; Zhao, Z.; Hoffmann, A.A.; Ma, S.; Shen, L.; Fang, J.; Zhu, J.; Yu, W.; Jiang, W. Phylogeny and Density Dynamics of Wolbachia Infection of the Health Pest Paederus fuscipes Curtis (Coleoptera: Staphylinidae). Insects 2020, 11, 625. https://doi.org/10.3390/insects11090625
Ge C, Hu J, Zhao Z, Hoffmann AA, Ma S, Shen L, Fang J, Zhu J, Yu W, Jiang W. Phylogeny and Density Dynamics of Wolbachia Infection of the Health Pest Paederus fuscipes Curtis (Coleoptera: Staphylinidae). Insects. 2020; 11(9):625. https://doi.org/10.3390/insects11090625
Chicago/Turabian StyleGe, Chen, Jiayao Hu, Zimiao Zhao, Ary A. Hoffmann, Shuojia Ma, Li Shen, Jie Fang, Jianqing Zhu, Weidong Yu, and Weibin Jiang. 2020. "Phylogeny and Density Dynamics of Wolbachia Infection of the Health Pest Paederus fuscipes Curtis (Coleoptera: Staphylinidae)" Insects 11, no. 9: 625. https://doi.org/10.3390/insects11090625