Impact of Sacbrood Virus on Larval Microbiome of Apis mellifera and Apis cerana


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Inoculum
2.2. SBV Infection in Honeybee Larvae
2.3. Total RNA and DNA Extraction
2.4. SBV Expression in Honeybee Larvae
2.5. Amplicon Sequencing Analysis
2.6. Sequence Analysis
2.6.1. Sequences Processing
2.6.2. Clustering Sequences into Operational Taxonomic Units (OTUs)
2.7. Alpha Diversity Analysis
2.8. Beta Diversity Analysis
2.9. Statistical Analysis
3. Results
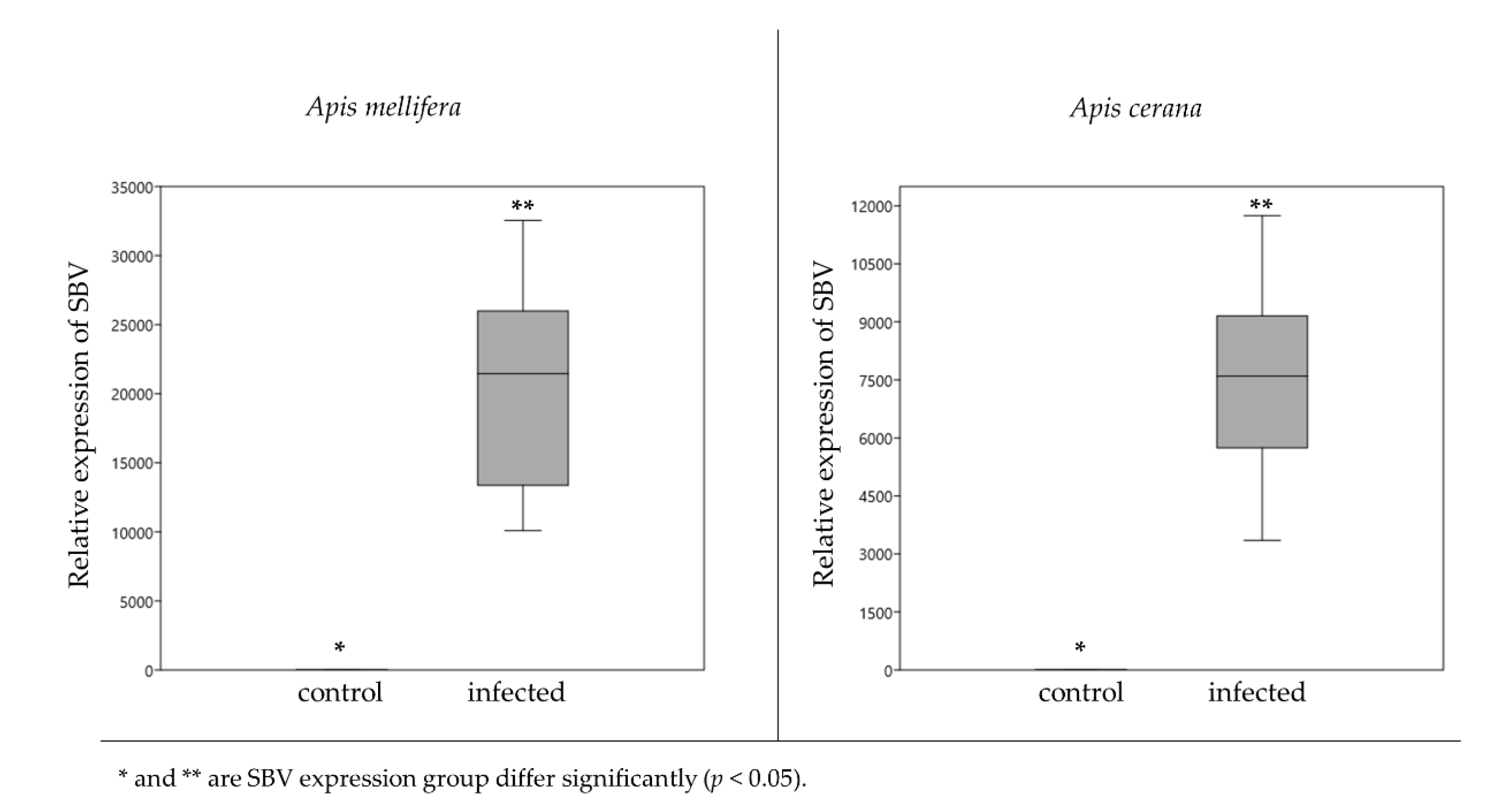
3.1. Sacbrood Virus Infection
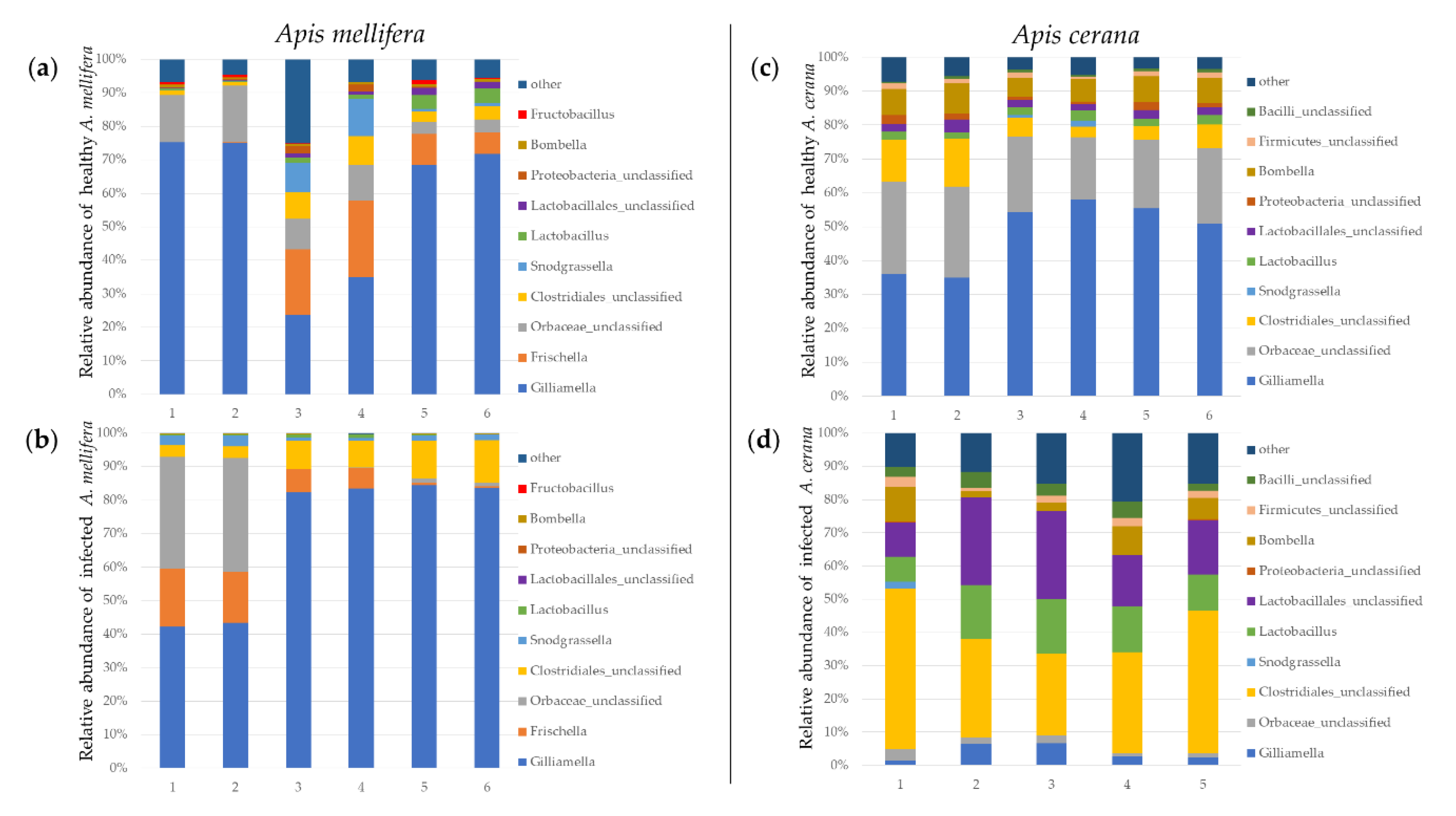
3.2. Larval Microbiome
3.2.1. The Gut Bacteria of A. mellifera in the Control and Infected Larvae
3.2.2. The Larval Microbiome of A. cerana in the Control and Infected Larvae
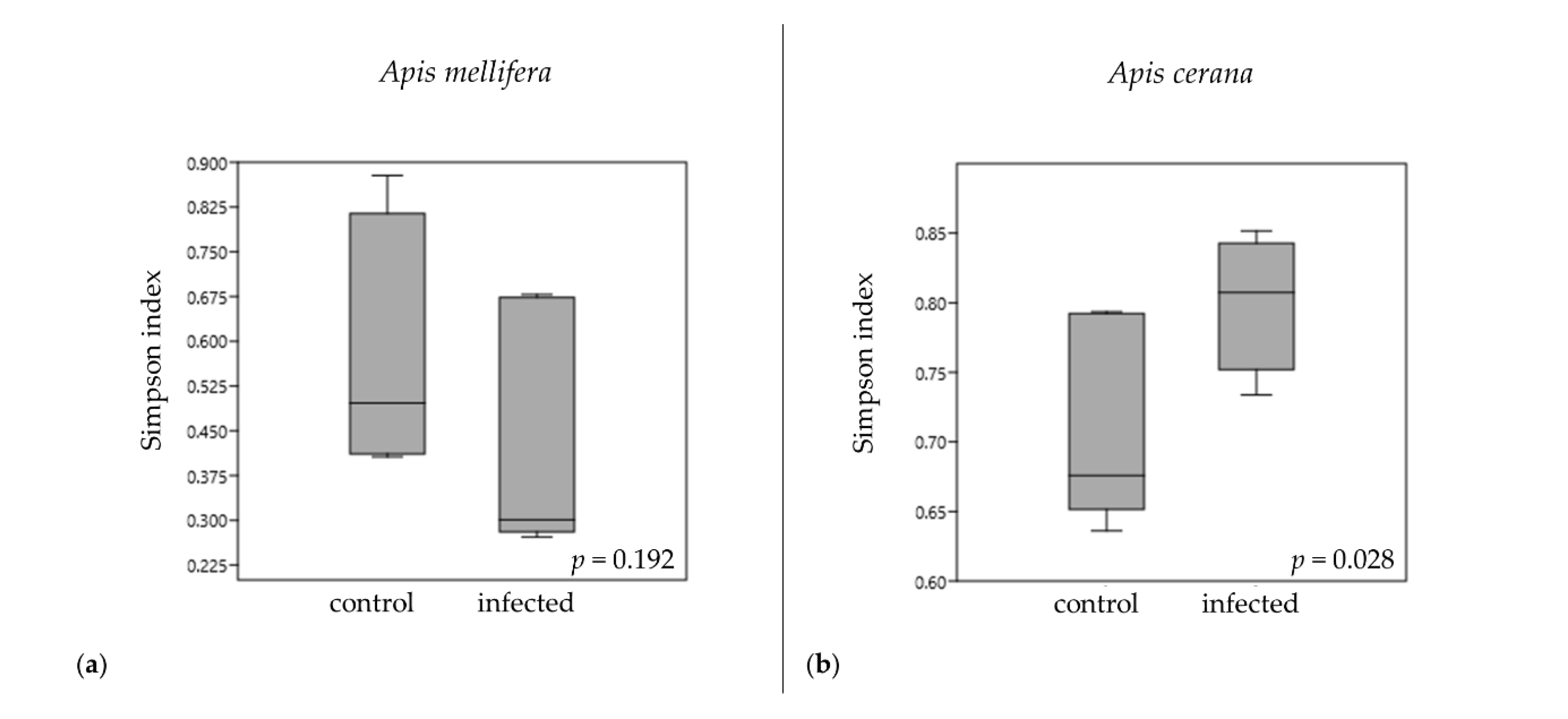
3.3. Alpha Diversity Indices of Gut Bacterial Communities
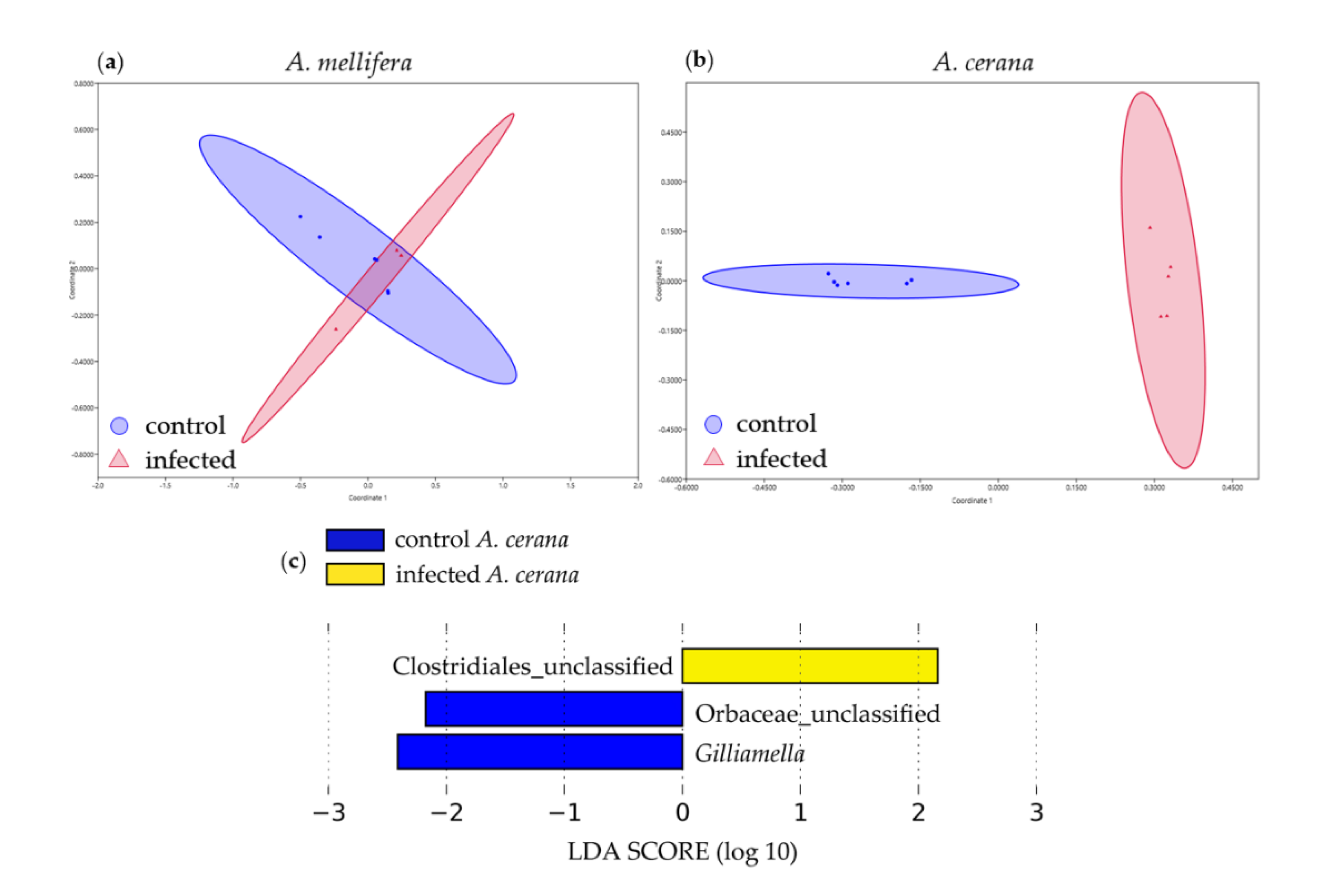
3.4. Beta Diversity Analysis of Larval Bacterial Communities
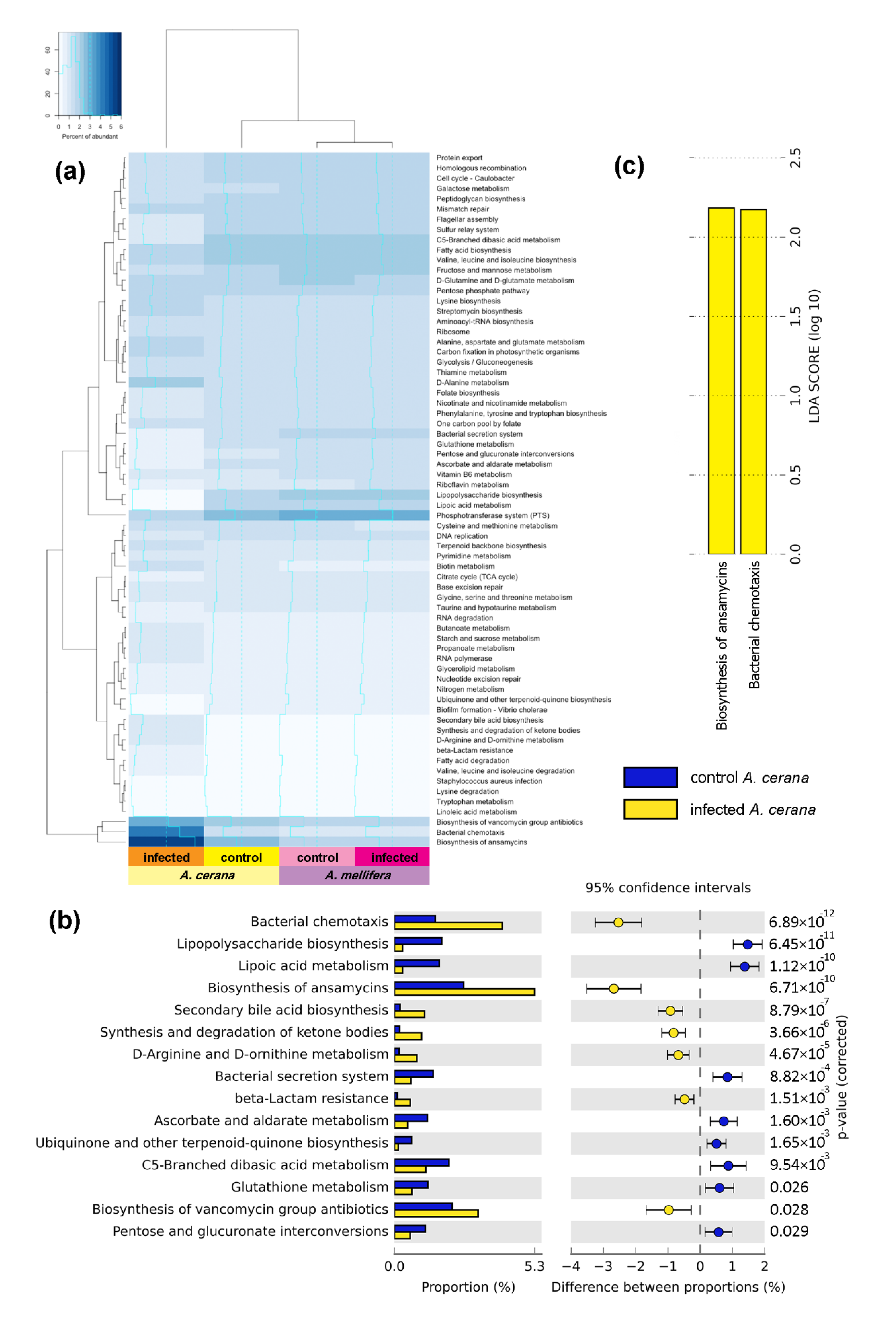
3.5. Functional Gene Prediction from 16S rRNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Kremen, C.; Morales, J.M.; Bommarco, R.; Cunningham, S.; Carvalheiro, L.G.; Chacoff, N.P.; Dudenhöffer, J.-H.; Greenleaf, S.S.; et al. Stability of pollination services decreases with isolation from natural areas despite honey bee visits. Ecol. Lett. 2011, 14, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.Q.; Adams, M.J.; Lefkowitz, E.J.; Carstens, E.B. (Eds.) Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Philadelphia, PA, USA, 2012. [Google Scholar]
- White, G.F. Sacbrood; US Department of Agriculture: Washington, DC, USA, 1917; Volume 431, pp. 1–55. [Google Scholar] [CrossRef]
- Nguyen, N.T.B.; Le, T.H. Complete Genome Sequence of Sacbrood Virus Strain SBM2, Isolated from the Honeybee Apis cerana in Vietnam. Genome Announc. 2013, 1, e00076-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, L. Recent research on honey bee viruses. Bee World 1975, 56, 55–64. [Google Scholar] [CrossRef]
- Anderson, D.L.; Gibbs, A.J. Transpuparial transmission of Kashmir bee virus and sacbrood virus in the honeybee (Apis mellifera). Ann. Appl. Biol. 1989, 114, 1–7. [Google Scholar] [CrossRef]
- Wang, D.-I.; Mofller, F.E. The division of labor and Queen attendance behavior of nosema-infected worker honey bees. J. Econ. Entomol. 1970, 63, 1539–1541. [Google Scholar] [CrossRef]
- Bailey, L.; Fernando, E.F.W. Effects of sacbrood virus on adult honey-bees. Ann. Appl. Boil. 1972, 72, 27–35. [Google Scholar] [CrossRef]
- Ellis, J.D.; A Munn, P. The worldwide health status of honey bees. Bee World 2005, 86, 88–101. [Google Scholar] [CrossRef]
- Choe, S.E.; Nguyen, L.T.; Noh, J.H.; Kweon, C.H.; Reddy, K.E.; Koh, H.B.; Chang, K.Y.; Kang, S.W. Analysis of the complete genome sequence of two Korean sacbrood viruses in the Honey bee, Apis mellifera. Virology 2012, 432, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Bailey, L.; Gibbs, A.; Woods, R. Sacbrood virus of the larval honey bee (Apis mellifera Linnaeus). Virology 1964, 23, 425–429. [Google Scholar] [CrossRef]
- Allen, M.; Ball, B. The incidence and world distribution of honeybee viruses. Bee world 1996, 77, 141–162. [Google Scholar] [CrossRef]
- Anderson, D.L. Viruses of Apis cerana and Apis mellifera; Enviroquest Ltd.: Cambridge, ON, Canada, 1995; pp. 161–170. [Google Scholar]
- Liu, X.; Zhang, Y.; Yan, X.; Han, R. Prevention of Chinese sacbrood virus infection in Apis cerana using RNA Interference. Curr. Microbiol. 2010, 61, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Rana, B.; Garg, I.; Khurana, S.P.; Verma, L.; Agrawal, H. Thai sacbrood virus of honeybees (Apis cerana indica F) in north-west Himalayas. Indian J. Virol. 1986, 2, 127–131. [Google Scholar]
- Bailey, L.; Ball, B.V.; Perry, J.N. Association of viruses with two protozoal pathogens of the honey bee. Ann. Appl. Boil. 1983, 103, 13–20. [Google Scholar] [CrossRef]
- Choe, S.-E.; Nguyen, T.T.-D.; Hyun, B.-H.; Noh, J.-H.; Lee, H.-S.; Lee, C.-H.; Kang, S.-W. Genetic and phylogenetic analysis of South Korean sacbrood virus isolates from infected honey bees (Apis cerana). Veter. Microbiol. 2012, 157, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Mehmood, S.; Zhang, G.; Song, Y.; Su, S.; Huang, W.F.; Huang, H.; Zhang, Y.; Geng, H.; Huang, W.-F. Visualizing sacbrood virus of honey bees via transformation and coupling with enhanced green fluorescent protein. Viruses 2020, 12, 224. [Google Scholar] [CrossRef] [Green Version]
- Lee, F.J.; Rusch, D.B.; Stewart, F.J.; Mattila, H.R. Newton, I.L. Saccharide breakdown and fermentation by the honeybee gut microbiome. Environ. Microbiol. 2014, 17, 796–815. [Google Scholar] [CrossRef]
- Wu, M.; Sugimura, Y.; Takaya, N.; Takamatsu, D.; Kobayashi, M.; Taylor, D.; Yoshiyama, M. Characterization of bifidobacteria in the digestive tract of the Japanese honeybee Apis cerana japonica. J. Invertebr. Pathol. 2013, 112, 88–93. [Google Scholar] [CrossRef]
- Dale, C.; Moran, N.A. Molecular interactions between bacterial symbionts and their hosts. Cell 2006, 126, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Martinson, V.G.; Moy, J.; Moran, N.A. Establishment of characteristic gut bacteria during development of the honeybee worker. Appl. Environ. Microbiol. 2012, 78, 2830–2840. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.H.; Hong, I.P.; Bok, J.I.; Kim, B.Y.; Song, J.; Weon, H.Y. Pyrosequencing analysis of the bacterial communities in the guts of honeybees Apis cerana and Apis mellifera in Korea. J. Microbiol. 2012, 50, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Vojvodic, S.; Rehan, S.M.; Anderson, K.E. Microbial gut diversity of Africanized and European honeybee larval instars. PLoS ONE 2013, 8, e72106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazzi, F.; Brown, S.P.; Annoscia, D.; Del Piccolo, F.; Di Prisco, G.; Varricchio, P.; Della Vedova, G.; Cattonaro, F.; Caprio, E.; Pennacchio, F. Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLOS Pathog. 2012, 8, e1002735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boncristiani, H.F.; Evans, J.D.; Chen, Y.; Pettis, J.; Murphy, C.; Lopez, D.L.; Finstrom, M.S.; Strand, M.; Tarpy, D.R.; Rueppell, O. In vitro infection of pupae with israeli acute paralysis virus suggests disturbance of transcriptional homeostasis in honey bees (Apis mellifera). PLoS ONE 2013, 8, e73429. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D. Beepath: An ordered quantitative-PCR array for exploring honey bee immunity and disease. J. Invertebr. Pathol. 2006, 93, 135–139. [Google Scholar] [CrossRef]
- Kaftanoglu, O.; Linksvayer, T.A.; Page, R.E., Jr. Rearing honey bees, Apis mellifera, in vitro I: Effects of sugar concentrations on survival and development. J. Insect Sci. 2011, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2018, 47, D590–D595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Cultivation and characterization of the gut symbionts of honey bees and bumble bees: Description of Snodgrassella alvi gen. nov., sp. nov., a member of the family Neisseriaceae of the Betaproteobacteria, and Gilliamella apicola gen. nov., sp. nov., a member of Orbaceae fam. nov., Orbales ord. nov., a sister taxon to the order‘ Enterobacteriales’ of the Gammaproteobacteria. Int. J. Syst. Evol. Microbiol. 2013, 63, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Babendreier, D.; Joller, D.; Romeis, J.; Bigler, F.; Widmer, F.; Romeis, J. Bacterial community structures in honeybee intestines and their response to two insecticidal proteins. FEMS Microbiol. Ecol. 2007, 59, 600–610. [Google Scholar] [CrossRef]
- Martinson, V.G.; Danforth, B.N.; Minckley, R.L.; Rueppell, O.; Tingek, S.; Moran, N.A. A simple and distinctive microbiota associated with honey bees and bumble bees. Mol. Ecol. 2010, 20, 619–628. [Google Scholar] [CrossRef]
- Scardovi, V.; Trovatelli, L.D. New species of bifid bacteria from Apis mellifica L. and Apis indica F. A contribution to the taxonomy and biochemistry of the genus Bifidobacterium. Zent. Bakteriol. Parasitenkd. Infekt. Hyg. 1969, 123, 64–88. [Google Scholar]
- Bottacini, F.; Milani, C.; Turroni, F.; Sanchez, B.; Foroni, E.; Duranti, S.; Serafini, F.; Viappiani, A.; Strati, F.; Ferrarini, A.; et al. Bifidobacterium asteroides PRL2011 Genome Analysis Reveals Clues for Colonization of the Insect Gut. PLoS ONE 2012, 7, e44229. [Google Scholar] [CrossRef] [Green Version]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.; Palacios, G.F.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.E.; Yoo, M.S.; Kim, Y.-H.; Kim, N.-H.; Ramya, M.; Jung, H.-N.; Thao, L.T.B.; Lee, H.-S.; Kang, S.-W. Homology differences between complete Sacbrood virus genomes from infected Apis mellifera and Apis cerana honeybees in Korea. Virus Genes 2016, 52, 281–289. [Google Scholar] [CrossRef]
- Engel, P.; Martinson, V.G.; Moran, N.A. Functional diversity within the simple gut microbiota of the honey bee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancini, G.; Grandi, M. Biosynthesis of Ansamycins. In Biosynthesis; Springer: Berlin/Heidelberg, Germany, 1981; pp. 12–40. [Google Scholar]
- Wehrli, W. Ansamycins. Chemistry, biosynthesis and biological activity. In Medicinal Chemistry; Springer: Berlin/Heidelberg, Germany, 1977; pp. 21–49. [Google Scholar]
- Lu, C.; Shen, Y. A Novel Ansamycin, Naphthomycin K from Streptomyces sp. J. Antibiot. 2007, 60, 649–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, J. Chemotaxis in Bacteria. Annu. Rev. Biochem. 1975, 44, 341–356. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yongsawas, R.; Chaimanee, V.; Pettis, J.S.; Boncristiani Junior, H.F.; Lopez, D.; In-on, A.; Chantawannakul, P.; Disayathanoowat, T. Impact of Sacbrood Virus on Larval Microbiome of Apis mellifera and Apis cerana. Insects 2020, 11, 439. https://doi.org/10.3390/insects11070439
Yongsawas R, Chaimanee V, Pettis JS, Boncristiani Junior HF, Lopez D, In-on A, Chantawannakul P, Disayathanoowat T. Impact of Sacbrood Virus on Larval Microbiome of Apis mellifera and Apis cerana. Insects. 2020; 11(7):439. https://doi.org/10.3390/insects11070439
Chicago/Turabian StyleYongsawas, Rujipas, Veeranan Chaimanee, Jeffery S. Pettis, Humberto Freire Boncristiani Junior, Dawn Lopez, Ammarin In-on, Panuwan Chantawannakul, and Terd Disayathanoowat. 2020. "Impact of Sacbrood Virus on Larval Microbiome of Apis mellifera and Apis cerana" Insects 11, no. 7: 439. https://doi.org/10.3390/insects11070439