Relish as a Candidate Marker for Transgenerational Immune Priming in a Dampwood Termite (Blattodae: Archeotermopsidae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
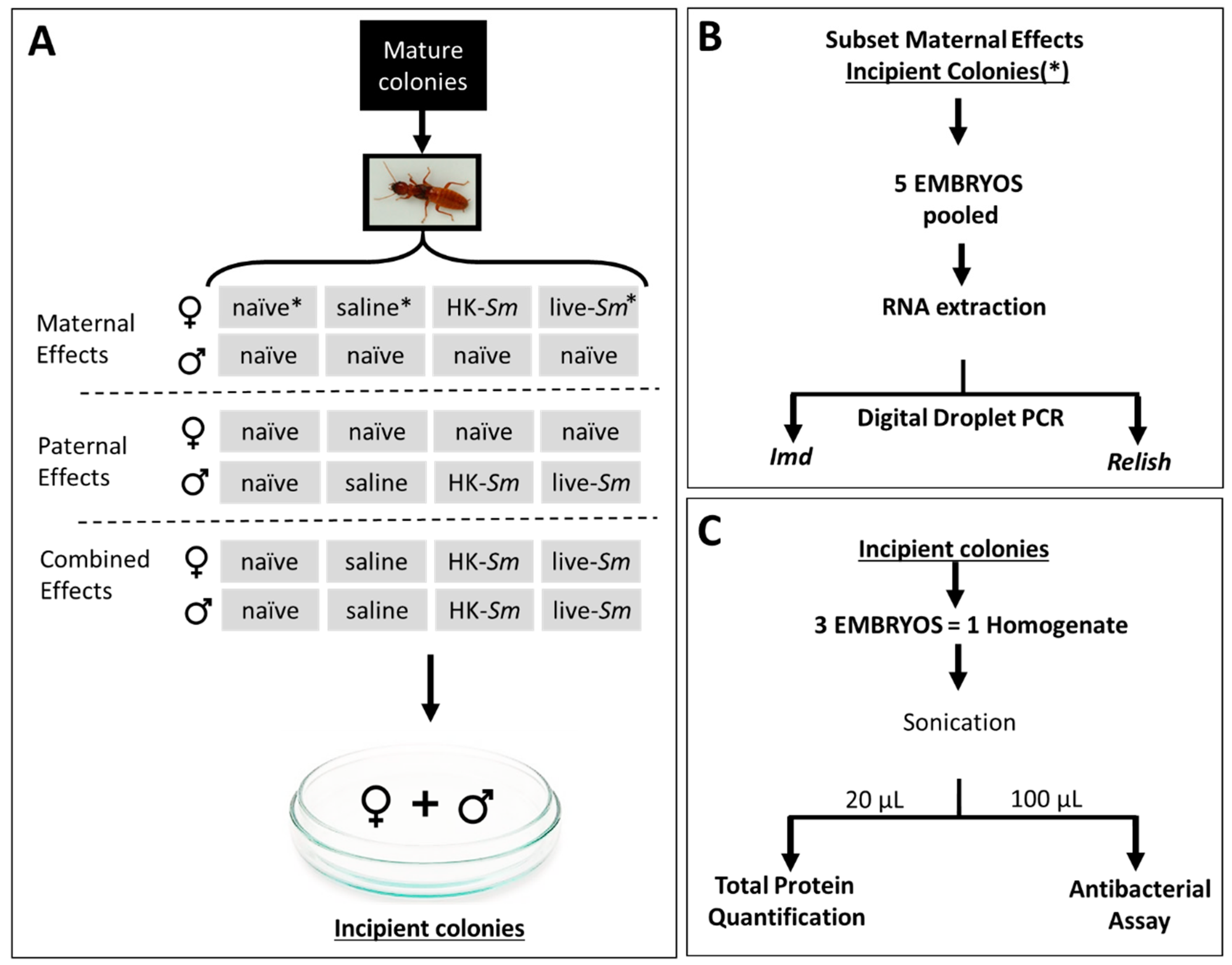
2.1. Colony Collection and Establishment of Incipient Termite Colonies
2.2. Parental Treatments, Microinjections, and Embryo Collection
2.3. Transcript Analyses
2.4. Quantification of Total Embryonic Protein
2.5. Antibacterial Assay
2.6. Statistical Analysis
3. Results
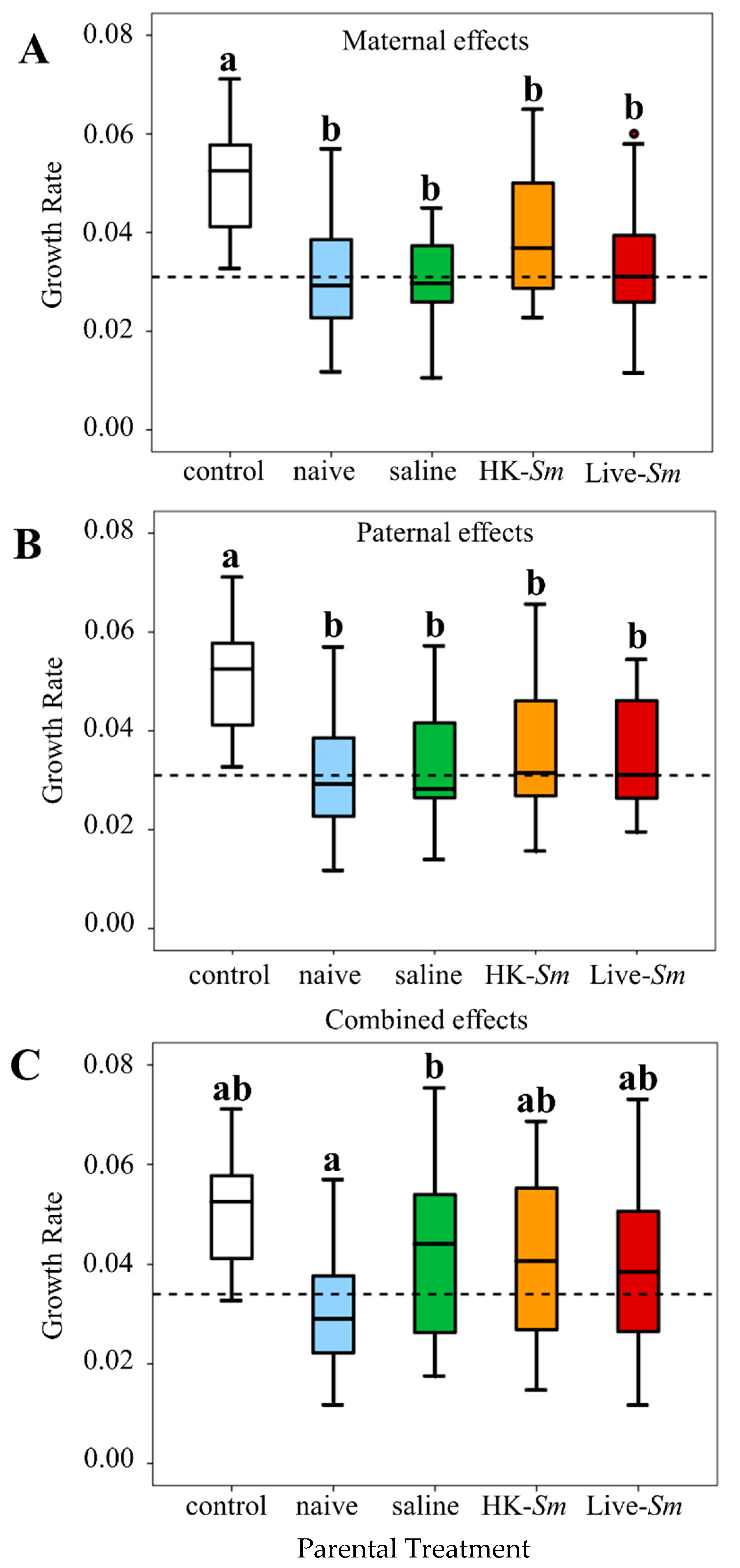
3.1. mRNA Abundance of IMD and Relish
3.2. Total Protein
3.3. Embryonic Antibacterial Properties
4. Discussion
4.1. mRNA Abundance of Relish
4.2. Incongruity between Transcriptional and Antibacterial Data
4.3. Termite Embryos Have Antimicrobial Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jablonka, E.; Raz, G. Transgenerational epigenetic inheritance: Prevalence, mechanisms, and implications for the study of heredity and evolution. Q. Rev. Biol. 2009, 2, 131–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondurianski, R.; Crean, A.J.; Day, T. The implications of nongenetic inheritance for evolution in changing environments. Evol. Appl. 2012, 5, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Klosin, A.; Casas, E.; Hildago-Carcedo, C.; Vavouri, T.; Lehner, B. Transgenerational transmission of environmental information in C. elegans. Science 2017, 356, 320–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, O.; Beemelmanns, A.; Barribeau, S.M.; Sadd, B.M. Recent advances in vertebrate and invertebrate transgenerational immunity in the light of ecology and evolution. Heredity 2018, 121, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tetreau, G.; Dhinaut, B.; Gourbal, B.; Moret, Y. Trans-generational immune priming in invertebrates: Current knowledge and future prospects. Front. Immunol. 2019, 10, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glastad, K.M.; Hunt, B.G.; Goodisman, M.A.D. Epigenetics in insects: Genome regulation and the generation of phenotypic diversity. Annu. Rev. Entomol. 2019, 64, 185–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacour, G.; Vernichon, F.; Cadilhac, N.; Boyer, S.; Lagneau, C.; Hance, T. When mothers anticipate: Effects of the prediapause stage on embryo development time and of maternal photoperiod on eggs of a temperate and a tropical strains of Aedes albopictus (Diptera: Culicidae). J. Insect Physiol. 2014, 71, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Zizzari, Z.V.; Van Straalen, N.M.; Ellers, J. Transgenerational effects of nutrition are different for sons and daughters. J. Evol. Biol. 2016, 29, 1317–1327. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, S.A.; Saul, M.C.; Seward, C.H.; Zhang, H.; Bensky, M.; James, N.; Zhao, S.D.; Chandrasekaran, S.; Stubbs, L.; Bell, A.M. Temporal dynamics of neurogenomic plasticity in response to social interactions in male threespined sticklebacks. PLoS Genet. 2017, 13, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.R.; Castoe, T.; Holmes, J.; Packer, M.; Biles, K.; Walsh, M.; Munch, S.B.; Post, D.M. Local adaptation in transgenerational responses to predators. Proc. R. Soc. 2016, 283, 1–9. [Google Scholar] [CrossRef]
- Mousseu, T.A.; Fox, C.W. The adaptive significance of maternal effects. Trends Ecol. Evol. 1998, 13, 403–407. [Google Scholar] [CrossRef]
- Bonduriansky, R.; Day, T. Nongenetic inheritance and its evolutionary implications. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 103–125. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A. Transgenerational epigenetic inheritance: Focus on soma to germline information transfer. Prog. Biophys. Mol. Biol. 2013, 113, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Sadd, B.; Kleinlogel, Y.; Schmid-Hempel, R.; Schmid-Hempel, P. Trans-generational immune priming in a social insect. Biol. Lett. 2005, 1, 386–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigeault, R.; Garnier, R.; Rivero, A.; Gandon, S. Evolution of transgenerational immunity in invertebrates. Proc. R. Soc. 2016, 283, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fraune, S.; Augustin, R.; Anton-Erxleben, F.; Wittlieb, J.; Gelhaus, C.; Klimovich, V.B.; Samoilovich, M.P.; Bosch, T.C.G. In an early branching metazoan, bacterial colonization of the embryo is controlled by maternal antimicrobial peptides. Proc. Natl. Acad. Sci. USA 2014, 107, 18067–18072. [Google Scholar] [CrossRef] [Green Version]
- Seppola, M.; Johnsen, H.; Mennen, S.; Myrnes, B.; Tveiten, H. Maternal transfer and transcriptional onset of immune genes during ontogenesis in Atlantic cod. Dev. Comp. Immunol. 2009, 33, 1205–1211. [Google Scholar] [CrossRef]
- Freitak, D.; Schmidtberg, H.; Dickel, F.; Lochnit, G.; Vogel, H.; Vilcinskas, A. The maternal transfer of bacteria can mediate trans-generational immune priming in insects. Virulence 2014, 5, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Knorr, E.; Schmidtberg, H.; Arslan, D.; Bingsohn, L.; Vilcinskas, A. Translocation of bacteria from the gut to the egg triggers maternal transgenerational immune priming in Tribolium casteneum. Biol. Lett. 2015, 11, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Trauer-kizilelma, U.; Hilker, M. Insect parents improve the anti-parasitic and anti-bacterial defence of their offspring by priming the expression of immune-relevant genes. Insect Biochem. Mol. 2015, 64, 91–99. [Google Scholar] [CrossRef]
- Gegner, J.; Baudach, A.; Mukherjee, K.; Halitschke, R.; Vogel, H.; Vilcinskas, A. Epigenetic mechanisms are involved in sex-specific trans-generational immune priming in the Lepidopteran model host Manduca sexta. Front. Physiol. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trauer, U.; Hilker, M. Parental legacy in insects: Variation of transgenerational immune priming during offspring development. PLoS ONE 2013, 8, e63392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosengaus, R.B.; Hays, N.; Biro, C.; Kemos, J.; Zaman, M.; Murray, J.; Gezahegan, B.; Smith, W. Pathogen-induced maternal effects result in enhanced immune responsiveness across generations. Ecol. Evol. 2017, 7, 2925–2935. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, C.G.C.; van der Zee, M. Immune competence in insect eggs depends on the extraembryonic serosa. Dev. Comp. Immunol. 2013, 41, 263–269. [Google Scholar] [CrossRef]
- Eggert, H.; Kurtz, J.; de Burh, M.F.D. Different effects of paternal trans-generational immune priming on survival and immunity in step and genetic offspring. Proc. R. Soc. 2014, 281, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.J.; Hajek, A.E. Maternal exposure of a beetle to pathogens protects offspring against fungal disease. PLoS ONE 2015, 10, e0125197. [Google Scholar] [CrossRef]
- Linder, J.E.; Promislow, D.E.L. Cross-generational fitness effects of infection in Drosophila melanogaster. Fly 2009, 3, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Moret, Y. ‘Trans-generational immune priming’: Specific enhancement of the antimicrobial immune response in the mealworm beetle, Tenebrio Molit. Proc. R. Soc. 2006, 273, 1399–1405. [Google Scholar] [CrossRef] [Green Version]
- Rubenstein, D.R.; Abbot, P. Comparative Social Evolution; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Gao, Q.; Thompson, G.J. Social context affects immune gene expression in a subterranean termite. Insect Soc. 2015, 62, 167–170. [Google Scholar] [CrossRef]
- Smith, M.J.; Telfer, S.; Kallio, E.R.; Burthe, S.; Cook, A.R.; Lambin, X.; Begon, M. Host-pathogen time series data in wildlife support a transmission function between density and frequency dependence. Proc. Natl. Acad. Sci. USA 2009, 106, 7905–7909. [Google Scholar] [CrossRef] [Green Version]
- Barribeau, S.M.; Schmid-Hempel, P.; Sadd, B.M. Royal decree: Gene expression in trans-generationally immune primed bumblebee workers mimics a primary immune response. PLoS ONE 2016, 11, e0159635. [Google Scholar] [CrossRef] [PubMed]
- Sadd, B.M.; Schmid-Hempel, P. Facultative but persistent transgenerational immunity via the mother’s eggs in bumblebees. Curr. Biol. 2007, 17, R1046–R1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, J.H.; Schuehly, W.; Crailsheim, K. Trans-generational immune priming in honeybees. Proc. R. Soc. 2014, 281, 1–8. [Google Scholar] [CrossRef]
- Salmela, H.; Amdam, G.V.; Freitak, D. Transfer of immunity from mother to offspring in mediated via egg-yolk protein viellogenin. PLoS Pathog. 2015, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bordoni, A.; Dapporto, L.; Tatini, I.; Celli, M.; Bercigli, M.; Barrufet, S.R.; Perito, B.; Turillazzi, S. Trans-generational immunization in the acrobat ant Cremtogaster scutellaris. Biol. Lett. 2018, 14, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Fuch, S.-A.; Sundström, L.; Bos, N.; Stucki, D.; Freitak, D. Induced immune responses in Formica fusca (Hymenoptera: Formicidae). Myrmecol. News 2018, 28, 53–66. [Google Scholar] [CrossRef]
- Rosengaus, R.B.; Traniello, J.F.A.; Bulmer, M. Ecology, behavior, and evolution of disease resistance in termites. In Biology of Termites: A Modern Synthesis; Bignell, D., Yves, R., Lo, N., Eds.; Springer: New York, NY, USA, 2011; pp. 165–191. [Google Scholar]
- Rosengaus, R.B.; Traniello, J.F.A. Disease risk as a cost of outbreeding in the termite Zootermopsis angusticollis. Proc. Nat. Acad. Sci. USA 1993, 90, 6641–6645. [Google Scholar] [CrossRef] [Green Version]
- Calleri II, D.V.; Rosengaus, R.B.; Traniello, J.F.A. Disease and colony foundation in the dampwood termite Zootermopsis angusticollis: The survival advantage of nestmate pairs. Naturwissenschaften 2005, 92, 300–304. [Google Scholar] [CrossRef]
- Hartke, T.R.; Rosengaus, R.B. Costs of pleometrosis in a polygamous termite. Proc. R. Soc. 2013, 280, 20122563. [Google Scholar] [CrossRef] [Green Version]
- Cole, E.L.; Ilieş, I.; Rosengaus, R.B. Competing physiological demands during incipient colony foundation in a social insects: Consequences of pathogenic stress. Front. Ecol. Evol. 2018, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cole, E.L.; Bayne, H.; McDavid, J.J.; Rosengaus, R.B. Young but not Defenseless: Antifungal Activity during Embryonic Development of a Social Insect; Dryad, 2020; Available online: https://datadryad.org/stash/dataset/doi:10.5061/dryad.r9f25fj (accessed on 18 February 2020).
- Rosengaus, R.B.; Traniello, J.F.A. Temporal polytheism in incipient colonies of the primitive termite Zootermopsis angusticollis: A single multiage caste. J. Insect Behav. 1993, 6, 237–252. [Google Scholar] [CrossRef]
- Traniello, J.F.A.; Rosengaus, R.B.; Savoie, K. The development of immunity in a social insect: Evidence for the group facilitation of disease resistance. Proc. Nat. Acad. Sci. USA 2002, 99, 6838–6842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, S.; Pull, C.D.; Fürst, M.A. Social immunity: Emergence and evolution of colony-level disease protection. Annu. Rev. Entomol. 2018, 63, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Myllymäki, H.; Valanne, S.; Rämet, M. The Drosophila Imd signaling pathway. J. Immunol. 2014, 192, 3455–3462. [Google Scholar] [CrossRef] [Green Version]
- Rosengaus, R.B.; Traniello, J.F.A. Biparental care in incipient colonies of the dampwood termite Zootermopsis angusticollis Hagen (Isoptera: Termopsidae). J. Insect Behav. 1991, 4, 633–647. [Google Scholar] [CrossRef]
- Chouvenc, T.; Su, N.-Y.; Grace, J.K. Fifty years of attempted biological control of termites—Analysis of a failure. Biol. Control 2011, 59, 69–82. [Google Scholar] [CrossRef]
- Mirabito, D.; Rosengaus, R.B. A double-edged sword? The cost of proctodeal trophallaxis in termites. Insectes Soc. 2016, 63, 135–142. [Google Scholar] [CrossRef]
- Day, E.; Dear, P.H.; McCaughan, F. Digital PCR strategies in the development and analysis of molecular biomarkers for personalized medicine. Methods 2013, 59, 101–107. [Google Scholar] [CrossRef]
- Taylor, S.C.; Laperriere, G.; Germain, H. Droplet digital PCR versus qPCR for gene expression analysis with low abundant targets: From variable nonsense to publication quality data. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, G.; Garvey, A.; Croke, M.; Kavanagh, K. Innate humoral immune defences in mammals and insects: The same, with differences? Virulence 2018, 9, 1625–1639. [Google Scholar] [CrossRef] [Green Version]
- Terrapon, N.; Li, C.; Robertson, H.M.; Ji, L.; Meng, X.; Booth, W.; Chen, Z.; Childers, C.P.; Glastad, K.M.; Gokhale, K.; et al. Molecular traces of alternative social organization in a termite genome. Nat. Commun. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basics: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L. Evolutionary conservation of amino acid composition in paralogous insect vitellogenin. Gene 2010, 467, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haine, E.R.; Moret, Y.; Siva-Jothy, M.T.; Rolff, J. Antimicrobial defense and persistent infection in insects. Science 2008, 322, 1257–1259. [Google Scholar] [CrossRef]
- Moret, Y.; Schmid-Hempel, P. Survival for immunity: The price of immune system activation for bumblebee workers. Science 2000, 290, 1166–1168. [Google Scholar] [CrossRef]
- Sleigh, J. Antibiotic resistance in Serratia Marcescens. Br. Med. J. 1983, 286, 1651–1653. [Google Scholar] [CrossRef] [Green Version]
- Thorne, B.L.; Traniello, J.F.A. Comparative social biology of basal taxa of ants and termites. Annu. Rev. Entomol. 2003, 48, 283–306. [Google Scholar] [CrossRef] [Green Version]
- Inward, D.; Beccaloni, G.; Eggleton, P. Death of an order: A comprehensive molecular phylogenetic study confirms that termites are eusocial cockroaches. Biol. Lett. 2007, 3, 331–335. [Google Scholar] [CrossRef]
- Moeller, M.E.; Danielsen, E.T.; Herder, R.; O’Connor, M.B.; Rewitz, K.F. Dynamic feedback circuits function as a switch for shaping a maturation-inducing steroid pulse in Drosophila. Development 2013, 140, 4730–4739. [Google Scholar] [CrossRef] [Green Version]
- Neyen, C.; Runchel, C.; Schüpfer, F.; Meier, P.; Lemaitre, B. The regulatory isoform rPGRP-LC induces immune resolution via endosomal degradation of receptors. Nat. Immunol. 2016, 17, 1150–1160. [Google Scholar] [CrossRef] [Green Version]
- Wigby, S.; Domanitskaya, E.V.; Choffat, Y.; Kubli, E.; Chapman, T. The effect of mating on immunity can be masked by experimental piercing in female Drosophila melanogaster. J. Insect Physiol. 2008, 54, 414–420. [Google Scholar] [CrossRef]
- Krautz, R.; Arefin, B.; Theopold, U. Damage signals in the insect immune response. Front. Plant Sci. 2014, 5, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Riddell, C.E.; Garces, J.D.L.; Adams, S.; Barribeau, S.M.; Twell, D.; Mallon, E.B. Differential gene expression and alternative splicing in insect immune specificity. BMC Genom. 2014, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.; Eleftherianos, I. Memory and specificity in the insect immune system: Current perspectives and future challenges. Front. Immunol. 2017, 8, 1–6. [Google Scholar] [CrossRef]
- Dhinaut, J.; Chogne, M.; Moret, M. Immune priming specificity within and across generations reveals the range of pathogens affecting evolution of immunity in an insect. J. Anim. Ecol. 2017, 87, 448–463. [Google Scholar] [CrossRef] [Green Version]
- Sanda, N.B.; Hou, B.; Muhammad, A.; Ali, H.; Hou, Y. Exploring the role of Relish on antimicrobial peptide expressions (AMPs) upon nematode-bacteria complex challenge in the Nipa Palm Hispid Beetle, Octodonta nipae Maulik (Coleoptera: Chrysomelidae). Front. Microbiol. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Lamberty, M.; Zachary, D.; Lanot, R.; Bordereau, C.; Robert, A.; Hoffmann, J.A.; Bulet, P. Insect immunity. J. Biol. Chem. 2001, 276, 4085–4092. [Google Scholar] [CrossRef] [Green Version]
- Blum, M.S.; Hilker, M. Chemical protection of insect eggs. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell Publishing: Berlin, Germany; Vienna, Austria, 2002; pp. 61–90. [Google Scholar]
- Matsuura, K.; Tamura, T.; Kobayashi, N.; Yashiro, T.; Tatsumi, S. The antibacterial protein lysozyme identified as the termite egg recognition pheromone. PLoS ONE 2007, 2, e813. [Google Scholar] [CrossRef] [Green Version]
- Bulmer, M.S.; Bachelet, I.; Raman, R.; Rosengaus, R.B.; Sasisekharan, R. Targeting an antimicrobial effector function in insect immunity as a pest control strategy. Proc. Natl. Acad. Sci. USA 2009, 106, 12652–12657. [Google Scholar] [CrossRef] [Green Version]
- Gorman, M.J.; Kankanala, P.; Kanost, M.R. Bacterial challenges stimulates innate immune responses in exta-embryonic tissues of tobacco hornworm eggs. Insect Mol. Biol. 2004, 13, 19–24. [Google Scholar] [CrossRef]
- Reed, D.A.; Luhring, K.A.; Stafford, C.A.; Hansen, A.K.; Miller, J.G.; Hanks, L.M.; Paine, T.D. Host defensive response against an egg parasitoid involves cellular encapsulation and melanization. Biol. Control 2007, 41, 214–222. [Google Scholar] [CrossRef]
- Abdel-latief, M.; Hilker, M. Innate immunity: Eggs of Manduca sexta are able to respond to parasitism by Trichogramma evanescens. Insect Biochem. Mol. Biol. 2008, 38, 136–145. [Google Scholar] [CrossRef]
- Jacobs, C.G.C.; Gallagher, J.D.; Evison, S.E.F.; Heckel, D.G.; Vilcinskas, A.; Vogel, H. Endogenous egg immune defenses in the yellow mealworm beetle (Tenebrio molitor). Dev. Comp. Immunol. 2017, 70, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, C.G.C.; Spaink, H.P.; van der Zee, M. The extraembryonic serosa is a frontier epithelium providing the insect egg with a full-range innate immune response. eLife 2014, 3, 1–21. [Google Scholar] [CrossRef]
- Vlisidou, I.; Dowling, A.J.; Evans, I.R.; Waterfield, N.; French-Constant, R.H.; Wood, W. Drosophila embryos as model systems for monitoring bacterial infection in real time. PLoS Pathog. 2009, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Salmela, H.; Sundström, L. Vitellogenin in inflammation and immunity in social insects. Inflamm. Cell Signal. 2017, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Zhang, S. Immune-relevant and antioxidant activities of vitellogenin and yolk proteins in fish. Nutrients 2015, 7, 8818–8829. [Google Scholar] [CrossRef] [Green Version]
- Fiołka, M.J.; Ptaszyńska, A.A.; Czarniawski, W. Antibacterial and antifungal lysozyme-type activity in Cameraria ohridella pupae. J. Invertebr. Pathol. 2005, 90, 1–9. [Google Scholar] [CrossRef]
- Sowa-Jasiłek, A.; Zdybicka-Barabas, A.; Stączek, S.; Wydrych, J.; Skrzypiec, K.; Mak, P.; Deryło, K.; Tchórzewski, M.; Cytryńska, M. Galleria mellonella lysozyme induces apoptic changes in Candida albicans cells. Microbiol. Res. 2016, 193, 121–131. [Google Scholar] [CrossRef]
- Lo, N.; Li, B.; Ujvari, B. DNA methylation in the termite Coptoteremes lacteus. Insectes Soc. 2012, 59, 257–261. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parental Treatment 1 | Incipient Colonies Established | Samples for Digital Droplet PCR 2 | Samples for Total Protein 3 | Samples for Antibacterial Assay 3 | |
|---|---|---|---|---|---|
| NQ + NK | 66 | 5 | 33 | 33 | |
| SQ + NK | 45 | 5 | 10 | 10 | |
| Maternal Effects | HQ + NK | 45 | -- | 15 | 15 |
| LQ + NK | 97 | 5 | 17 | 17 | |
| NQ + SK | 41 | -- | 11 | 11 | |
| Paternal Effects | NQ + HK | 43 | -- | 15 | 15 |
| NQ + LK | 78 | -- | 11 | 11 | |
| SQ + SK | 58 | -- | 21 | 21 | |
| Combined Effects | HQ + HK | 51 | -- | 12 | 12 |
| LQ + LK | 99 | -- | 11 | 11 | |
| Total | 623 | 15 | 156 | 156 |
| Locus (Accession) | Primer Sequence (5′–3′) | Amplicon (bp) | |
|---|---|---|---|
| Relish (Znev_11193) | F | TCT GCA CAC TCC TGC TTA AA | 137 |
| R | AAT CAT CAT CAC TCT CCG GC | ||
| IMD (Znev_02405) | F | GTG CAA AAT TCT CCC AGT ACA | 148 |
| R | CTC TCC AAT GTT CTC CGA CA | ||
| RPL13a (Znev_00068) | F | CAC TTC AGA GCA CCA AGC AA | 152 |
| R | ACG TTT CAA TGC TGC CTT TC |
| Control | Well Contents |
|---|---|
| + control for bacterial growth | 150 μL TSB + 50 μL of Arthrobacter suspension |
| - control | 200 μL TSB |
| - control | 50 μL embryo sonicate + 150 μL sterile water |
| - control | 200 μL sterile water |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cole, E.L.; Empringham, J.S.; Biro, C.; Thompson, G.J.; Rosengaus, R.B. Relish as a Candidate Marker for Transgenerational Immune Priming in a Dampwood Termite (Blattodae: Archeotermopsidae). Insects 2020, 11, 149. https://doi.org/10.3390/insects11030149
Cole EL, Empringham JS, Biro C, Thompson GJ, Rosengaus RB. Relish as a Candidate Marker for Transgenerational Immune Priming in a Dampwood Termite (Blattodae: Archeotermopsidae). Insects. 2020; 11(3):149. https://doi.org/10.3390/insects11030149
Chicago/Turabian StyleCole, Erin L., Jessica S. Empringham, Colette Biro, Graham J. Thompson, and Rebeca B. Rosengaus. 2020. "Relish as a Candidate Marker for Transgenerational Immune Priming in a Dampwood Termite (Blattodae: Archeotermopsidae)" Insects 11, no. 3: 149. https://doi.org/10.3390/insects11030149