The Clicking Elateroidea from Chinese Mesozoic Deposits (Insecta, Coleoptera)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
- (1)
- Antennomere 2 attached subapically to antennomere 1 (i.e., pedicel to scape).
- (2)
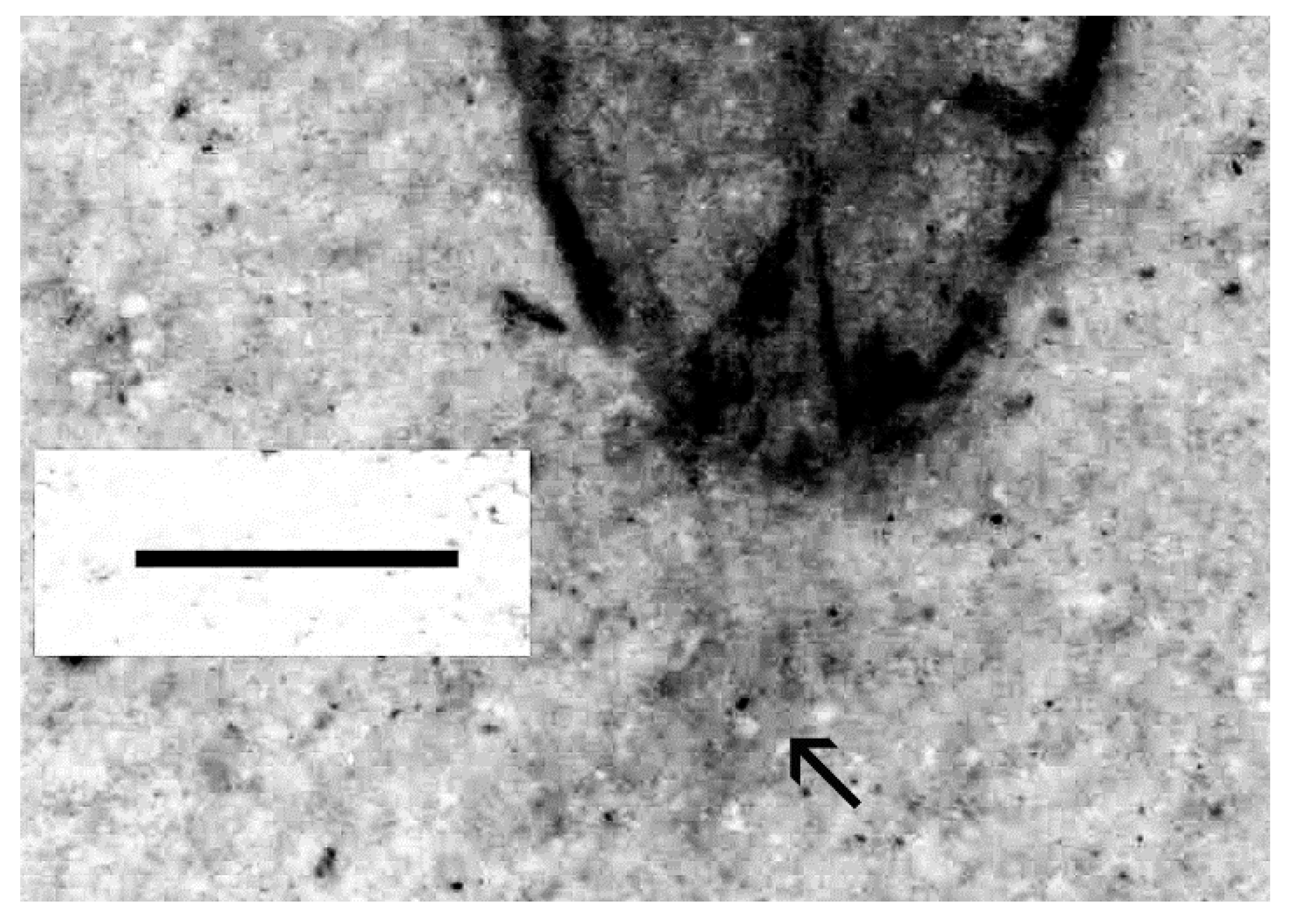
- Elytral apices have specialized enlarged pores along the first interstices, often extending towards the front along the elytra laterally or present only there (Figure 1).
- (3)
- Labrum attached underneath the expanded frontoclypeal region.
- (4)
- All visible abdominal ventrites connate.
- (5)
- Lateral margins of hypomera with antennal grooves.
- (6)
- Antennae with three enlarged apical antennomeres (i.e., flagellomeres 7 to 9).
- (7)
- One or two protibial apical spurs.
- (8)
- Male protarsomere one with sex-comb.
- (9)
- Pronotal hind angles without short sublateral carina.
- (10)
- Mesothoracic sclerites fused.
3. Results
3.1. Species Belonging to Eucnemidae
- Anoixis complanus Chang, Kirejtshuk & Ren, 2010
- Apoclion clavatus Chang, Kirejtshuk & Ren, 2010
- Apoclion antennatus Chang, Kirejtshuk & Ren, 2010
- Apoclion dolini Chang, Kirejtshuk & Ren, 2010
- Palaeoxenus sinensis Chang, Muona & Teräväinen, 2016
- Paradesmatus baiae Chang, Kirejtshuk & Ren, 2009
- Paradesmatus dilatatus Chang, Kirejtshuk & Ren, 2010
3.2. Species Belonging to Throscidae
- Archaeolus funestus Lin, 1986
- Lithomerus wunda Martin, 2010
3.3. Species Retained in Elateridae
- Artinama qinghuanensis Lin, 1986
- Bilineariselater foveatus Chang & Ren, 2008
- Cryptocoelus buffoni Dolin & Nel, 2002
- Cryptocoelus giganteus Chang, Ren & Shih, 2007
- Cryptocoelus major Dolin & Nel, 2002
- Curtelater wui Chang & Ren, 2008
- Desmatinus cognatus Chang, Kirejtshuk & Ren, 2010
- Desmatus ponomarenkoi (Chang, Kirejtshuk & Ren, 2009) NEW COMBINATION
- Paralithomerus exquisitius Chang, Zhang & Ren, 2008
- Paralithomerus parallelus Chang, Zhang & Ren, 2008
- Paraprotagrypnus superbus Zhang, Zhao & Ren, 2009
- Protagrypnus robustus Chang, Kireijtshuk & Ren, 2009
3.4. Species Showing Affinities with Eucnemidae and Elateridae
- Clavelater ningchengensis Dong et Huang, 2011
- Gripecolous enallus Lin, 1986
- Lithomerus buyssoni Dolin & Nel, 2002
3.5. Species with Uncertain Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Throscidae |
| Archaeolus funestus Lin |
| Eucnemidae |
| Schizophilinae |
| Apoclion clavatus Chang, Kiretjshuk & Ren |
| Apoclion antennatus Chang, Kiretjshuk & Ren |
| Apoclion dolini Chang, Kiretjshuk & Ren |
| Paradesmatus baiae Chang, Kiretjshuk & Ren |
| Paradesmatus dilatatus Chang, Kiretjshuk & Ren |
| Palaeoxeninae |
| Anoixis complanus Chang, Kiretjshuk & Ren |
| Palaeoxenus sinensis Chang, Muona & Teräväinen |
| Eucnemidae/Elateridae |
| Clavelater ningchengensis Dong, Huang |
| Gripecolous enallus Lin |
| Lithomerus buyssoni Dolin, Nel |
| Elateridae |
| Protagrypninae |
| Protagrypnini |
| Paraprotagrypnus superbus Zhang, Zhao & Ren |
| Protagrypnus robustus Chang, Kirejtshuk & Ren |
| Desmatini |
| Desmatinus cognatus Chang, Kirejtshuk & Ren |
| Desmatus ponomarenkoi (Chang, Kirejtshuk & Ren) NEW COMBINATION |
| (=Paradesmatus ponomarenkoi Chang, Kirejtshuk & Ren, 2009) |
| Elateridae incertae sedis |
| Artinama qinghuanensis Lin |
| Bilineariselater foveatus Chang, Ren |
| Cryptocoelus buffoni Dolin, Nel |
| Cryptocoelus giganteus Chang, Ren & Shih |
| Cryptocoelus major Dolin, Nel |
| Curtelater wui Chang, Ren |
| Paralithomerus exquisitius Chang, Zhang & Ren |
| Paralithomerus parallelus Chang, Zhang & Ren |
| Unknown placement |
| Fengningia punctata Hong |
| Microcoleus brunneus Hong |
| Ovivagina longa Zhang |
| Sinoelaterium melanocolor Ping |
| Sinolithomerus dolini Dong, Huang |
References
- Dolin, V.G. Fossil forms of click-beetles (Elateridae, Coleoptera) from lower jurassic of Middle Asia, 1973. In Fauna and Biology of Insects of Moldavia; ‘Shtiintsa’: Kishinev, Moldova, 1973; pp. 72–82. (In Russian) [Google Scholar]
- Chang, H.; Kirejtshuk, A.G.; Ren, D.; Shih, C. First fossil click beetles from the middle jurassic of inner Mongolia, China (Coleoptera: Elateridae). Ann. Zool. 2009, 59, 7–14. [Google Scholar] [CrossRef]
- Chang, H.; Kirejtshuk, A.G.; Ren, D. New fossil elaterids (Coleoptera: Polyphaga: Elateridae) from the Jehol Biota in China. Ann. Entomol. Soc. Am. 2010, 103, 866–874. [Google Scholar] [CrossRef] [Green Version]
- Oberprieler, R.; Ashman, L.; Friese, M.; Slipinski, A. The first elateroid beetles (Coleoptera: Polyphaga: Elateroidea) from the Upper Jurassic of Australia. Zootaxa 2016, 4147, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.F.; Ślipiński, A.; Seago, A.E.; Thayer, M.K.; Newton, A.F.; Marvaldi, A.E. Phylogeny of the coleoptera based on morphological characters of adults and larvae. Ann. Zool. 2011, 61, 1–217. [Google Scholar] [CrossRef]
- Muona, J. Review of the phylogeny, classification and biology of the family Eucnemidae (Coleoptera). Entomol. Scand. Suppl. 1993, 43, 1–133. [Google Scholar]
- Lawrence, J.F.; Muona, J.; Teräväinen, M.; Ståhls, G.; Vahtera, V. Anischia, Perothops and the phylogeny of Elateroidea (Coleoptera: Elateriformia). Insect Syst. Evol. 2007, 38, 205–239. [Google Scholar]
- Muona, J. Throscidae (Coleoptera) relationships, with descriptions of new fossil genera and species. Zootaxa 2019, 4576, 521–543. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Slipinski, A.; Lawrence, J.F.; Yan, E.; Ren, D.; Pang, H. Reconciling past and present: Mesozoic fossil record and a new phylogeny of the family Cerophytidae (Coleoptera: Elateroidea). Cretac. Res. 2019, 99, 51–70. [Google Scholar] [CrossRef]
- Douglas, H. Phylogenetic relationships of Elateridae inferred from adult morphology, with special reference to the position of Cardiophorinae. Zootaxa 2011, 2900, 1–45. [Google Scholar] [CrossRef]
- Calder, A. Click beetles. Genera of the Australian Elateridae (Coleoptera). In Monographs of Invertebrate Taxonomy; CSIRO: Canberra, Australia, 1996; Volume 2, 401p. [Google Scholar]
- Muona, J.; Teräväinen, M. Notes on the biology and morphology of click-beetle larvae (Coleoptera, Eucnemidae). Coleopt. Bull. 2008, 62, 475–479. [Google Scholar] [CrossRef]
- McKenna, D.; Wild, A.L.; Kanda, K.; Bellamy, C.L.; Beutel, R.G.; Caterino, M.S.; Farnum, C.W.; Hawks, D.L.; Ivie, M.A.; Jameson, M.L.; et al. The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution. Syst. Entomol. 2015, 40, 835–880. [Google Scholar] [CrossRef] [Green Version]
- Kusy, D.; Motyka, M.; Andujar, C.; Bocek, M.; Masek, M.; Sklenarova, K.; Kokas, F.; Bocakova, M.; Vogler, A.; Bocak, L. Genome sequencing of Rhinorhipus Lawrence exposes an early branch of the Coleoptera. Front. Zool. 2018, 15, 21. [Google Scholar] [CrossRef] [PubMed]
- Kusy, D.; Motyka, M.; Bocek, M.; Vogler, A.; Bocak, L. Genome sequences identify three families of Coleoptera as morphologically derived click beetles (Elateridae). Sci. Rep. 2018, 8, 17084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.Q.; Che, L.H.; Li, Y.; Liang, D.; Pang, H.; Ślipiński, A.; Zhang, P. Evolutionary history of Coleoptera revealed by extensive sampling of genes and species. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- McKenna, D.S.; Shin, D.; Ahrens, M.; Balke, C.; Beza-Beza, D.; Clarke, A.; Donath, H.; Escalona, F.; Friedrich, H.; Letsch, S.; et al. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 24729–24737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalev, A.; Kirejtshuk, A.G.; Azar, D. The oldest representatives of the family Throscidae (Coleoptera: Elateriformia) from the Lower Cretaceous Lebanese amber. Cretac. Res. 2013, 44, 157–165. [Google Scholar] [CrossRef]
- Chang, H.; Kirejtshuk, A.G.; Ren, D. On taxonomy and distribution of fossil Cerophytidae (Coleoptera: Elateriformia) with description of a new Mesozoic species of Necromera Martynov, 1926. Ann. Soc. Entomol. Fr. (N.S.) 2011, 47, 33–44. [Google Scholar] [CrossRef]
- Muona, J.; Teräväinen, M. A Re-assessment of the Eucnemidae larval characters. Papéis Avulsos Zool. 2020, 60, e202060. [Google Scholar] [CrossRef]
- Muona, J. A revision of the Indomalesian tribe Galbitini new tribe (Coleoptera. Eucnemidae). Entomol. Scand. Suppl. 1991, 39, 1–67. [Google Scholar]
- Otto, R.; Gruber, J. Eucnemid larvae of the Nearctic region. Part VI: Descriptions of the fifth instar and prepupal larval stages Stethon pectorosus LeConte, (Coleoptera: Eucnemidae: Eucneminae: Mesogenini), with notes on their biology. Insecta Mundi 2016, 474, 1–11. [Google Scholar]
- Chang, H.; Muona, J.; Pu, H.; Xu, L.; Wang, C.; Teräväinen, M.; Ren, D.; Yang, Q.; Zhang, X.; Jia, S. Chinese Cretaceous larva exposes a southern Californian living fossil (Insecta, Coleoptera, Eucnemidae). Cladistics 2016, 32, 211–214. [Google Scholar] [CrossRef]
- Dong, F.; Cai, C.; Huang, D. Revision of five Mesozoic beetles from Southern China. Acta Paleontol. Sin. 2011, 50, 481–491. [Google Scholar]
- Martin, S.K. Early Jurassic coleopterans from the Mintaja Insect Locality, Western Australia. Acta Geol. Sin. 2010, 84, 925–953. [Google Scholar] [CrossRef]
- Dolin, V.G.; Nel, A. Trois nouveaux fossiles du Mésozoïque supérieur de Chine (Coleoptera). Bull. Soc. Entomol. Fr. 2002, 107, 341–346. [Google Scholar]
- Chang, H.; Zhao, Y.; Ren, D. New fossil elaterids (Insect: Coleoptera: Polyphaga: Elateridae) from the Middle Jurassic of Inner Mongolia, China. Prog. Nat. Sci. 2009, 19, 1433–1437. [Google Scholar] [CrossRef]
- Ren, D.; Shih, C.; Gao, T.; Yao, Y.; Zhao, Y. Silent Stories—Insect Fossil Treasures from Dinosaur Era of the Northeastern China; Science Press: Beijing, China, 2010; 322p. [Google Scholar]
- Yu, Y.; Liu, Z.; Shih, C.; Ren, D. Coleoptera—Beetles. In Rhythms of Insect Evolution. Evidence from the Jurassic and Cretaceous in Northern China; Ren, D., Shih, C., Gao, T., Wang, Y., Yao, Y., Eds.; Wiley Blackwell: Hoboken, NJ, USA, 2019; Chapter 20; pp. 337–428. [Google Scholar]
- Dong, F.; Huang, D. A new elaterid from the Middle Jurassic Daohugou Biota (Coleoptera: Elateridae: Protagrypninae). Acta Geol. Sin. 2011, 85, 1224–1230. [Google Scholar]
- Zheng, D.; Chang, S.; Wang, H.; Fang, Y.; Wang, J.; Feng, C.; Xie, G.; Jarzembowski, E.A.; Zhang, H.; Wang, B. Middle-Late Triassic insect radiation revealed by diverse fossils and isotopic ages from China. Sci. Adv. 2008, 4, eaat1380. [Google Scholar] [CrossRef] [Green Version]
- Kundrata, R.; Packova, G.; Hoffmannova, J. Fossil genera in Elateridae (Insecta, Coleoptera): A triassic origin and jurassic diversification insects. Insects 2020, 11, 394. [Google Scholar] [CrossRef]
- Kundrata, R.; Bocakova, M.; Bocak, L. The comprehensive phylogeny of the superfamily Elateroidea (Coleoptera: Elateriformia). Mol. Phylogenet. Evol. 2014, 76, 162–171. [Google Scholar] [CrossRef]
- Li, Y.D.; Tihelka, E.; Liu, Z.H.; Huang, D.; Cai, C.Y. Muonabuntor gen. nov., a new genus of false click beetles from mid-Cretaceous Burmese amber (Coleoptera: Elateroidea: Eucnemidae). Palaeoentomology 2020, 003, 399–406. [Google Scholar] [CrossRef]
- Whalley, P.E.S. The systematics and palaeogeography of the Lower Jurassic insects of Dorset, England. Bull. Br. Mus. Nat. Hist. Geol. 1985, 39, 107–189. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muona, J.; Chang, H.; Ren, D. The Clicking Elateroidea from Chinese Mesozoic Deposits (Insecta, Coleoptera). Insects 2020, 11, 875. https://doi.org/10.3390/insects11120875
Muona J, Chang H, Ren D. The Clicking Elateroidea from Chinese Mesozoic Deposits (Insecta, Coleoptera). Insects. 2020; 11(12):875. https://doi.org/10.3390/insects11120875
Chicago/Turabian StyleMuona, Jyrki, Huali Chang, and Dong Ren. 2020. "The Clicking Elateroidea from Chinese Mesozoic Deposits (Insecta, Coleoptera)" Insects 11, no. 12: 875. https://doi.org/10.3390/insects11120875