Mutual and Opposing Responses of Carabid Beetles and Predatory Wasps to Local and Landscape Factors in Vineyards


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Landscape Analysis
2.3. Vegetation Parameters
2.4. Arthropod Sampling
2.4.1. Cavity-Nesting Wasps
2.4.2. Carabid Beetles
2.5. Data Analysis
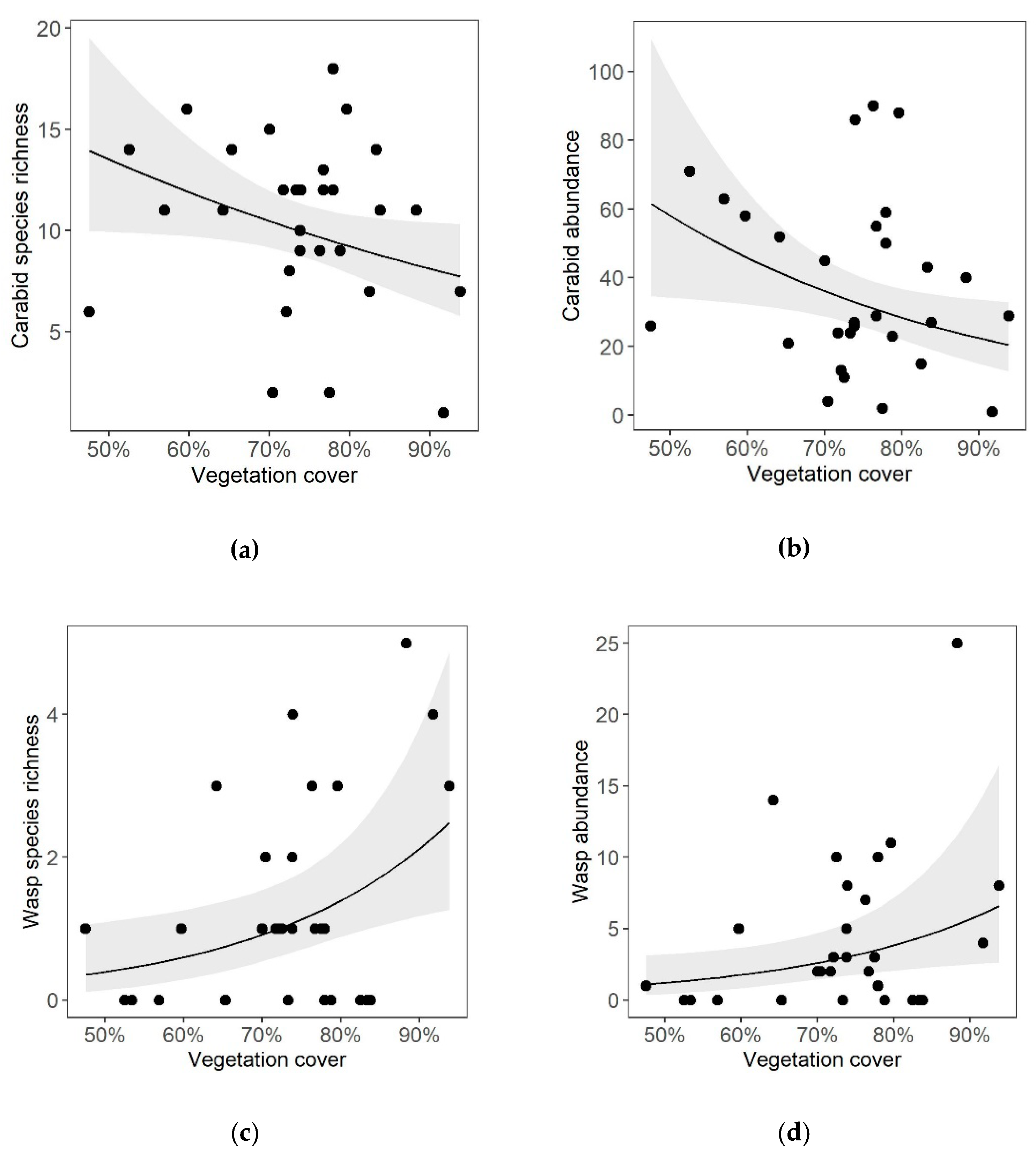
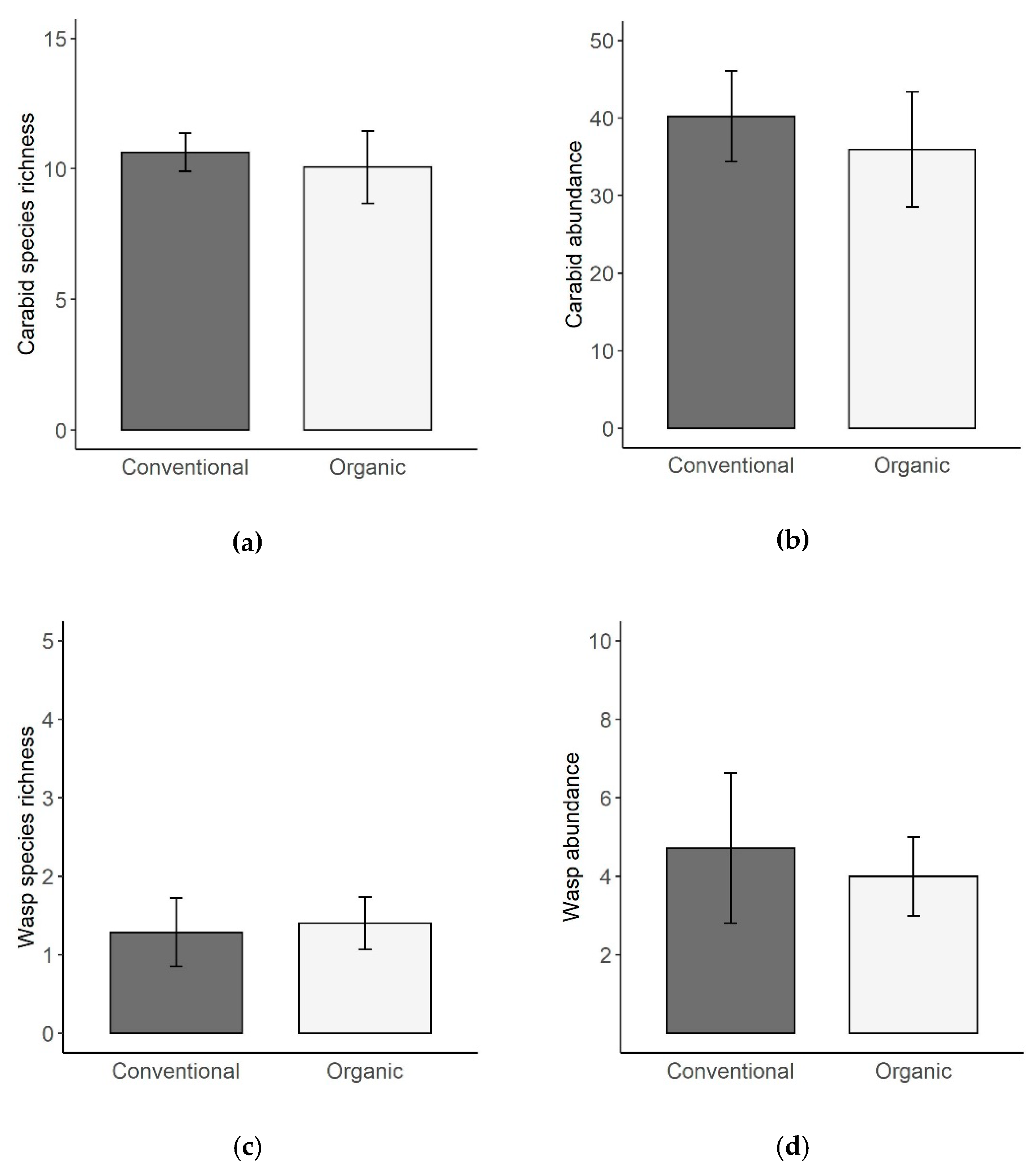
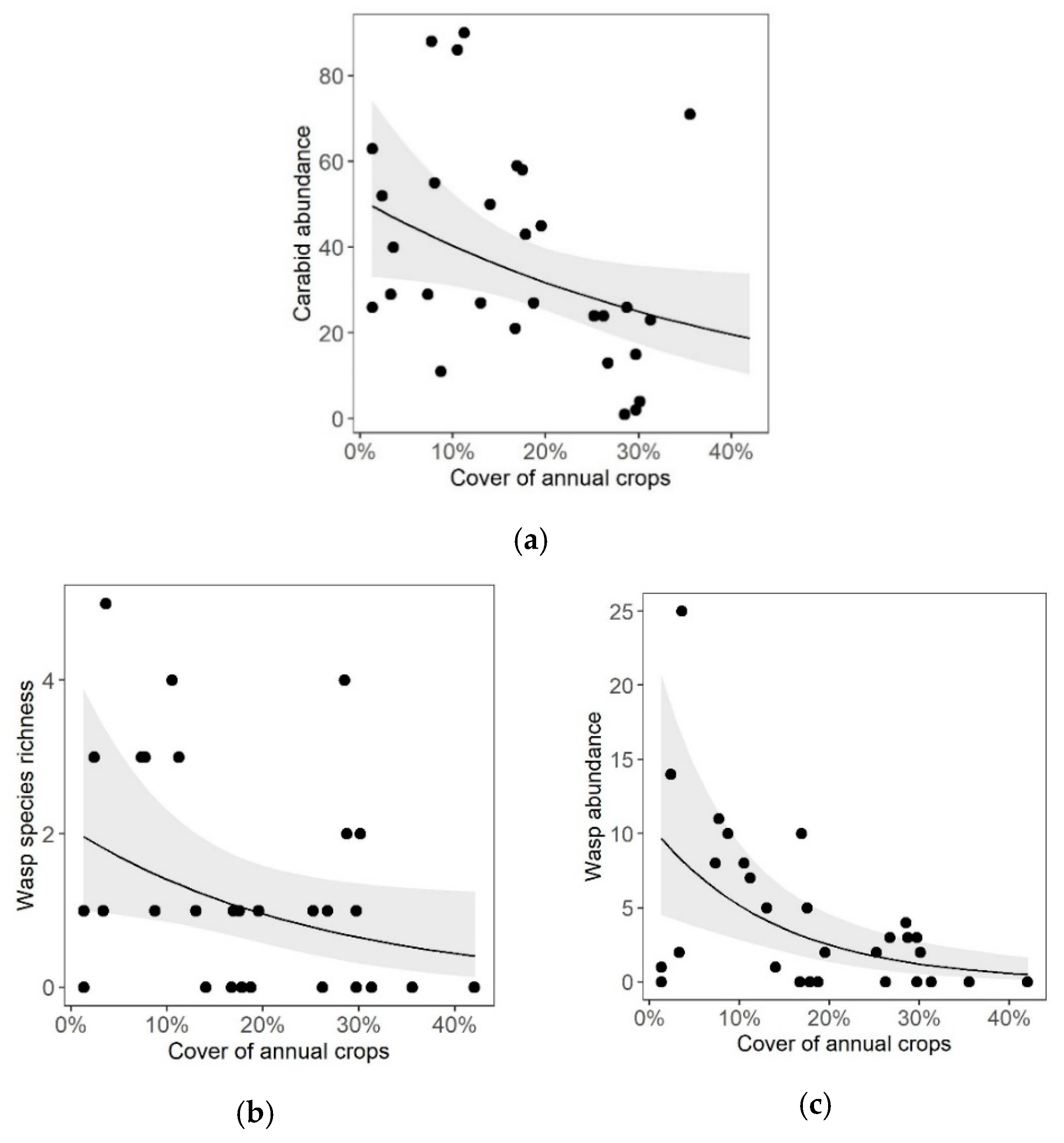
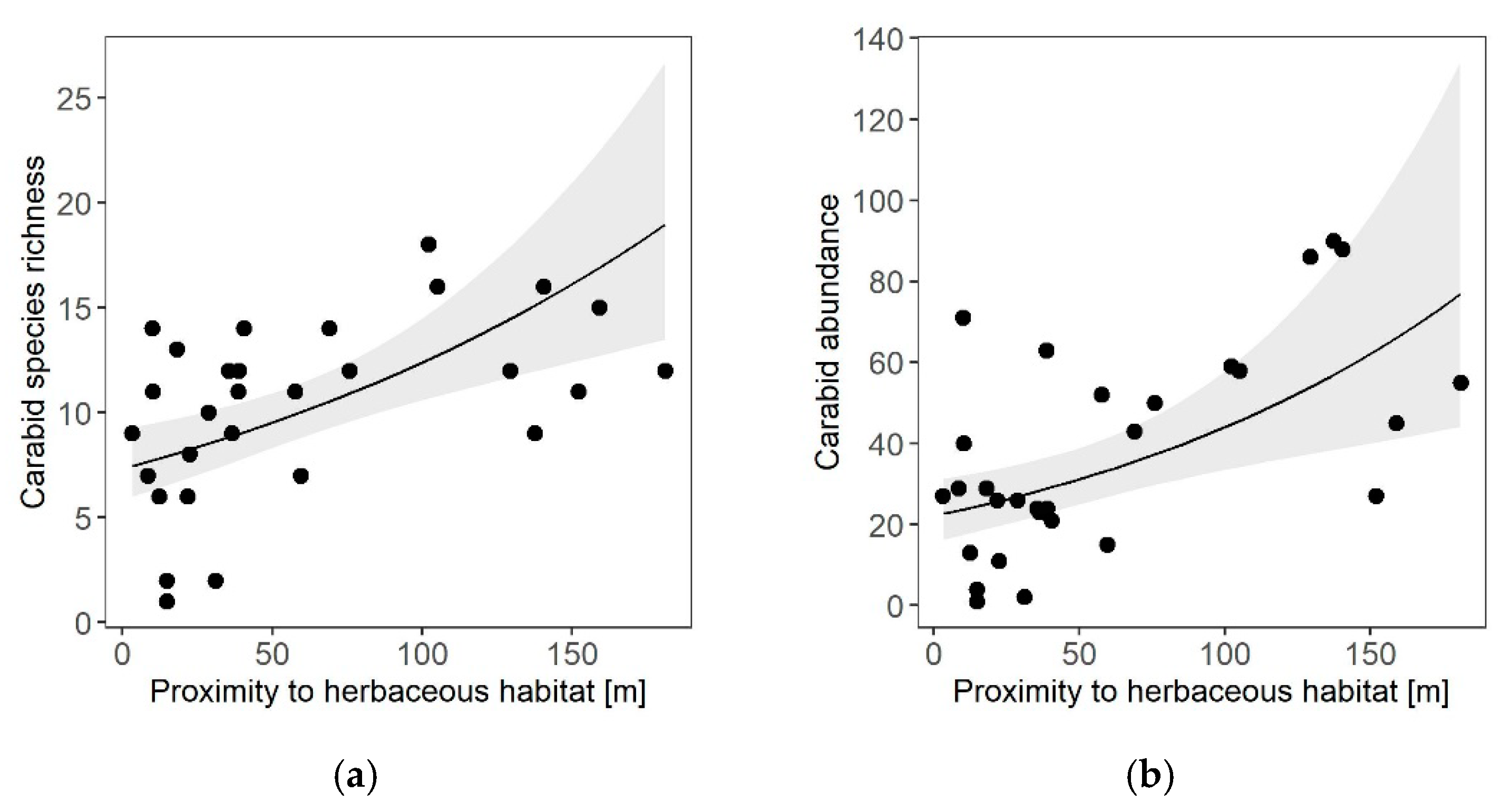
3. Results
4. Discussion
4.1. Effects of Vineyard Inter-Row Vegetation Cover on Carabid Beetles and Cavity-Nesting Wasps
4.2. Carabid Beetles and Cavity-Nesting Wasps in Organic and Conventional Vineyards
4.3. Landscape Effects on Carabid Beetles and Cavity-Nesting Wasps
4.4. Relative Biocontrol Potential of Cavity-Nesting Wasps in Viticulture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Management System | Total | |
|---|---|---|---|---|
| Conventional | Organic | |||
| Crabronidae | Trypoxylon attentuatum-deceptorium | 3 | 0 | 3 |
| Trypoxylon clavicerum | 3 | 1 | 4 | |
| Trypoxylon figulus | 11 | 8 | 19 | |
| Trypoxylon spec. | 6 | 7 | 13 | |
| Pompilidae | Dipogon spec. | 4 | 1 | 5 |
| Dipogon subintermedium | 2 | 1 | 3 | |
| Vespidae | Allodynerus rossii | 1 | 0 | 1 |
| Ancistrocerus antilope | 0 | 13 | 13 | |
| Ancistrocerus claripennis | 1 | 4 | 5 | |
| Ancistrocerus gazella | 4 | 2 | 6 | |
| Ancistrocerus nigricornis | 6 | 0 | 6 | |
| Ancistrocerus parietinus | 22 | 17 | 45 | |
| Discoelius doufourii | 0 | 1 | 1 | |
| Discoelius zonalis | 1 | 1 | 2 | |
| Microdynerus longicollis | 0 | 3 | 3 | |
| Microdynerus timidus | 2 | 3 | 5 | |
| Sum | 66 | 62 | 128 | |
| Family | Species | Management System | Total | |
|---|---|---|---|---|
| Conventional | Organic | |||
| Carabidae | Acupalpus interstitialis | 2 | 2 | 4 |
| Acupalpus meridianus | 3 | 8 | 11 | |
| Amara aenea | 109 | 50 | 159 | |
| Amara anthobia | 12 | 12 | ||
| Amara cf convexior | 1 | 1 | ||
| Amara familiaris | 3 | 5 | 8 | |
| Amara lunicollis | 1 | 1 | ||
| Amara ovata | 2 | 11 | 13 | |
| Amara similata | 1 | 2 | 3 | |
| Anchomenus dorsalis | 1 | 1 | 2 | |
| Anisodactylus binotatus | 1 | 1 | ||
| Asaphidion flavipes | 1 | 1 | ||
| Badister bullatus | 1 | 1 | ||
| Badister sodalis | 2 | 2 | ||
| Bembidion lampros | 7 | 10 | 17 | |
| Bembidion properans | 3 | 3 | ||
| Bembidion quadrimaculatum | 1 | 1 | ||
| Brachinus crepitans | 10 | 31 | 41 | |
| Brachinus explodens | 60 | 75 | 135 | |
| Callistus lunatus | 2 | 2 | ||
| Carabus auratus | 6 | 8 | 14 | |
| Carabus monilis | 1 | 1 | ||
| Cicindela campestris | 10 | 1 | 11 | |
| Harpalus affinis | 46 | 31 | 77 | |
| Harpalus atratus | 3 | 3 | ||
| Harpalus dimidiatus | 18 | 23 | 41 | |
| Harpalus distinguendus | 83 | 24 | 107 | |
| Harpalus pumilus | 6 | 13 | 19 | |
| Harpalus subcylindricus | 1 | 1 | ||
| Harpalus tardus | 26 | 40 | 66 | |
| Leistus spinibaris | 3 | 1 | 4 | |
| Microlestes maurus | 13 | 16 | 29 | |
| Microlestes minutulus | 32 | 47 | 79 | |
| Nebria brevicollis | 78 | 81 | 159 | |
| Nebria salina | 4 | 7 | 11 | |
| Notiophilus aestuans | 4 | 5 | 9 | |
| Ophonus azureus | 24 | 16 | 40 | |
| Platynus dorsalis | 2 | 7 | 9 | |
| Poecilus cupreus | 3 | 4 | 7 | |
| Semiophonus signaticornis | 1 | 1 | ||
References
- Díaz, S.; Settele, J.; Brondízio, E.; Ngo, H.T.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Butchart, S.; et al. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES: Bonn, Germany, 2019. [Google Scholar]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Ambarl, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, C.; Linsenmair, K.E.; et al. Arthropod decline in grasslands and forests is associated with drivers at landscape level. Nature 2019, 574, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Kok, M.T.J.; Alkemade, R.; Bakkenes, M.; van Eerdt, M.; Janse, J.; Mandryk, M.; Kram, T.; Lazarova, T.; Meijer, J.; van Oorschot, M.; et al. Pathways for agriculture and forestry to contribute to terrestrial biodiversity conservation: A global scenario-study. Biol. Conserv. 2018, 221, 137–150. [Google Scholar] [CrossRef]
- Navarro, A.; López-Bao, J.V. Towards a greener Common Agricultural Policy. Nat. Ecol. Evol. 2018, 2, 1830–1833. [Google Scholar] [CrossRef]
- Habel, J.C.; Samways, M.J.; Schmitt, T. Mitigating the precipitous decline of terrestrial European insects: Requirements for a new strategy. Biodivers. Conserv. 2019. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.; Entling, M.H.; Moonen, A.-C.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Holland, J.M.; Douma, J.C.; Crowley, L.; James, L.; Kor, L.; Stevenson, D.R.W.; Smith, B.M. Semi-natural habitats support biological control, pollination and soil conservation in Europe. A review. Agron. Sustain. Dev. 2017, 37, 31. [Google Scholar] [CrossRef] [Green Version]
- Bruggisser, O.T.; Schmidt-Entling, M.H.; Bacher, S. Effects of vineyard management on biodiversity at three trophic levels. Biol. Conserv. 2010, 143, 1521–1528. [Google Scholar] [CrossRef] [Green Version]
- Viers, J.H.; Williams, J.N.; Nicholas, K.A.; Barbosa, O.; Kotzé, I.; Spence, L.; Webb, L.B.; Merenlender, A.; Reynolds, M. Vinecology: Pairing wine with nature. Conserv. Lett. 2013, 6, 287–299. [Google Scholar] [CrossRef]
- Winter, S.; Bauer, T.; Strauss, P.; Kratschmer, S.; Paredes, D.; Popescu, D.; Landa, B.; Guzmán, G.; Gómez, J.A.; Guernion, M.; et al. Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: A meta-analysis. J. Appl. Ecol. 2018, 55, 2484–2495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Košulič, O.; Michalko, R.; Hula, V. Recent artificial vineyard terraces as a refuge for rare and endangered spiders in a modern agricultural landscape. Ecol. Eng. 2014, 68, 133–142. [Google Scholar] [CrossRef]
- Schmitt, T.; Augenstein, B.; Finger, A. The influence of changes in viticulture management on the butterfly (Lepidoptera) diversity in a wine growing region of southwestern Germany. Eur. J. Entomol. 2008, 105, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, M.; Wratten, S.D. The importance of viticultural landscape features and ecosystem service enhancement for native butterflies in New Zealand vineyards. J. Insect Conserv. 2012, 16, 13–23. [Google Scholar] [CrossRef]
- James, D.G.; Seymour, L.; Lauby, G.; Buckley, K. J. Insect Conserv. 2015, 19, 341–348. [CrossRef]
- Nascimbene, J.; Zottini, M.; Ivan, D.; Casagrande, V.; Marini, L. Do vineyards in contrasting landscapes contribute to conserve plant species of dry calcareous grasslands? Sci. Total Environ. 2016, 546, 244–249. [Google Scholar] [CrossRef]
- Winkler, K.J.; Viers, J.H.; Nicholas, K.A. Assessing Ecosystem Services and Multifunctionality for Vineyard Systems. Front. Environ. Sci. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Márquez-García, M.; Jacobson, S.K.; Barbosa, O. Wine with a Bouquet of Biodiversity: Assessing Agricultural Adoption of Conservation Practices in Chile. Environ. Conserv. 2019, 46, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, J.; Ahnström, J.; Weibull, A.C. The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Lichtenberg, E.M.; Kennedy, C.M.; Kremen, C.; Batáry, P.; Berendse, F.; Bommarco, R.; Bosque-Pérez, N.A.; Carvalheiro, L.G.; Snyder, W.E.; Williams, N.M.; et al. A global synthesis of the effects of diversified farming systems on arthropod diversity within fields and across agricultural landscapes. Glob. Chang. Biol. 2017, 23, 4946–4957. [Google Scholar] [CrossRef] [Green Version]
- Garratt, M.P.D.; Wright, D.J.; Leather, S.R. The effects of farming system and fertilisers on pests and natural enemies: A synthesis of current research. Agric. Ecosyst. Environ. 2011, 141, 261–270. [Google Scholar] [CrossRef]
- Winqvist, C.; Ahnström, J.; Bengtsson, J. Effects of organic farming on biodiversity and ecosystem services: Taking landscape complexity into account. Ann. N. Y. Acad. Sci. 2012, 1249, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Tuck, S.L.; Winqvist, C.; Mota, F.; Ahnström, J.; Turnbull, L.A.; Bengtsson, J. Land-use intensity and the effects of organic farming on biodiversity: A hierarchical meta-analysis. J. Appl. Ecol. 2014, 51, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Birkhofer, K.; Ekroos, J.; Corlett, E.B.; Smith, H.G. Winners and losers of organic cereal farming in animal communities across Central and Northern Europe. Biol. Conserv. 2014, 175, 25–33. [Google Scholar] [CrossRef]
- Schneider, M.K.; Lüscher, G.; Jeanneret, P.; Arndorfer, M.; Ammari, Y.; Bailey, D.; Balázs, K.; Báldi, A.; Choisis, J.P.; Dennis, P.; et al. Gains to species diversity in organically farmed fields are not propagated at the farm level. Nat. Commun. 2014, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Shackelford, G.; Steward, P.R.; Benton, T.G.; Kunin, W.E.; Potts, S.G.; Biesmeijer, J.C.; Sait, S.M. Comparison of pollinators and natural enemies: A meta-analysis of landscape and local effects on abundance and richness in crops. Biol. Rev. 2013, 88, 1002–1021. [Google Scholar] [CrossRef]
- Gallé, R.; Happe, A.K.; Baillod, A.B.; Tscharntke, T.; Batáry, P. Landscape configuration, organic management, and within-field position drive functional diversity of spiders and carabids. J. Appl. Ecol. 2019, 56, 63–72. [Google Scholar] [CrossRef]
- Mestre, L.; Schirmel, J.; Hetz, J.; Kolb, S.; Pfister, S.C.; Amato, M.; Sutter, L.; Jeanneret, P.; Albrecht, M.; Entling, M.H. Both woody and herbaceous semi-natural habitats are essential for spider overwintering in European farmland. Agric. Ecosyst. Environ. 2018, 267, 141–146. [Google Scholar] [CrossRef]
- Schüepp, C.; Uzman, D.; Herzog, F.; Entling, M.H. Habitat isolation affects plant-herbivore-enemy interactions on cherry trees. Biol. Control 2014, 71, 56–64. [Google Scholar] [CrossRef]
- Dainese, M.; Montecchiari, S.; Sitzia, T.; Sigura, M.; Marini, L. High cover of hedgerows in the landscape supports multiple ecosystem services in Mediterranean cereal fields. J. Appl. Ecol. 2016, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Mauremooto, J.R.; Wratten, S.D.; Worner, S.P.; Fry, G.L.A. Permeability of hedgerows to predatory carabid beetles. Agric. Ecosyst. Environ. 1995, 52, 141–148. [Google Scholar] [CrossRef]
- Kratschmer, S.; Pachinger, B.; Schwantzer, M.; Paredes, D.; Guzmán, G.; Goméz, J.A.; Entrenas, J.A.; Guernion, M.; Burel, F.; Nicolai, A.; et al. Response of wild bee diversity, abundance, and functional traits to vineyard inter-row management intensity and landscape diversity across Europe. Ecol. Evol. 2019, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joern, A.; Laws, A.N. Ecological Mechanisms Underlying Arthropod Species Diversity in Grasslands. Annu. Rev. Entomol. 2013, 58, 19–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommaggio, D.; Peretti, E.; Burgio, G. The effect of cover plants management on soil invertebrate fauna in vineyard in Northern Italy. BioControl 2018, 63, 795–806. [Google Scholar] [CrossRef]
- Holland, J.M. The Agroecology of Carabid Beetles; Intercept Limited: Andover, UK, 2002. [Google Scholar]
- Diehl, E.; Wolters, V.; Birkhofer, K. Arable weeds in organically managed wheat fields foster carabid beetles by resource- and structure-mediated effects. Arthropod. Plant. Interact. 2012, 6, 75–82. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Stockan, J.A. Drivers of carabid functional diversity: Abiotic environment, plant functional traits, or plant functional diversity? Ecology 2014, 95, 1213–1224. [Google Scholar] [CrossRef]
- Dassou, A.G.; Tixier, P. Response of pest control by generalist predators to local-scale plant diversity: A meta-analysis. Ecol. Evol. 2016, 6, 1143–1153. [Google Scholar] [CrossRef] [Green Version]
- Ebeling, A.; Klein, A.M.; Weisser, W.W.; Tscharntke, T. Multitrophic effects of experimental changes in plant diversity on cavity-nesting bees, wasps, and their parasitoids. Oecologia 2012, 169, 453–465. [Google Scholar] [CrossRef]
- Fabian, Y.; Sandau, N.; Bruggisser, O.T.; Aebi, A.; Kehrli, P.; Rohr, R.P.; Naisbit, R.E.; Bersier, L.-F. Plant diversity in a nutshell: Testing for small-scale effects on trap nesting wild bees and wasps. Ecosphere 2014, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Maciejewski, K.; Cumming, G.S. Multi-scale network analysis shows scale-dependency of significance of individual protected areas for connectivity. Landsc. Ecol. 2016, 31, 761–774. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Duan, M.; Yu, Z.; Axmacher, J.C. Different response patterns of epigaeic spiders and carabid beetles to varying environmental conditions in fields and semi-natural habitats of an intensively cultivated agricultural landscape. Agric. Ecosyst. Environ. 2018, 264, 54–62. [Google Scholar] [CrossRef]
- Reynolds, C.; Fletcher, R.J.; Carneiro, C.M.; Jennings, N.; Ke, A.; LaScaleia, M.C.; Lukhele, M.B.; Mamba, M.L.; Sibiya, M.D.; Austin, J.D.; et al. Inconsistent effects of landscape heterogeneity and land-use on animal diversity in an agricultural mosaic: A multi-scale and multi-taxon investigation. Landsc. Ecol. 2018, 33, 241–255. [Google Scholar] [CrossRef]
- Witt, R. Wespen; Vademecum-Verlag: Oldenburg, Germany, 2009. [Google Scholar]
- Kotze, D.J.; Brandmayr, P.; Casale, A.; Dauffy-Richard, E.; Dekoninck, W.; Koivula, M.; Lovei, G.; Mossakowski, D.; Noordijk, J.; Paarmann, W.; et al. Forty years of carabid beetle research in Europe—From taxonomy, biology, ecology and population studies to bioindication, habitat assessment and conservation. Zookeys 2011, 100, 55–148. [Google Scholar] [CrossRef] [PubMed]
- Trautner, J. Die Laufk{ä}fer Baden-W{ü}rttembergs; Ulmer Eugen Verlag: Stuttgart, Germany, 2017. [Google Scholar]
- Adamski, Z.; Bufo, S.A.; Chowański, S.; Falabella, P.; Lubawy, J.; Marciniak, P.; Pacholska-Bogalska, J.; Salvia, R.; Scrano, L.; Słocińska, M.; et al. Beetles as model organisms in physiological, biomedical and environmental studies—A review. Front. Physiol. 2019, 10, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can Generalist Predators be Effective Biocontrol Agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.S.; Dosdall, L.M.; Willenborg, C.J. The Role of Ground Beetles (Coleoptera: Carabidae) in Weed Seed Consumption: A Review. Weed Sci. 2015, 63, 335–376. [Google Scholar] [CrossRef]
- Sotherton, N.W. The distribution and abundance of predatory Coleoptera overwintering in field boundaries. Ann. Appl. Biol. 1985, 106, 17–21. [Google Scholar] [CrossRef]
- Pfiffner, L.; Luka, H. Overwintering of arthropods in soils of arable fields and adjacent semi-natural habitats. Agric. Ecosyst. Environ. 2000, 78, 215–222. [Google Scholar] [CrossRef]
- Homburg, K.; Homburg, N.; Schäfer, F.; Schuldt, A.; Assmann, T. Carabids.org—A dynamic online database of ground beetle species traits (Coleoptera, Carabidae). Insect Conserv. Divers. 2014, 7, 195–205. [Google Scholar] [CrossRef]
- Gobbi, M.; Fontaneto, D.; Bragalanti, N.; Pedrotti, L.; Lencioni, V. Carabid beetle (Coleoptera: Carabidae) richness and functional traits in relation to differently managed grasslands in the Alps. Ann. la Soc. Entomol. Fr. 2015, 51, 52–59. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Klein, A.-M.; Tscharntke, T. Spatiotemporal variation in the diversity of hymenoptera across a tropical habitat gradient. Ecology 2005, 86, 3296–3302. [Google Scholar] [CrossRef]
- Holzschuh, A.; Steffan-Dewenter, I.; Tscharntke, T. Grass strip corridors in agricultural landscapes enhance nest-site colonization by solitary wasps. Ecol. Appl. 2009, 19, 123–132. [Google Scholar] [CrossRef]
- Klein, A.M.; Steffan-Dewenter, I.; Tscharntke, T. Foraging trip duration and density of megachilid bees, eumenid wasps and pompilid wasps in tropical agroforestry systems. J. Anim. Ecol. 2004, 73, 517–525. [Google Scholar] [CrossRef]
- Tscharntke, T.; Gathmann, A.; Steffan-Dewenter, I. Bioindication using trap-nesting bees and wasps and their natural enemies: Community structure and interactions. J. Appl. Ecol. 1998, 35, 708–719. [Google Scholar] [CrossRef]
- Staab, M.; Pufal, G.; Tscharntke, T.; Klein, A.-M. Trap nests for bees and wasps to analyse trophic interactions in changing environments—A systematic overview and user guide. Methods Ecol. Evol. 2018, 9, 2226–2239. [Google Scholar] [CrossRef]
- Yeo, P.F.; Corbet, S.A.; Disney, R.H.L. Solitary Wasps; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Diekötter, T.; Peter, F.; Jauker, B.; Wolters, V.; Jauker, F. Mass-flowering crops increase richness of cavity-nesting bees and wasps in modern agro-ecosystems. GCB Bioenergy 2014, 6, 219–226. [Google Scholar] [CrossRef]
- Harris, A.C. Ancistrocerus gazella (Hymenoptera: Vespoidea: Eumenidae): A potentially useful biological control agent for leafrollers Planotortrix octo, P. excessana, Ctenopseustis obliquana, C. herana, and Epiphyas postvittana (Lepidoptera: Tortricidae) in New Zealand. New Zeal. J. Crop Hortic. Sci. 1994, 22, 235–238. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I. Importance of Habitat Area and Landscape Context for Species Richness of Bees and Wasps in Fragmented Orchard Meadows. Conserv. Biol. 2003, 17, 1036–1044. [Google Scholar] [CrossRef]
- Krewenka, K.M.; Holzschuh, A.; Tscharntke, T.; Dormann, C.F. Landscape elements as potential barriers and corridors for bees, wasps and parasitoids. Biol. Conserv. 2011, 144, 1816–1825. [Google Scholar] [CrossRef]
- Uzman, D.; Pliester, J.; Leyer, I.; Entling, M.H.; Reineke, A. Drivers of entomopathogenic fungi presence in organic and conventional vineyard soils. Appl. Soil Ecol. 2019, 133, 89–97. [Google Scholar] [CrossRef]
- Kolb, S.; Uzman, D.; Leyer, I.; Reineke, A.; Entling, M.H. Differential effects of semi-natural habitats and organic management on spiders in viticultural landscapes. Agric. Ecosyst. Environ. 2020, 287, 106695. [Google Scholar] [CrossRef]
- Uzman, D.; Reineke, A.; Entling, M.H.; Leyer, I. Habitat area and connectivity support cavity-nesting bees in vineyards more than organic management. Biol. Conserv. 2020, 242, 108419. [Google Scholar] [CrossRef]
- McGourty, G.T.; Ohmart, J.; Chaney, D. Organic Winegrowing Manual; UCANR Publications: Richmond, CA, USA, 2011; Volume 3511. [Google Scholar]
- Woodcock, B.A.; Potts, S.G.; Westbury, D.B.; Ramsay, A.J.; Lambert, M.; Harris, S.J.; Brown, V.K. The importance of sward architectural complexity in structuring predatory and phytophagous invertebrate assemblages. Ecol. Entomol. 2007, 32, 302–311. [Google Scholar] [CrossRef]
- Lüdecke, D. sjstats: Statistical Functions for Regression Models, (Version 0.17.1). 2018. Available online: https://zenodo.org/record/1400701 (accessed on 30 October 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Viena, Austria, 2018. [Google Scholar]
- Wickham, H. Reshaping Data with the Reshape Package. 2007. Available online: https://www.jstatsoft.org/article/view/v021i12 (accessed on 30 October 2020).
- Wickham, H.; François, R.; Henry, L.; Müller, K. dplyr: A Grammar of Data Manipulation. 2018. Available online: https://dplyr.tidyverse.org/ (accessed on 30 October 2020).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB: Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Barton, K. MuMIn: Multi-Model Inference. 2018. Available online: https://cran.r-project.org/web/packages/MuMIn/index.html (accessed on 30 October 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Ng, K.; McIntyre, S.; Macfadyen, S.; Barton, P.S.; Driscoll, D.A.; Lindenmayer, D.B. Dynamic effects of ground-layer plant communities on beetles in a fragmented farming landscape. Biodivers. Conserv. 2018, 27, 2131–2153. [Google Scholar] [CrossRef]
- Djoudi, E.A.; Marie, A.; Mangenot, A.; Puech, C.; Aviron, S.; Plantegenest, M.; Pétillon, J. Farming system and landscape characteristics differentially affect two dominant taxa of predatory arthropods. Agric. Ecosyst. Environ. 2018, 259, 98–110. [Google Scholar] [CrossRef]
- Fiera, C.; Ulrich, W.; Popescu, D.; Bunea, C.-I.; Manu, M.; Nae, I.; Stan, M.; Markó, B.; Urák, I.; Giurginca, A.; et al. Effects of vineyard inter-row management on the diversity and abundance of plants and surface-dwelling invertebrates in Central Romania. J. Insect Conserv. 2020. [Google Scholar] [CrossRef] [Green Version]
- Greenslade, P.J.M. Pitfall Trapping as a Method for Studying Populations of Carabidae (Coleoptera). J. Anim. Ecol. 1964, 33, 301. [Google Scholar] [CrossRef]
- Thomas, C.F.G.; Brown, N.J.; Kendall, D.A. Carabid movement and vegetation density: Implications for interpreting pitfall trap data from split-field trials. Agric. Ecosyst. Environ. 2006, 113, 51–61. [Google Scholar] [CrossRef]
- Shearin, A.F.; Reberg-Horton, S.C.; Gallandt, E.R. Direct Effects of Tillage on the Activity Density of Ground Beetle (Coleoptera: Carabidae) Weed Seed Predators. Environ. Entomol. 2007, 36, 1140–1146. [Google Scholar] [CrossRef]
- Uzman, D.; (Geisenheim University, Geisenheim, Germany). Personal observation, 2020.
- Langellotto, G.A.; Denno, R.F. Responses of invertebrate natural enemies to complex-structured habitats: A meta-analytical synthesis. Oecologia 2004, 139, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rubene, D.; Schroeder, M.; Ranius, T. Diversity patterns of wild bees and wasps in managed boreal forests: Effects of spatial structure, local habitat and surrounding landscape. Biol. Conserv. 2015, 184, 201–208. [Google Scholar] [CrossRef]
- Wood, T.J.; Holland, J.M.; Goulson, D. Pollinator-friendly management does not increase the diversity of farmland bees and wasps. Biol. Conserv. 2015, 187, 120–126. [Google Scholar] [CrossRef]
- Campbell, J.; Smithers, C.; Irvin, A.; Kimmel, C.; Stanley-Stahr, C.; Daniels, J.; Ellis, J. Trap Nesting Wasps and Bees in Agriculture: A Comparison of Sown Wildflower and Fallow Plots in Florida. Insects 2017, 8, 107. [Google Scholar] [CrossRef] [Green Version]
- Danne, A.; Thomson, L.J.; Sharley, D.J.; Penfold, C.M.; Hoffmann, A.A. Effects of native grass cover crops on beneficial and pest invertebrates in Australian vineyards. Environ. Entomol. 2010, 39, 970–978. [Google Scholar] [CrossRef]
- Daane, K.M.; Hogg, B.N.; Wilson, H. Native grass ground covers provide multiple ecosystem services in Californian vineyards. J. Appl. Ecol. 2018, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Bauer, K. Weinbau; AVBuch-Austria, Agrarverlag: Wien, Austria, 2008; ISBN 978-370402284-4. [Google Scholar]
- Rundlöf, M.; Smith, H.G.; Birkhofer, K. Effects of Organic Farming on Biodiversity. In eLS; John Wiley & Sons Ltd.: Chichester, UK, 2016; pp. 1–7. ISBN 9780470015902. [Google Scholar]
- Fischer, C.; Thies, C.; Tscharntke, T. Mixed effects of landscape complexity and farming practice on weed seed removal. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 297–303. [Google Scholar] [CrossRef]
- Djoudi, E.A.; Plantegenest, M.; Aviron, S.; Pétillon, J. Local vs. landscape characteristics differentially shape emerging and circulating assemblages of carabid beetles in agroecosystems. Agric. Ecosyst. Environ. 2019, 270–271, 149–158. [Google Scholar] [CrossRef]
- Adhikari, S.; Menalled, F. Impacts of Dryland Farm Management Systems on Weeds and Ground Beetles (Carabidae) in the Northern Great Plains. Sustainability 2018, 10, 2146. [Google Scholar] [CrossRef] [Green Version]
- Purtauf, T.; Roschewitz, I.; Dauber, J.; Thies, C.; Tscharntke, T.; Wolters, V. Landscape context of organic and conventional farms: Influences on carabid beetle diversity. Agric. Ecosyst. Environ. 2005, 108, 165–174. [Google Scholar] [CrossRef]
- Birkhofer, K.; Bezemer, T.M.; Bloem, J.; Bonkowski, M.; Christensen, S.; Dubois, D.; Ekelund, F.; Fließbach, A.; Gunst, L.; Hedlund, K.; et al. Long-term organic farming fosters below and aboveground biota: Implications for soil quality, biological control and productivity. Soil Biol. Biochem. 2008, 40, 2297–2308. [Google Scholar] [CrossRef]
- Diekötter, T.; Wamser, S.; Wolters, V.; Birkhofer, K. Landscape and management effects on structure and function of soil arthropod communities in winter wheat. Agric. Ecosyst. Environ. 2010, 137, 108–112. [Google Scholar] [CrossRef]
- Caprio, E.; Nervo, B.; Isaia, M.; Allegro, G.; Rolando, A. Organic versus conventional systems in viticulture: Comparative effects on spiders and carabids in vineyards and adjacent forests. Agric. Syst. 2015, 136, 61–69. [Google Scholar] [CrossRef]
- Menalled, F.D.; Smith, R.G.; Dauer, J.T.; Fox, T.B. Impact of agricultural management on carabid communities and weed seed predation. Agric. Ecosyst. Environ. 2007, 118, 49–54. [Google Scholar] [CrossRef]
- Kromp, B. Carabid beetles (Coleoptera, Carabidae) as bioindicators in biological and conventional farming in Austrian potato fields. Biol. Fertil. Soils 1990, 9, 182–187. [Google Scholar] [CrossRef]
- Döring, T.F.; Kromp, B. Which carabid species benefit from organic agriculture?—A review of comparative studies in winter cereals from Germany and Switzerland. Agric. Ecosyst. Environ. 2003, 98, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Holzschuh, A.; Steffan-Dewenter, I.; Tscharntke, T. How do landscape composition and configuration, organic farming and fallow strips affect the diversity of bees, wasps and their parasitoids? J. Anim. Ecol. 2010, 79, 491–500. [Google Scholar] [CrossRef]
- Froidevaux, J.S.P.P.; Louboutin, B.; Jones, G. Does organic farming enhance biodiversity in Mediterranean vineyards? A case study with bats and arachnids. Agric. Ecosyst. Environ. 2017, 249, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Batáry, P.; Holzschuh, A.; Orci, K.M.; Samu, F.; Tscharntke, T. Responses of plant, insect and spider biodiversity to local and landscape scale management intensity in cereal crops and grasslands. Agric. Ecosyst. Environ. 2012, 146, 130–136. [Google Scholar] [CrossRef]
- Birkhofer, K.; Arvidsson, F.; Ehlers, D.; Mader, V.L.; Bengtsson, J.; Smith, H.G. Organic farming affects the biological control of hemipteran pests and yields in spring barley independent of landscape complexity. Landsc. Ecol. 2015, 31, 567–579. [Google Scholar] [CrossRef]
- Coudrain, V.; Herzog, F.; Entling, M.H. Effects of Habitat Fragmentation on Abundance, Larval Food and Parasitism of a Spider-Hunting Wasp. PLoS ONE 2013, 8, e59286. [Google Scholar] [CrossRef] [Green Version]
- Trichard, A.; Alignier, A.; Biju-Duval, L.; Petit, S. The relative effects of local management and landscape context on weed seed predation and carabid functional groups. Basic Appl. Ecol. 2013, 14, 235–245. [Google Scholar] [CrossRef]
- Tamburini, G.; De Simone, S.; Sigura, M.; Boscutti, F.; Marini, L. Conservation tillage mitigates the negative effect of landscape simplification on biological control. J. Appl. Ecol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Rusch, A.; Binet, D.; Delbac, L.; Thiéry, D. Local and landscape effects of agricultural intensification on Carabid community structure and weed seed predation in a perennial cropping system. Landsc. Ecol. 2016, 1–12. [Google Scholar] [CrossRef]
- Petit, S.; Trichard, A.; Biju-Duval, L.; McLaughlin, Ó.B.B.; Bohan, D.A.A. Interactions between conservation agricultural practice and landscape composition promote weed seed predation by invertebrates. Agric. Ecosyst. Environ. 2017, 240, 45–53. [Google Scholar] [CrossRef]
- Winqvist, C.; Bengtsson, J.; Aavik, T.; Berendse, F.; Clement, L.W.; Fischer, C.; Flohre, A.; Geiger, F.; Liira, J.; Thies, C.; et al. Mixed effects of organic farming and landscape complexity on farmland biodiversity and biological control potential across Europe. J. Appl. Ecol. 2011, 48, 570–579. [Google Scholar] [CrossRef]
- Rainio, J.; Niemelä, J. Ground beetles (Coleoptera: Carabidae) as bioindicators. Biodivers. Conserv. 2003, 12, 487–506. [Google Scholar] [CrossRef]
- Jonason, D.; Smith, H.G.; Bengtsson, J.; Birkhofer, K. Landscape simplification promotes weed seed predation by carabid beetles (Coleoptera: Carabidae). Landsc. Ecol. 2013, 28, 487–494. [Google Scholar] [CrossRef]
- Martin, E.A.; Reineking, B.; Seo, B.; Steffan-Dewenter, I. Natural enemy interactions constrain pest control in complex agricultural landscapes. Proc. Natl. Acad. Sci. USA 2013, 110, 5534–5539. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, U.S.; Jauker, F.; Lanzen, J.; Warzecha, D.; Wolters, V.; Diekötter, T. Prey-dependent benefits of sown wildflower strips on solitary wasps in agroecosystems. Insect Conserv. Divers. 2018, 11, 42–49. [Google Scholar] [CrossRef]
- Schüepp, C.; Herrmann, J.D.; Herzog, F.; Schmidt-Entling, M.H. Differential effects of habitat isolation and landscape composition on wasps, bees, and their enemies. Oecologia 2011, 165, 713–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muneret, L.; Auriol, A.; Thiéry, D.; Rusch, A. Organic farming at local and landscape scales fosters biological pest control in vineyards. Ecol. Appl. 2019, 29, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Mohr, H.D. Farbatlas Krankheiten, Schädlinge und Nützlinge an der Weinrebe; Ulmer: Stuttgart, Germany, 2012; ISBN 3800141485. [Google Scholar]
- Wearing, C.H.; Harris, A.C. Evaluation of the predatory wasp, Ancistrocerus gazella, for biological control of leafrollers in Otago fruit crops: 1. Prey composition, nest structure and wasp productivity from artificial nests. Biocontrol Sci. Technol. 1999, 9, 315–325. [Google Scholar] [CrossRef]
- Stern, V.M.; Johnson, J.A. Biology and Control of the Grape Bud Beetle, Glyptoscelis squamulata (Coleoptera: Chrysomelidae), in Southern California Table Grapes. J. Econ. Entomol. 1984, 77, 1327–1334. [Google Scholar] [CrossRef]
- Roberto, S.R.; Genta, W.; Ventura, M.U. Diabrotica speciosa (Ger.) (Coleoptera: Chrysomelidae): New Pest in Table Grape Orchards. Neotrop. Entomol. 2001, 30, 721–722. [Google Scholar] [CrossRef]
- Albrecht, M.; Duelli, P.; Schmid, B.; Müller, C.B. Interaction diversity within quantified insect food webs in restored and adjacent intensively managed meadows. J. Anim. Ecol. 2007, 76, 1015–1025. [Google Scholar] [CrossRef] [PubMed]




| Response Variable | Predictors | Estimate ± SE | p-Value | R²cond/marg | Distribu-tion | AICc | dAICc (to Full Model) | AICc (Full Model) |
|---|---|---|---|---|---|---|---|---|
| Carabid beetles | ||||||||
| Species richness | (Intercept) | 2.31 ± 0.07 | <2 × 10−16 (***) | 0.38/0.44 | Poisson | 167.8 | 3.9 | 171.7 |
| SNH % | 0.15 ± 0.08 | 0.05 | ||||||
| Vegetation cover % | −0.14 ± 0.07 | 0.04 (*) | ||||||
| Distance to herbaceous habitat | 0.28 ± 0.08 | <0.001 (***) | ||||||
| Abundance | (Intercept) | 3.51 ± 0.11 | <2 × 10−16 (***) | 0.50/0.50 | Negative binomial | 262.1 | 3.6 | 265.7 |
| Annual crops % | −0.27 ± 0.13 | 0.03 (*) | ||||||
| Vegetation cover % | −0.26 ± 0.12 | 0.02 (*) | ||||||
| Distance to herbaceous habitat | 0.36 ± 0.12 | 0.002 (**) | ||||||
| Cavity-nesting wasps | ||||||||
| Species richness | (Intercept) | 0.04 ± 0.25 | 0.87 | 0.37/0.50 | Conway–Maxwell | 90.4 | 3.7 | 94.1 |
| Annual crops % | −0.44 ± 0.21 | 0.04 (*) | ||||||
| Vegetation cover % | 0.47 ± 0.18 | 0.01 (*) | ||||||
| Abundance | (Intercept) | 1.07 ± 0.30 | <0.001 (***) | 0.02/0.02 | Conway–Maxwell | 141.6 | 5.6 | 147.2 |
| Annual % | −0.82 ± 0.22 | <0.001 (***) | ||||||
| Vegetation cover % | 0.43 ± 0.20 | 0.03 (*) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uzman, D.; Entling, M.H.; Leyer, I.; Reineke, A. Mutual and Opposing Responses of Carabid Beetles and Predatory Wasps to Local and Landscape Factors in Vineyards. Insects 2020, 11, 746. https://doi.org/10.3390/insects11110746
Uzman D, Entling MH, Leyer I, Reineke A. Mutual and Opposing Responses of Carabid Beetles and Predatory Wasps to Local and Landscape Factors in Vineyards. Insects. 2020; 11(11):746. https://doi.org/10.3390/insects11110746
Chicago/Turabian StyleUzman, Deniz, Martin H. Entling, Ilona Leyer, and Annette Reineke. 2020. "Mutual and Opposing Responses of Carabid Beetles and Predatory Wasps to Local and Landscape Factors in Vineyards" Insects 11, no. 11: 746. https://doi.org/10.3390/insects11110746