Vegetation Pattern Modulates Ground Arthropod Diversity in Semi-Arid Mediterranean Steppes

 , and
, and
Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. Vegetation Data
2.3. Arthropods Data
2.4. Data Analyses
3. Results
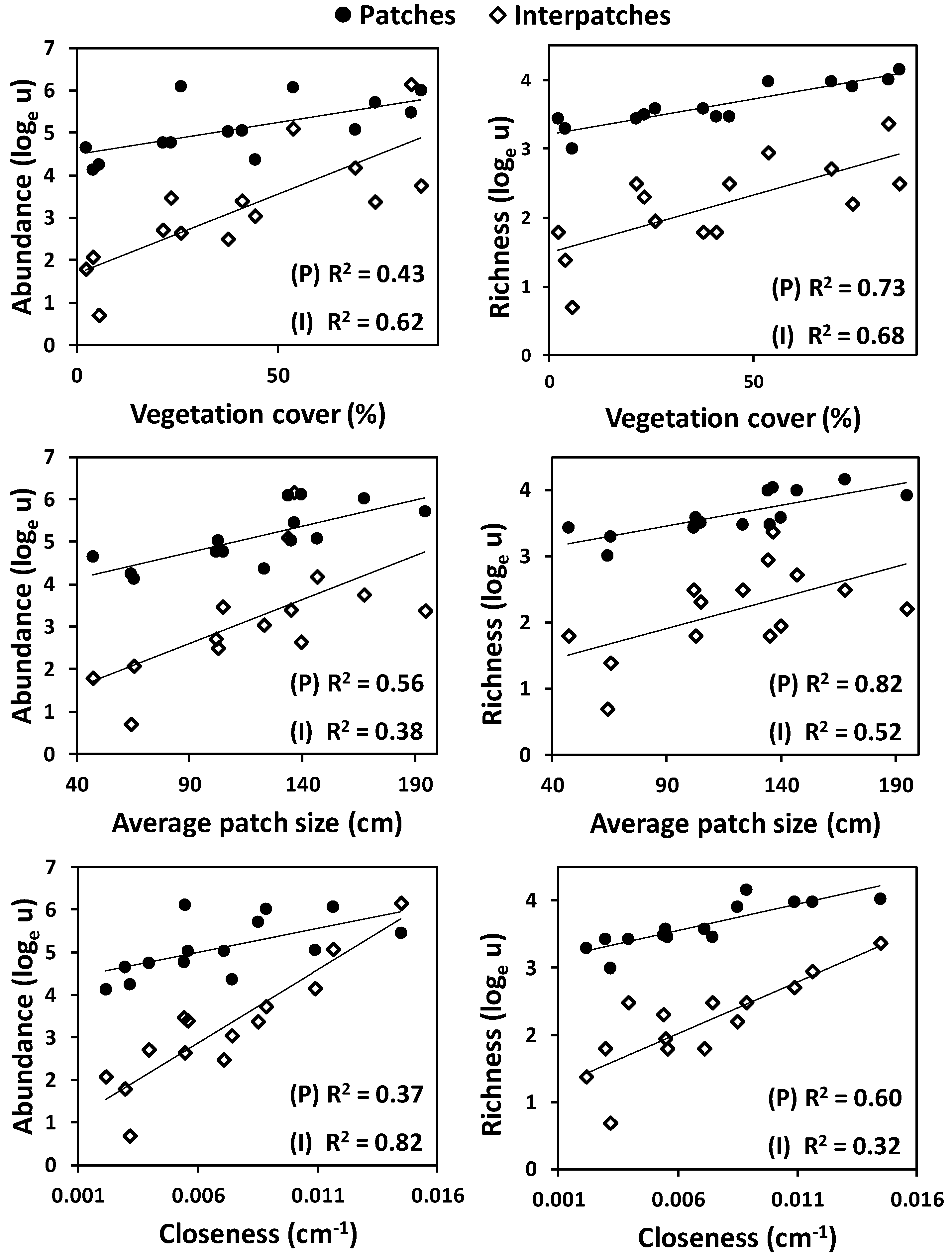
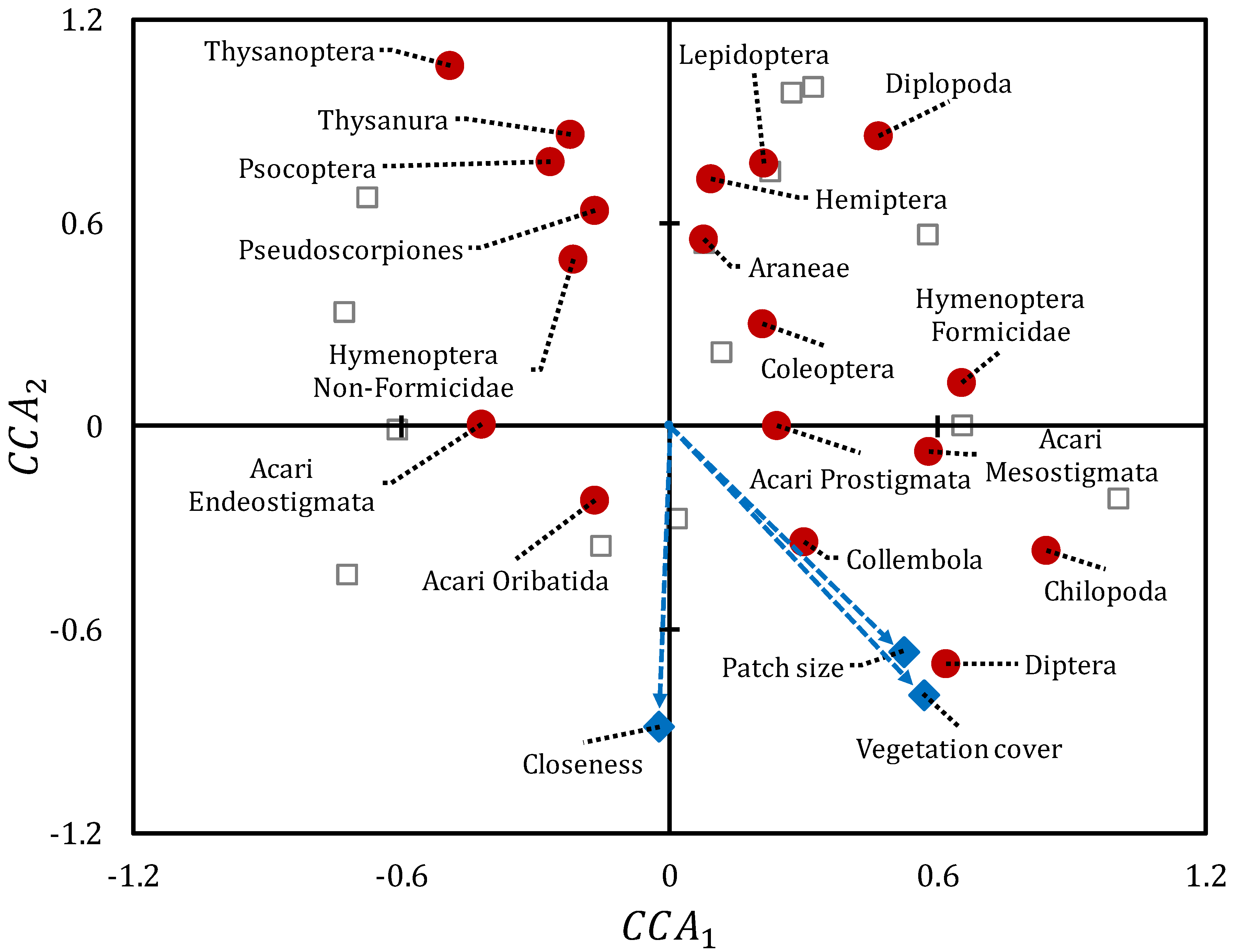
3.1. Landscape-Based Analyses
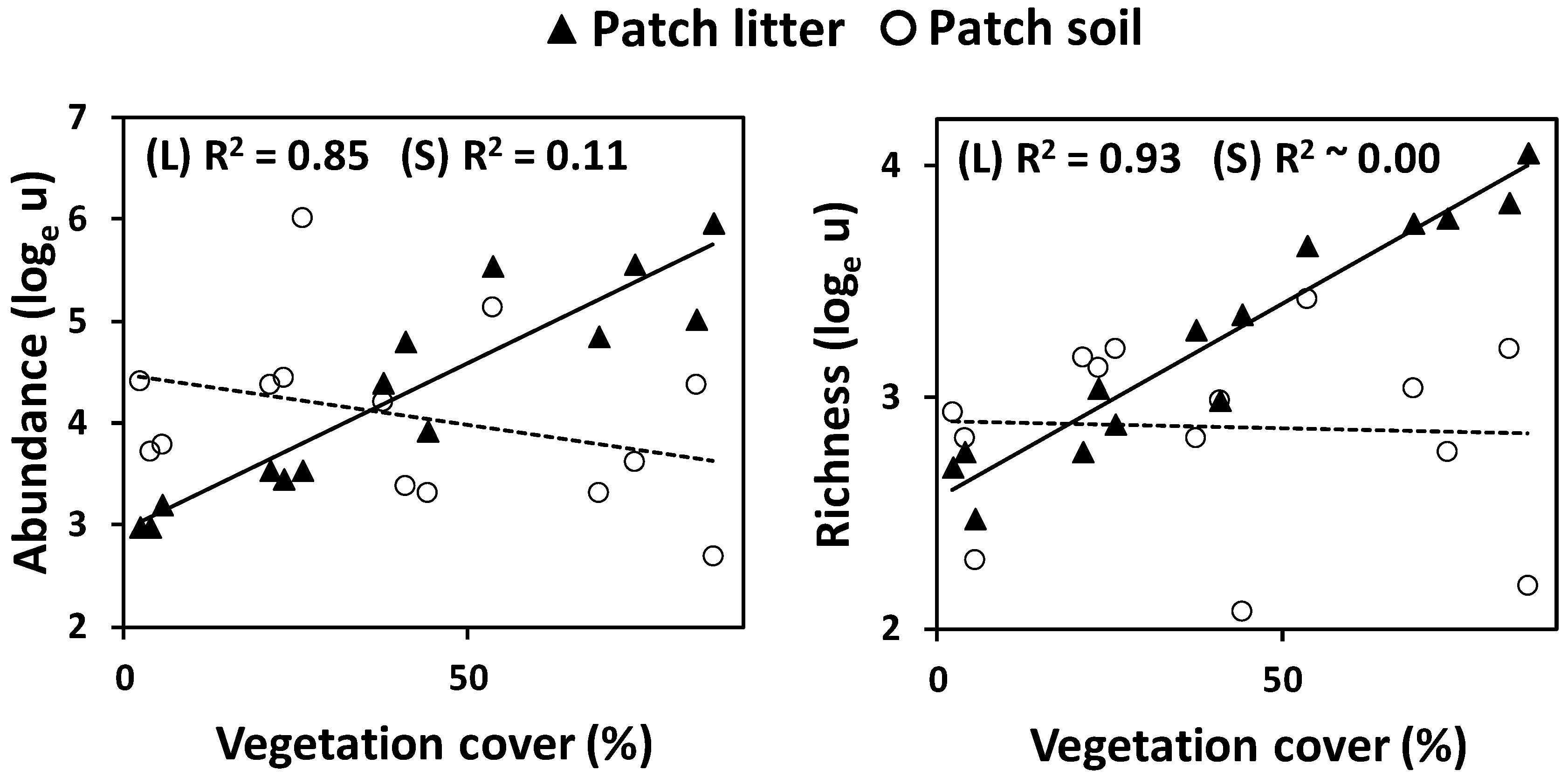
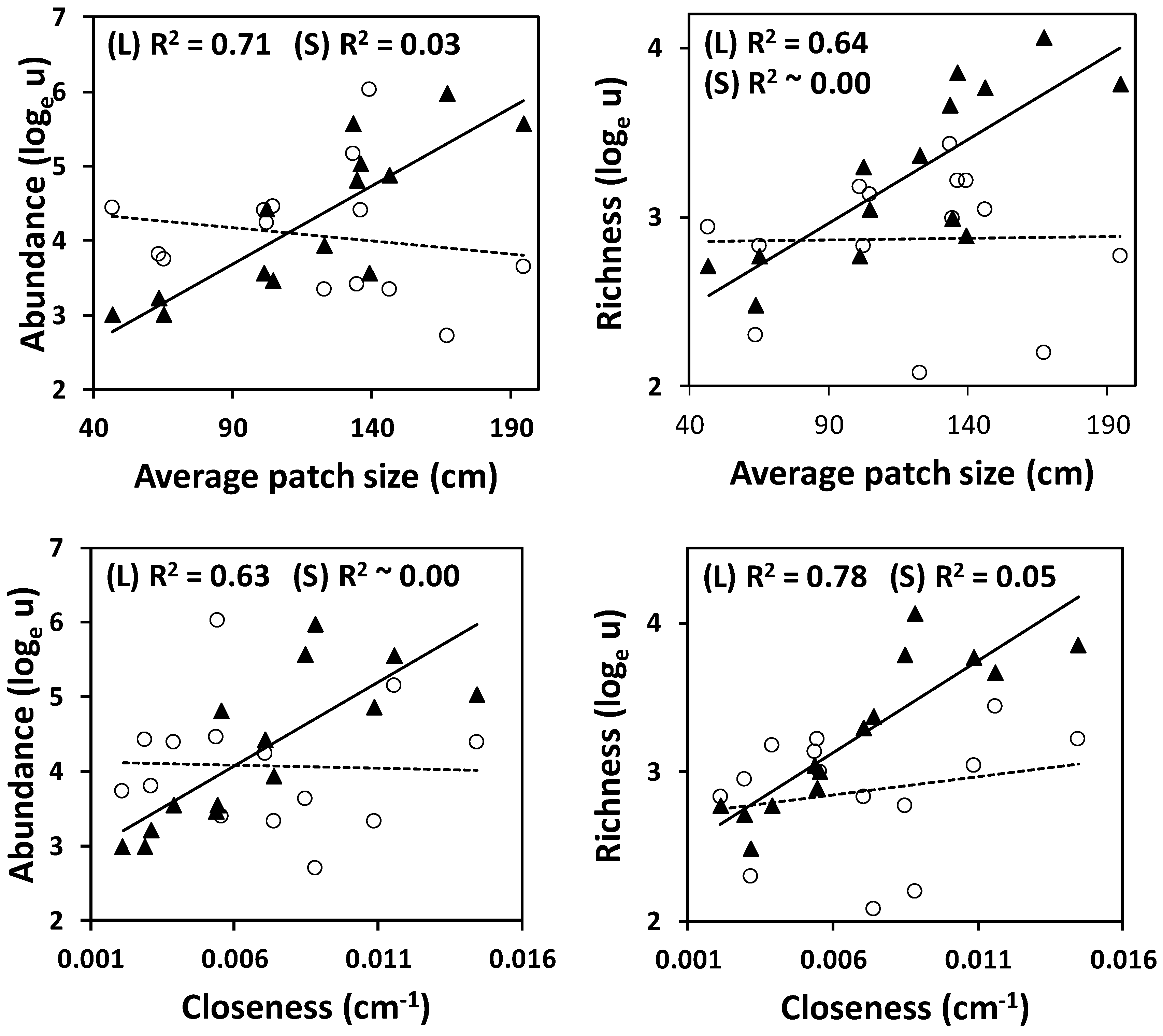
3.2. Sample-Based Analyses
4. Discussion
4.1. Ground Arthropods as a Function of Vegetation Pattern
4.2. Diversity of Meso-and Microarthropods in Mediterranean Drylands
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Millennium Ecosystem Assessment (MEA). Ecosystems and Human Well-Being: Desertification Synthesis; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Aguiar, M.R.; Sala, O.E. Patch structure, dynamics and implications for the functioning of arid ecosystems. Trends Ecol. Evol. 1999, 14, 273–277. [Google Scholar] [CrossRef]
- Tongway, D.J.; Hindley, N. Landscape Function Analysis: Procedures for Monitoring and Assessing Landscapes; CSIRO Publishing: Brisbane, Australia, 2004. [Google Scholar]
- Puigdefábregas, J. The role of vegetation patterns in structuring runoff and sediment fluxes in drylands. Earth Surf. Process. Landf. 2005, 30, 133–147. [Google Scholar] [CrossRef]
- Mayor, Á.G.; Bautista, S.; Bellot, J. Factors and interactions controlling infiltration, runoff, and soil loss at the microscale in a patchy Mediterranean semiarid landscape. Earth Surf. Process. Landf. 2009, 34, 1702–1711. [Google Scholar] [CrossRef]
- Maestre, F.T.; Eldridge, D.J.; Soliveres, S.; Kéfi, S.; Delgado-Baquerizo, M.; Bowker, M.A.; García-Palacios, P.; Gaitán, J.; Gallardo, A.; Lázaro, R.; et al. Structure and functioning of dryland ecosystems in a changing world. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, F.; Mayor, A.G.; Rietkerk, M.; Bautista, S. A null model for assessing the cover-independent role of bare soil connectivity as indicator of dryland functioning and dynamics. Ecol. Indic. 2018, 94, 512–519. [Google Scholar] [CrossRef]
- Titus, J.H.; Nowak, R.S.; Smith, S.D. Soil resources heterogeneity in the Mojave Desert. J. Arid Environ. 2002, 52, 269–292. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Zhu, F.; Song, N.; Yang, X.; Chai, Y. Seasonal distribution and diversity of ground arthropods in microhabitats following a shrub plantation age sequence in desertified steppe. PLoS ONE 2013, 8, e77962. [Google Scholar] [CrossRef] [Green Version]
- Kéfi, S.; Rietkerk, M.; Alados, C.L.; Pueyo, Y.; Papanastasis, V.P.; Elaich, A.; De Ruiter, P.C. Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature 2007, 449, 213–218. [Google Scholar] [CrossRef]
- Alados, C.L.; Puigdefábregas, J.; Martínez-Fernández, J. Ecological and socio-economical thresholds of land and plant-community degradation in semi-arid Mediterranean areas of southeastern Spain. J. Arid Environ. 2011, 75, 1368–1378. [Google Scholar] [CrossRef] [Green Version]
- Verwijmeren, M.; Rietkerk, M.; Bautista, S.; Mayor, A.G.; Wassen, M.J.; Smit, C. Drought and grazing combined: Contrasting shifts in plant interactions at species pair and community level. J. Arid Environ. 2014, 111, 53–60. [Google Scholar] [CrossRef]
- Rey, A.; Pegoraro, E.; Oyonarte, C.; Were, A.; Escribano, P.; Raimundo, J. Impact of land degradation on soil respiration in a steppe Stipa tenacissima L. semi-arid ecosystem in the SE of Spain. Soil Biol. Biochem. 2011, 432, 393–403. [Google Scholar] [CrossRef]
- Mayor, Á.G.; Kéfi, S.; Bautista, S.; Rodríguez, F.; Cartení, F.; Rietkerk, M. Feedbacks between vegetation pattern and resource loss dramatically decrease ecosystem resilience and restoration potential in a simple dryland model. Landsc. Ecol. 2013, 28, 931–942. [Google Scholar] [CrossRef]
- Zelnik, Y.R.; Kinast, S.; Yizhaq, H.; Bel, G.; Meron, E. Regime shifts in models of dryland vegetation. Philos. Trans. R. Soc. A 2013, 371, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hochstrasser, T.; Millington, J.D.A.; Papanastasis, V.P.; Parsons, A.J.; Roggero, P.P.; Brazier, R.E.; Estrany, J.; Farina, A.; Puttock, A. The Study of Land Degradation in Drylands: State of the Art. In Pattern of Land Degradation in Drylands; Mueller, E.N., Wainwright, J., Parsons, A.J., Turnbull, L., Eds.; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Mayor, A.G.; Bautista, S.; Rodriguez, F.; Kéfi, S. Connectivity-mediated ecohydrological feedbacks and regime shifts in drylands. Ecosystems 2019, 22, 1497–1511. [Google Scholar] [CrossRef] [Green Version]
- Lingbeek, B.J.; Higgins, C.L.; Muir, J.P.; Kattes, D.H.; Schwertner, T.W. Arthropod diversity and assemblage structure response to deforestation and desertification in the Sahel of western Senegal. Glob. Ecol. Conserv. 2017, 11, 165–176. [Google Scholar] [CrossRef]
- Meloni, F.; Varanda, E.M. Litter and soil arthropod colonization in reforested semi-deciduous seasonal Atlantic forests. Restor. Ecol. 2015, 23, 690–697. [Google Scholar] [CrossRef]
- Eisenbeis, G.; Wichard, W. Atlas on the Biology of Soil Arthropods; Springer: Berlin, Germany, 1987. [Google Scholar]
- Piñero, F.S.; Tinaut, A.; Aguirre-Segura, A.; Miñano, J.; Lencina, J.L.; Ortiz-Sánchez, F.J.; Pérez-López, F.J. Terrestrial arthropod fauna of arid areas of SE Spain: Diversity, biogeography, and conservation. J. Arid Environ. 2011, 75, 1321–1332. [Google Scholar] [CrossRef]
- Liu, J.L.; Li, F.R.; Liu, C.; Liu, Q.J.; Niu, R.X. Influences of shrub vegetation on distribution and diversity of a ground beetle community in a Gobi desert ecosystem. Biodivers. Conserv. 2012, 21, 2601–2619. [Google Scholar] [CrossRef]
- Whitford, W.G. The importance of biodiversity of soil biota in arid ecosystems. Biodivers. Conserv. 1996, 5, 185–195. [Google Scholar] [CrossRef]
- Wanner, M.; Dunger, W. Primary immigration and succession of soil organisms on reclaimed opencast coal mining areas in eastern Germany. Eur. J. Soil Biol. 2002, 38, 137–143. [Google Scholar] [CrossRef]
- Fitter, A.H.; Gilligan, C.A.; Hollingworth, K.; Kleczkowski, A.; Twyman, R.M.; Pitchford, J.W. Biodiversity and ecosystem function in soil. Funct. Ecol. 2005, 19, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Frouz, J.; Prack, K.; Pízl, V.; Hánel, L.; Starý, J.; Tajavský, K.; Materna, J.; Balík, V.; Kalcík, J.; Rehounková, K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Coleman, D.C.; Crossley, D.A.J.; Hendrix, P.F. Fundamentals of Soil Ecology, 3rd ed.; Academic Press: San Diego, CA, USA, 2018. [Google Scholar]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Siemann, E.; Haarstad, J.; Tilman, D. Dynamics of plant and arthropod diversity during old field succession. Ecography 1999, 22, 406–414. [Google Scholar] [CrossRef]
- Parisi, V.; Menta, C.; Gardi, C.; Jacomini, C.; Mozzanica, E. Microarthropod communities as a tool to assess soil quality and biodiversity: A new approach in Italy. Agric. Ecosyst. Environ. 2005, 105, 323–333. [Google Scholar] [CrossRef]
- Meloni, F. Influência do Desenvolvimento Florestal Sobre a Comunidade Edáfico-Epígea de Arthropoda e a Mirmecofauna: Bases Para a Bioindicação do Processo Sucessional na Restauração Ecológica. Ph.D. Thesis, FFCRP-USP, Ribeirão Preto, Brazil, 2012. [Google Scholar] [CrossRef] [Green Version]
- Ribas, C.R.; Campos, R.B.F.; Schmidt, F.A.; Solar, R.R.C. Ants as Indicators in Brazil: A Review with Suggestions to Improve the Use of Ants in Environmental Monitoring Programs. Psyche 2012, 2012, 636749. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, G.; Kumari, R.; Sharma, B.M. Mesofaunal biodiversity and its importance in Thar desert. J. Environ. Biol. 2007, 282, 503–515. [Google Scholar]
- Doblas-Miranda, E.; Sánchez-Piñero, F.; González-Megías, A. Different structuring factors but connected dynamics shape litter and belowground soil macrofaunal food webs. Soil Biol. Biochem. 2009, 41, 2543–2550. [Google Scholar] [CrossRef]
- Darby, B.J.; Neher, D.A.; Housman, D.C.; Belnap, J. Few apparent short-term effects of elevated soil temperature and increased frequency of summer precipitation on the abundance and taxonomic diversity of desert soil micro- and meso-fauna. Soil Biol. Biochem. 2011, 437, 1474–1481. [Google Scholar] [CrossRef]
- Walmsley, A.; Cerdà, A. Soil macrofauna and organic matter in irrigated orchards under Mediterranean climate. Biol. Agric. Hortic. 2017, 33, 247–257. [Google Scholar] [CrossRef]
- Liu, R.; Yosef, S.; Hou, J.; Zhao, J.; Liu, J.; Chang, H.; Zhang, J.; Luo, Y. Conversion of cropland into agroforestry land versus naturally-restored grassland alters soil macro-faunal diversity and trophic structure in the semi-arid agro-pasture zone of northern China. J. Arid Land 2019, 11, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Noble, J.C.; Whitford, W.G.; Kaliszweski, M. Soil and litter microarthropod populations from two contrasting ecosystems in semi-arid eastern Australia. J. Arid Environ. 1996, 32, 329–346. [Google Scholar] [CrossRef]
- Aquino, A.M. Fauna de solo e sua inserção na regulação funcional do agroecossistema. In Processos Biológicos no Sistema Solo-Planta; Aquino, A.M., Assis, R.L., Eds.; Embrapa: Brasília, Brazil, 2005. [Google Scholar]
- Maestre, F.T.; Escudero, A. Is the patch size distribution of vegetation a suitable indicator of desertification processes? Ecology 2009, 90, 1729–1735. [Google Scholar] [CrossRef] [PubMed]
- Berdugo, M.; Kéfi, S.; Soliveres, S.; Maestre, F.T. Plant spatial patterns identify alternative ecosystem multifunctionality states in global drylands. Nat. Ecol. Evol. 2017, 1, 0003. [Google Scholar] [CrossRef] [PubMed]
- Bascompte, J.; Rodrıíguez, M.A. Habitat patchiness and plant species richness. Ecol. Lett. 2001, 4, 417–420. [Google Scholar] [CrossRef]
- Mayor, A.G.; Bautista, S. Multi-scale evaluation of soil functional indicators for the assessment of soil and water retention in Mediterranean semiarid landscapes. Ecol. Indic. 2012, 20, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Urgeghe, A.M.; Bautista, S. Size and connectivity of upslope runoff-source areas modulate the performance of woody plants in Mediterranean drylands. Ecohydrology 2015, 8, 1292–1303. [Google Scholar] [CrossRef]
- Mayor, A.G.; Goirán, S.B.; Vallejo, V.R.; Bautista, S. Variation in soil enzyme activity as a function of vegetation amount, type, and spatial structure in fire-prone Mediterranean shrublands. Sci. Total Environ. 2016, 573, 1209–1216. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. World Reference Base for Soil Resources 2014: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resource Reports 106; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Oñate, J.J.; Peco, B. Policy impact on desertification: stakeholders’ perceptions in southeast Spain. Land Use Policy 2005, 22, 103–114. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to imageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Meloni, F.; Granzotti, C.R.F.; Bautista, S.; Martinez, A.S. Scale dependence and patch size distribution: Clarifying patch patterns in Mediterranean drylands. Ecosphere 2017, 8, e01690. [Google Scholar] [CrossRef]
- Meloni, F.; Nakamura, G.M.; Granzotti, C.R.F.; Martinez, A.S. Vegetation cover reveals the phase diagram of patch patterns in Mediterranean drylands. Physica A 2019, 534, 122048. [Google Scholar] [CrossRef]
- Groc, S.; Delabie, J.H.C.; Longino, J.T.; Orivel, J.; Majer, J.D.; Vasconcelos, H.L.; Dejean, A. A new method based on taxonomic sufficiency to simplify studies on Neotropical ant assemblages. Biol. Conserv. 2010, 143, 2832–2839. [Google Scholar] [CrossRef]
- Krell, F.T. Parataxonomy vs. taxonomy in biodiversity studies–pitfalls and applicability of “morphospecies” sorting. Biodivers. Conserv. 2004, 13, 795–812. [Google Scholar] [CrossRef]
- Madej, G.; Barczyk, G.; Gdawiec, M. Evaluation of soil biological quality index QBS-ar: Its sensitivity and usefulness in the post-mining chronosequence—Preliminary research. Pol. J. Environ. Stud. 2011, 205, 1367–1372. [Google Scholar]
- Ter Braak, C.J.F. Canonical Correspondence Analysis: A new eigenvector technique for multivariate direct gradiente analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B. Vegan: Community Ecology Package. R Package Version 2.5-6. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 1 September 2019).
- Swift, M.J.; Bignell, D.E.; Moreira, F.M.S.; Huising, E.J. The inventory of Soil Biological Diversity: Concepts and General Guidelines. In A Handbook of Tropical Soil Biology; Moreira, F.M.S., Huising, E.J., Bignell, D.E., Eds.; Earthscan: London, UK, 2008. [Google Scholar]
- Bardgett, R. The Biology of Soil: A Community and Ecosystem Approach; Oxford University Press: New York, NY, USA, 2005. [Google Scholar]
- Bochet, E.; Rubio, J.L.; Poesen, J. Modified topsoil islands within patchy Mediterranean vegetation in SE Spain. Catena 1999, 38, 23–44. [Google Scholar] [CrossRef]
- Lavelle, P.; Spain, A.V. Soil Ecology; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Horváth, R.; Magura, T.; Szinetár, C.; Eichardt, J.; Kovács, E.; Tóthmérész, B. In stable, unmanaged grasslands local factors are more important than landscape-level factors in shaping spider assemblages. Agric. Ecosyst. Environ. 2015, 208, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, J.; Ochoa, V.; Gozalo, B.; Quero, J.L.; García-Gómez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef] [Green Version]
- Horváth, R.; Magura, T.; Tóthmérész, B.; Eichardt, J.; Szinetár, C. Both local and landscape-level factors are important drivers in shaping ground-dwelling spider assemblages of sandy grasslands. Biodivers. Conserv. 2019, 28, 297–313. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests—Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]
- Magura, T.; Lövei, G.L.; Tóthmérész, B. Edge responses are different in edges under natural versus anthropogenic influence: A meta-analysis using ground beetles. Ecol. Evol. 2017, 7, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogoegraphy; Princenton University Press: Princenton, NJ, USA, 1967. [Google Scholar]
- Kitto, J.A.J.; Gray, D.P.; Hamish, S.G.; Niyogi, D.K.; Harding, J.S. Meta-community theory and stream restoration: Evidence that spatial position constrains stream invertebrate communities in a mine impacted landscape. Restor. Ecol. 2015, 233, 284–291. [Google Scholar] [CrossRef]
- Sola, F.J.; Peri, P.L.; Huertas, L.; MartÍnez Pastur, G.J.; Lencinas, M.V. Above-ground arthropod community structure and influence of structural-retention management in southern Patagonian scrublands, Argentina. J. Insect Conserv. 2016, 20, 929–944. [Google Scholar] [CrossRef]
- Liu, R.; Zhu, F.; Steinberger, Y. Changes in ground-dwelling arthropod diversity related to the proximity of shrub cover in a desertified system. J. Arid Environ. 2016, 124, 172–179. [Google Scholar] [CrossRef]
- Doblas-Miranda, E.; Sánchez-Piñero, F.; González-Megías, A. Vertical distribution of soil macrofauna in an arid ecosystem: Are litter and belowground compartmentalized habitats? Pedobiologia 2009, 526, 361–373. [Google Scholar] [CrossRef]
- Ellers, J.; Berg, M.P.; Dias, A.T.C.; Fontana, S.; Ooms, A.; Moretti, M. Diversity in form and function: Vertical distribution of soil fauna mediates multidimensional trait variation. J. Anim. Ecol. 2018, 87, 933–944. [Google Scholar] [CrossRef] [Green Version]
- Doblas-Miranda, E.; Sánchez-Piñero, F.; González-Megías, A. Soil macroinvertebrate fauna of a Mediterranean arid system: Composition and temporal changes in the assemblage. Soil Biol. Biochem. 2007, 398, 1916–1925. [Google Scholar] [CrossRef]
- Bautista, S.; Mayor, A.G.; Bourakhouadar, J.; Bellot, J. Plant spatial pattern predicts hillslope runoff and erosion in a semiarid mediterranean landscape. Ecosystems 2007, 10, 987–998. [Google Scholar] [CrossRef] [Green Version]
- Whitford, W.; Parker, L.W. Contributions of soil fauna to decomposition and mineralization processes in semiarid and arid ecosystems. Arid Soil Res. Rehabil. 1989, 3, 199–215. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Raaijmakers, C.E.; Zoomer, H.R.; Berg, M.P.; de Ruiter, P.C.; Verhoef, H.A.; van der Putten, W.H. Soil invertebrate fauna enhances grassland succession and diversity. Nature 2003, 422, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Ettema, C.H.; Wardle, D.A. Spatial soil ecology. Trends Ecol. Evol. 2002, 17, 117–183. [Google Scholar] [CrossRef]
- Menta, C.; Leoni, A.; Gardi, C.; Conti, F.D. Are grasslands important habitats for soil microarthropod conservation? Biodivers. Conserv. 2011, 20, 1073–1087. [Google Scholar] [CrossRef]
- Coleman, D.C.; Whitman, W.B. Linking species richness, biodiversity and ecosystem function in soil systems. Pedobiologia 2005, 49, 479–497. [Google Scholar] [CrossRef] [Green Version]
- Cepeda-Pizarro, J.G.; Whitford, W.G. Species abundance distribution patterns of soil microarthropods in decomposing surface leaf-litter and mineral soil on a desert watershed. Pedobiologia 1989, 33, 254–268. [Google Scholar]
- Villarreal-Rosas, J.; Palacios-Vargas, J.G.; Maya, Y. Microarthropod communities related with biological soil crusts in a desert scrub in northwestern Mexico. Rev. Mex. Biodivers. 2014, 85, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Prăvălie, R. Drylands extent and environmental issues. A global approach. Earth-Sci. Rev. 2016, 16, 259–278. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
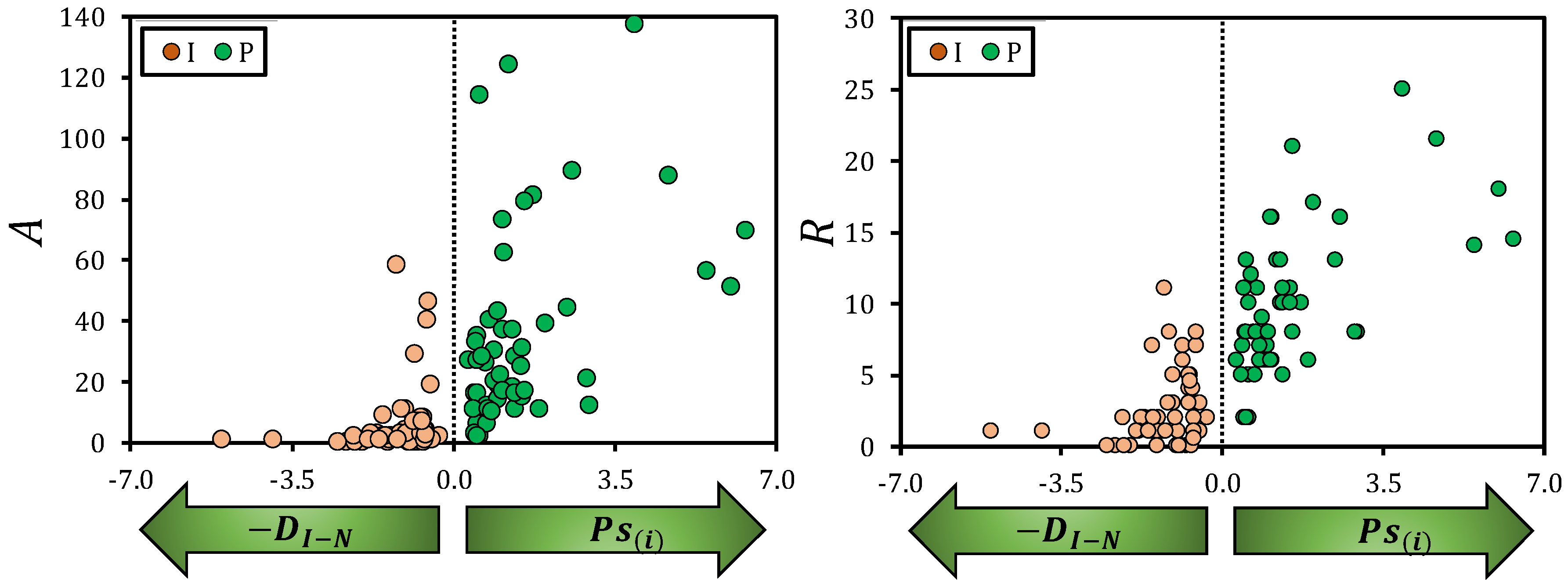{kind=link}
| ANCOVAs Microsite (M) | Abundance (log) | Richness (log) | ANCOVAs Soil Layer (L) | Abundance (log) | Richness (log) | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | ||
| VC | 28.5 | <0.001 *** | 30.5 | <0.001 *** | VC | 6.7 | 0.016 ** | 14.2 | <0.001 *** |
| M | 48.1 | <0.001 *** | 112.7 | <0.001 *** | L | 0.7 | 0.409 | 9.8 | 0.005 ** |
| VC:M‡ | 5.0 | 0.035 * | -- | -- | VC:L | 22.9 | <0.001 *** | 16.3 | <0.001 *** |
| F | p | F | p | F | p | F | p | ||
| PS | 16.2 | <0.001 *** | 14.3 | <0.001 *** | PS | 6.22 | 0.019 * | 7.9 | 0.010 ** |
| M | 34.4 | <0.001 *** | 79.8 | <0.001 *** | L | 0.6 | 0.461 | 7.0 | 0.014 * |
| PS:M‡ | -- | -- | -- | -- | PS:L | 12.4 | 0.002 ** | 7.3 | 0.012 * |
| F | p | F | p | F | p | F | p | ||
| Cl | 52.4 | <0.001 *** | 47.8 | <0.001 *** | Cl | 6.7 | 0.016 * | 15.7 | <0.001 *** |
| M | 74.9 | <0.001 *** | 156.1 | <0.001 *** | L | 0.5 | 0.483 | 8.4 | 0.008 ** |
| Cl:M | 13.2 | 0.001 ** | 5.0 | 0.035 * | Cl:L | 7.7 | 0.010 * | 7.0 | 0.014 * |
| Faunal Variable | Patches | Inter-Patches | |||||
|---|---|---|---|---|---|---|---|
| Vegetation Variable | Controlled By | p | p-Value | Controlled By | p | p-Value | |
| Abundance | Vc | Ps | 0.007 | 0.980 | Ps | 0.644 | 0.017 * |
| Cl | 0.313 | 0.297 | Cl | −0.065 | 0.831 | ||
| Ps; Cl | −0.301 | 0.341 | Ps; Cl | −0.154 | 0.633 | ||
| Ps | Vc | 0.482 | 0.095○ | Vc | −0.219 | 0.471 | |
| Cl | 0.578 | 0.038 * | Cl | 0.034 | 0.913 | ||
| Vc; Cl | 0.574 | 0.051○ | Vc; Cl | 0.143 | 0.657 | ||
| Cl | Vc | 0.085 | 0.782 | Vc | 0.728 | 0.005 ** | |
| Ps | 0.214 | 0.482 | Ps | 0.843 | <0.001 *** | ||
| Vc; Ps | 0.364 | 0.245 | Vc; Ps | 0.719 | 0.008 ** | ||
| Richness | Vc | Ps | 0.734 | 0.004 ** | Ps | 0.564 | 0.045 * |
| Cl | 0.616 | 0.025 * | Cl | −0.020 | 0.949 | ||
| Ps; Cl | 0.385 | 0.216 | Ps; Cl | −0.079 | 0.806 | ||
| Ps | Vc | −0.052 | 0.865 | Vc | −0.179 | 0.558 | |
| Cl | 0.525 | 0.065○ | Cl | 0.035 | 0.910 | ||
| Vc; Cl | 0.069 | 0.830 | Vc; Cl | 0.084 | 0.794 | ||
| Cl | Vc | 0.266 | 0.379 | Vc | 0.571 | 0.041 * | |
| Ps | 0.709 | 0.007 ** | Ps | 0.725 | 0.005 ** | ||
| Vs; Ps | 0.270 | 0.396 | Vc; Ps | 0.556 | 0.060○ | ||
| Abundance | Richness | ||||||
|---|---|---|---|---|---|---|---|
| Veg. Variable | Controlled By | p | p-Value | Veg. Variable | Controlled By | p | p-Value |
| PS(i) | NS | 0.48 | <0.001 *** | PS(i) | NS | 0.47 | <0.001 *** |
| DP-N | 0.34 | 0.011 * | DP-N | 0.46 | <0.001 *** | ||
| NS; DP-N | 0.58 | <0.003 ** | NS; DP-N | 0.60 | 0.001 ** | ||
| NS | PS(i) | −0.19 | 0.155 | NS | PS(i) | 0.01 | 0.919 |
| DP-N | −0.08 | 0.569 | DP-N | 0.17 | 0.207 | ||
| PS(i); DP-N | 0.05 | 0.721 | PS(i); DP-N | 0.252 | 0.063○ | ||
| DP-N | PS(i) | 0.18 | 0.190 | DP-N | PS(i) | −0.18 | 0.189 |
| NS | 0.37 | 0.005 ** | NS | −0.28 | 0.041 * | ||
| PS(i); NS | 0.20 | 0.134 | PS(i); NS | −0.14 | 0.306 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meloni, F.; F. Civieta, B.; A. Zaragoza, J.; Lourdes Moraza, M.; Bautista, S. Vegetation Pattern Modulates Ground Arthropod Diversity in Semi-Arid Mediterranean Steppes. Insects 2020, 11, 59. https://doi.org/10.3390/insects11010059
Meloni F, F. Civieta B, A. Zaragoza J, Lourdes Moraza M, Bautista S. Vegetation Pattern Modulates Ground Arthropod Diversity in Semi-Arid Mediterranean Steppes. Insects. 2020; 11(1):59. https://doi.org/10.3390/insects11010059
Chicago/Turabian StyleMeloni, Fernando, Berta F. Civieta, Juan A. Zaragoza, María Lourdes Moraza, and Susana Bautista. 2020. "Vegetation Pattern Modulates Ground Arthropod Diversity in Semi-Arid Mediterranean Steppes" Insects 11, no. 1: 59. https://doi.org/10.3390/insects11010059