The Parasitoid Eretmocerus hayati Is Compatible with Barrier Cropping to Decrease Whitefly (Bemisia tabaci MED) Densities on Cotton in China



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (1)
- Whitefly densities would remain lower under intercropping than perimeter planting;
- (2)
- The effect of parasitoids would be additive to the effect of barrier cropping;
- (3)
- These treatments would cause an increase in cotton yield.
2. Materials and Methods
2.1. Experimental Design and Plot Management
2.2. Parasitoid Rearing
2.3. Parasitoid Release
2.4. Sampling
2.4.1. B. tabaci Population
2.4.2. Parasitoid Census
2.4.3. Cotton Yield
2.5. Data Analysis
2.5.1. Barrier/Trap Crop Effect
2.5.2. Barrier/Trap Crop and Parasitoid Release Effect
2.5.3. Cotton Yield
3. Results
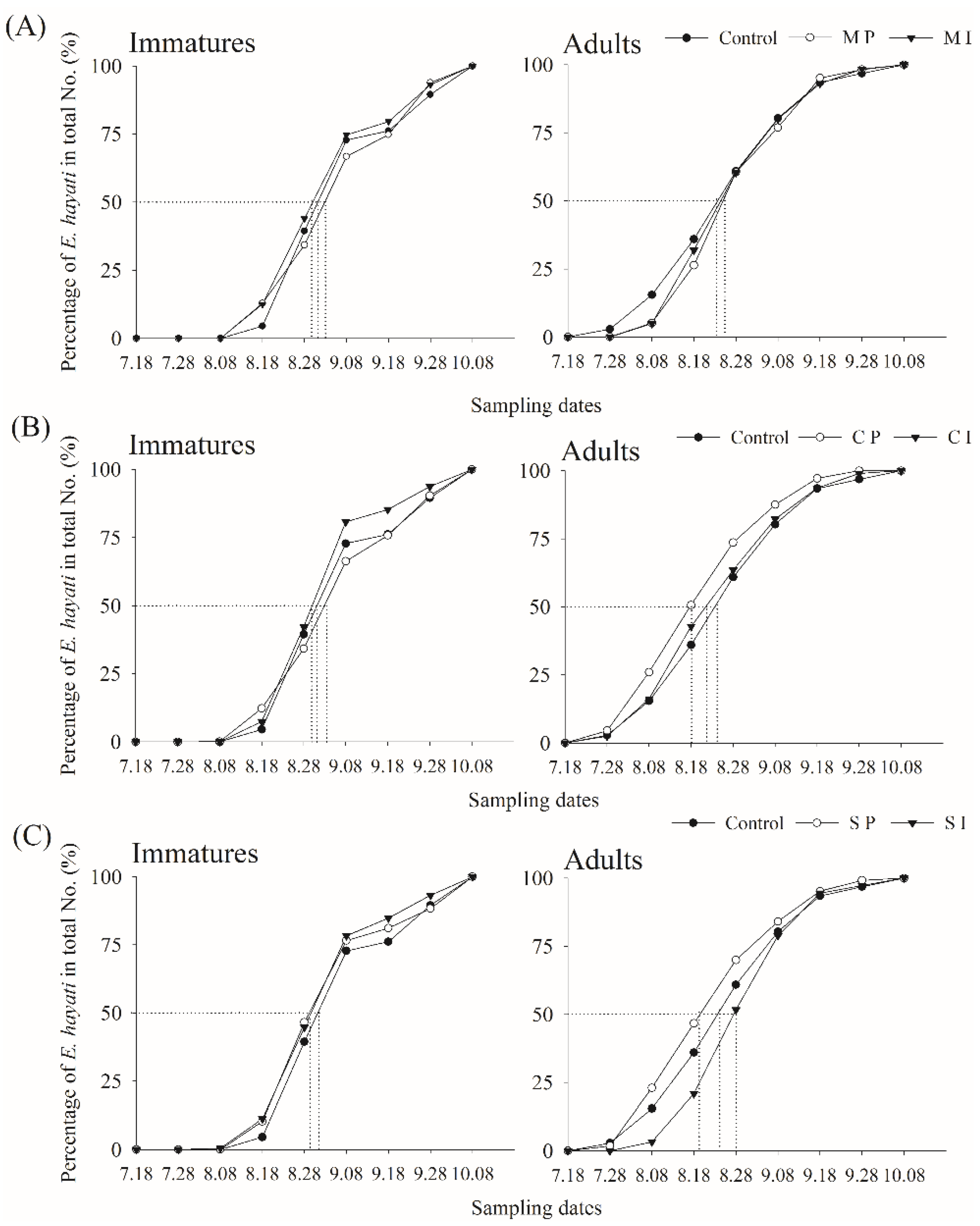
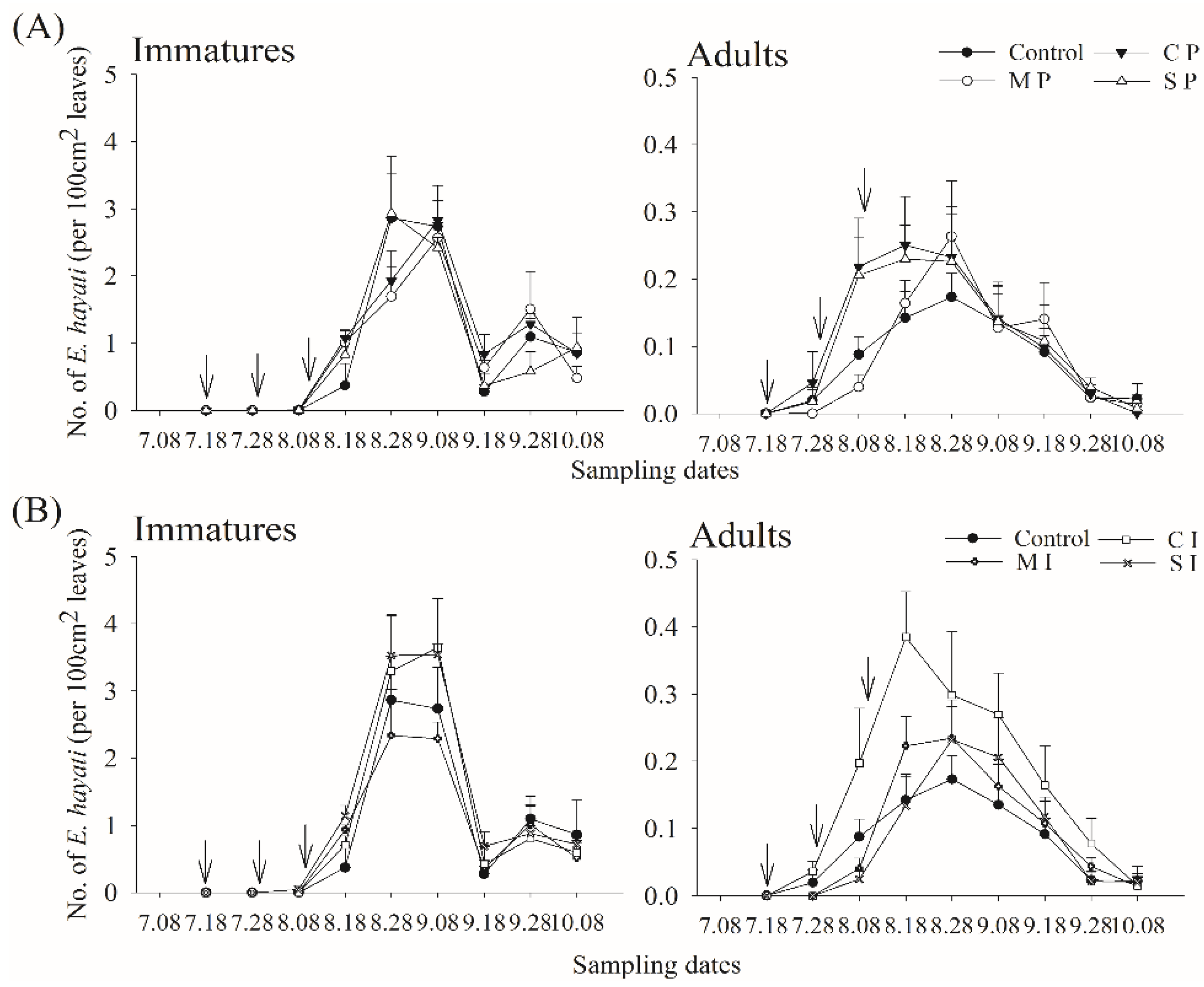
3.1. Adult and Immature Whitefly and Parasitoid Densities
3.2. Barrier/Trap Crop Effect
3.3. Barrier/Trap Crop and Parasitoid Release Effect
3.4. Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Whalon, M.E.; Mota-Sanchez, D.; Hollingworth, R.M. Global Pesticide Resistance in Arthropods; CABI: Wallingsford, UK, 2008; ISBN 978-1-84593-379-1. [Google Scholar]
- Kogan, M. Integrated pest management: Historical perspectives and contemporary developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, M.; Langrell, S.R.H.; Gomez-y-Paloma, S. Incentives and policies for integrated pest management in Europe: A review. Agron. Sustain. Dev. 2015, 35, 27–45. [Google Scholar] [CrossRef]
- Bostanian, N.J.; Goulet, H.; O’Hara, J.; Masner, L.; Racette, G. Towards insecticide free apple orchards: Flowering plants to attract beneficial arthropods. Biocontrol Sci. Technol. 2004, 14, 25–37. [Google Scholar] [CrossRef]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Lövei, G.L.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S.; et al. A functional overview of conservation biological control. Crop Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Andow, D. Effect of agricultural diversity on insect populations. In Proceedings of the 4th IFOAM International Scientific Conference, Malaysia, 29–31 August 1983; Praeger Science: New York, NY, USA, 1983; pp. 91–116. [Google Scholar]
- Andrews, D.J.; Kassam, A.H. The importance of multiple cropping in increasing world food supplies. In Multiple Cropping; ASA Special Publication: Wisconsin, WI, USA, 1976; pp. 1–10. [Google Scholar]
- Helenius, J. Intercropping, Insect Populations and Pest Damage: Case Study and Conceptual Model. Dep. Agric. For. Zool. Rep. 1989, 14, 1–49. [Google Scholar]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in Integrated Pest Management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, J.G. The Incidence and control of cauliflower mosaic in broccoli in South-West England. Ann. Appl. Biol. 1955, 43, 409–422. [Google Scholar] [CrossRef]
- Simons, J.N. Effects of insecticides and physical barriers on field spread of pepper veinbanding mosaic virus. Phytopathology 1957, 47, 139–145. [Google Scholar]
- Brown, J.K.; Frohlich, D.R.; Rosell, R.C. The sweetpotato or silverleaf whiteflies: Biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol. 1995, 40, 511–534. [Google Scholar] [CrossRef]
- Oliveira, M.R.V.; Henneberry, T.J.; Anderson, P. History, current status, and collaborative research projects for Bemisia tabaci. Crop Prot. 2001, 20, 709–723. [Google Scholar] [CrossRef] [Green Version]
- Boykin, L.M.; Shatters, R.G.; Rosell, R.C.; McKenzie, C.L.; Bagnall, R.A.; De Barro, P.; Frohlich, D.R. Global relationships of Bemisia tabaci (Hemiptera: Aleyrodidae) revealed using Bayesian analysis of mitochondrial COI DNA sequences. Mol. Phylogenet. Evol. 2007, 44, 1306–1319. [Google Scholar] [CrossRef] [PubMed]
- Yokomi, R.K.; Hoelmer, K.A.; Osborne, L.S. Relationships between the sweetpotato whitefly and the squash silverleaf disorder. Phytopathology 1990, 80, 895–900. [Google Scholar] [CrossRef]
- Zhang, X.M.; Yang, N.W.; Wan, F.H. Population density of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) on different plants in the field. Acta Ecol. Sin. 2014, 34, 4652–4661. (In Chinese) [Google Scholar] [CrossRef] [Green Version]
- Wan, F.H.; Yang, N.W. Invasion and management of agricultural alien insects in China. Annu. Rev. Entomol. 2016, 61, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.X.; Yang, Y.H.; Xie, B.Y.; Yang, C.R. Likely epidemics of virus diseases in vegetable crops caused by outbreaks of Bemisia tabaci. China Veg. 2001, 2, 34–35. (In Chinese) [Google Scholar] [CrossRef]
- Nauen, R.; Stumpf, N.; Elbert, A. Toxicological and mechanistic studies on neonicotinoid cross resistance in Q-type Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Manag. Sci. 2002, 58, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Rauch, N.; Nauen, R. Identification of biochemical markers linked to neonicotinoid cross resistance in Bemisia tabaci (Hemiptera: Aleyrodidae). Arch. Insect Biochem. 2003, 54, 165–176. [Google Scholar] [CrossRef]
- Morales, J.R.; Dardón, D.E.; Salguero, V.E. IPM Plot Validation and Transfer in Tomato. Manual of Management of Tomato Pests; Ministerio de Agricultura, Ganaderia y Alimentacion: Guatemala City, Guatemala, 1993; pp. 1991–1992. (In Spanish) [Google Scholar]
- Sharma, S.R.; Varma, A. Effect of cultural practices on virus infection in cowpea. Z. Acker Pflanzenbau 1984, 153, 23–31. [Google Scholar]
- Rataul, H.S.; Gill, C.K.; Brar, S. Use of barrier crop and some cultural measures in the management of yellow mosaic virus on soybean. J. Res. Punjab Agric. Univ. 1989, 26, 227–230. [Google Scholar]
- Gold, C.S.; Altieri, M.A.; Bellotti, A.C. Direct and residual effects of short duration intercrops on the cassava whiteflies Aleurotrachelus socialis and Trialeurodes variabilis (Homoptera: Aleyrodidae) in Colombia. Agric. Ecosyst. Environ. 1990, 32, 57–67. [Google Scholar] [CrossRef]
- Fargette, D.; Fauquet, C. A preliminary study on the influence of intercropping maize and cassava on the spread of African cassava mosaic virus by whiteflies. Asp. Appl. Biol. 1998, 17, 195–202. [Google Scholar]
- Zhang, X.M.; Lövei, G.L.; Ferrante, M.; Yang, N.W.; Wan, F.H. The potential of trap and barrier cropping to decrease densities of the whitefly Bemisia tabaci MED on cotton in China. Pest Manag. Sci. 2020, 76, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Gerling, D.; Horowitz, A.R.; Baumgaertner, J. Autecology of Bemisia tabaci. Agric. Ecosyst. Environ. 1986, 17, 5–19. [Google Scholar] [CrossRef]
- Breene, R.G.; Meagher, R.L., Jr.; Nordlund, D.A.; Wang, Y.-T. Biological control of Bemisia tabaci (Homoptera: Aleyrodidae) in a greenhouse using Chrysoperla rufilabris (Neuroptera: Chrysopidae). Biol. Control 1992, 2, 9–14. [Google Scholar] [CrossRef]
- Goolsby, J.A.; Ciomperlik, M.A.; Legaspi, B.C., Jr.; Legaspi, J.C.; Wendel, L.E. Laboratory and field evaluation of exotic parasitoids of Bemisia tabaci (Gennadius) (Biotype “B”) (Homoptera: Aleyrodidae) in the lower Rio Grande Valley of Texas. Biol. Control 1998, 12, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Zolnerowich, G.; Rose, M. Eretmocerus haldeman (Hymenoptera: Aphelinidae) imported and released in the United States for control of Bemisia tabaci complex (Homoptera: Aleyrodidae). Proc. Entomol. Soc. Wash. 1998, 100, 310–323. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Ripoll, R.; Gabarra, R.; Symondson, W.O.C.; King, R.A.; Agustí, N. Do the interactions among natural enemies compromise the biological control of the whitefly Bemisia tabaci? J. Pest Sci. 2014, 87, 133–141. [Google Scholar] [CrossRef]
- Gerling, D.; Alomar, Ò.; Arnò, J. Biological control of Bemisia tabaci using predators and parasitoids. Crop Prot. 2001, 20, 779–799. [Google Scholar] [CrossRef]
- Stansly, P.A.; Calvo, J.; Urbaneja, A. Release rates for control of Bemisia tabaci (Homoptera: Aleyrodidae) biotype “Q” with Eretmocerus mundus (Hymenoptera: Aphelinidae) in greenhouse tomato and pepper. Biol. Control 2005, 35, 124–133. [Google Scholar] [CrossRef]
- Yang, N.W.; Wan, F.H. Host suitability of different instars of Bemisia tabaci biotype B for the parasitoid Eretmocerus hayati. Biol. Control 2011, 59, 313–317. [Google Scholar] [CrossRef]
- De Barro, P.J.; Coombs, M.T. Post-release evaluation of Eretmocerus hayati Zolnerowich and Rose in Australia. Bull. Entomol. Res. 2009, 99, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Sivasubramaniam, V.; Subramaniam, S. Area-wide releases and evaluation of the parasitoid Eretmocerus hayati (Hymenoptera: Aphelinidae) for silverleaf whitefly control. In XXIX International Congress on Horticulture, International Symposia on Innovative Plant Protection in Horticulture, Biosecurity, Quarantine Pests, and Market Access; Hale, C., Hunter, D., Roberts, W., Ikin, R., McMaugh, S., Eds.; Acta Horticulturae: Brisbane, Australia, 2015; Volume 1105, pp. 81–88. [Google Scholar]
- Zhang, X.M.; Xu, H.Y.; Yang, N.W.; Wan, F.H. Field cage evaluation of the biocontrol effect of two aphelinid parasitoids on Bemisia tabaci Mediterranean (Hemiptera: Aleyrodidae). J. Plant Protect. 2018, 45, 1281–1288. (In Chinese) [Google Scholar] [CrossRef]
- Collier, T.R.; Hunter, M.S. Lethal interference competition in the whitefly parasitoids Eretmocerus eremicus and Encarsia sophia. Oecologia 2001, 129, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, S.E. Retrospective analysis of a classical biological control programme. J. Appl. Ecol. 2018, 55, 2439–2450. [Google Scholar] [CrossRef] [Green Version]
- Heinz, K.M.; Nelson, J.M. Interspecific interactions among natural enemies of Bemisia in an inundative biological control program. Biol. Control 1996, 6, 384–393. [Google Scholar] [CrossRef]
- Bográn, C.E.; Heinz, K.M.; Ciomperlik, M.A. Interspecific competition among insect parasitoids: Field experiments with whiteflies as hosts in cotton. Ecology 2002, 83, 653–668. [Google Scholar] [CrossRef]
- Zhang, X.M.; Yang, N.W.; Wan, F.H.; Lövei, G.L. Density and seasonal dynamics of Bemisia tabaci (Gennadius) Mediterranean on common crops and weeds around cotton fields in northern China. J. Integr. Agric. 2014, 13, 2211–2220. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.Y.; Yang, N.W.; Wan, F.H. Female reproductive system and ovary development of two parasitoids of tobacco whitefly. Sci. Technol. Rev. 2015, 33, 79–83. (In Chinese) [Google Scholar] [CrossRef]
- Jin, L.L.; Xu, M.; Wang, Z.S. Intercropping mode affecting yield and quality of cotton and peanut. Chin. Agric. Sci. Bull. 2018, 34, 51–55. (In Chinese) [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; Version 3.3.3; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Fazekas, J.; Kádár, F.; Sárospataki, M.; Lövei, G.L. Seasonal activity, age structure and egg production of the ground beetle Anisodactylus signatus (Coleoptera: Carabidae) in Hungary. Eur. J. Entomol. 1997, 94, 473–484. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Meth. Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Zuur, A.F.; Hilbe, J.M.; Ieno, E.N. A Beginner’s Guide to GLM and GLMM with R: A Frequentist and Bayesian Perspective for Ecologists; Highland Statistics Limited: Newburgh, UK, 2013.
- Gingerich, A.J.; Philipp, D.P.; Suski, C.D. Effects of nutritional status on metabolic rate, exercise and recovery in a freshwater fish. J. Comp. Physiol. B 2010, 180, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Selected Papers of Hirotugu Akaike; Parzen, E., Tanabe, K., Kitagawa, G., Eds.; Springer Series in Statistics; Springer: New York, NY, USA, 1998; pp. 199–213. [Google Scholar]
- Lavandero, B.; Wratten, S.; Shishehbor, P.; Worner, S. Enhancing the effectiveness of the parasitoid Diadegma semiclausum (Helen): Movement after use of nectar in the field. Biol. Control 2005, 34, 152–158. [Google Scholar] [CrossRef]
- Irvin, N.A.; Scarratt, S.L.; Wratten, S.D.; Frampton, C.M.; Chapman, R.B.; Tylianakis, J.M. The effects of floral understoreys on parasitism of leafrollers (Lepidoptera: Tortricidae) on apples in New Zealand. Agric. For. Entomol. 2006, 8, 25–34. [Google Scholar] [CrossRef]
- Gontijo, L.M.; Saldanha, A.V.; Souza, D.R.; Viana, R.S.; Bordin, B.C.; Antonio, A.C. Intercropping hampers the nocturnal biological control of aphids. Ann. Appl. Biol. 2018, 172, 148–159. [Google Scholar] [CrossRef]
- Razze, J.M.; Liburd, O.E.; Webb, S.E. Intercropping buckwheat with squash to reduce insect pests and disease incidence and increase yield. Agroecol. Sustain. Food Syst. 2016, 40, 863–891. [Google Scholar] [CrossRef]
- Hummel, J.D.; Dosdall, L.M.; Clayton, G.W.; Harker, K.N.; O’Donovan, J.T. Responses of the parasitoids of Delia radicum (Diptera: Anthomyiidae) to the vegetational diversity of intercrops. Biol. Control 2010, 55, 151–158. [Google Scholar] [CrossRef]
- Kristensen, N.P.; Barro, P.J.D.; Schellhorn, N.A. The initial dispersal and spread of an intentional invader at three spatial scales. PLoS ONE 2013, 8, e62407. [Google Scholar] [CrossRef] [Green Version]
- Castle, S.J. Concentration and management of Bemisia tabaci in cantaloupe as a trap crop for cotton. Crop Prot. 2006, 25, 574–584. [Google Scholar] [CrossRef]
- Mansion-Vaquié, A.; Ferrante, M.; Cook, S.M.; Pell, J.K.; Lövei, G.L. Manipulating field margins to increase predation intensity in fields of winter wheat (Triticum aestivum). J. Appl. Entomol. 2017, 141, 600–611. [Google Scholar] [CrossRef] [Green Version]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Hogg, B.N.; Bugg, R.L.; Daane, K.M. Attractiveness of common insectary and harvestable floral resources to beneficial insects. Biol. Control 2011, 56, 76–84. [Google Scholar] [CrossRef]
- Balzan, M.V.; Wäckers, F.L. Flowers to selectively enhance the fitness of a host–feeding parasitoid: Adult feeding by Tuta absoluta and its parasitoid Necremnus Artynes. Biol. Control 2013, 67, 21–31. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Ferrante, M.; Wan, F.; Yang, N.; Lövei, G.L. The Parasitoid Eretmocerus hayati Is Compatible with Barrier Cropping to Decrease Whitefly (Bemisia tabaci MED) Densities on Cotton in China. Insects 2020, 11, 57. https://doi.org/10.3390/insects11010057
Zhang X, Ferrante M, Wan F, Yang N, Lövei GL. The Parasitoid Eretmocerus hayati Is Compatible with Barrier Cropping to Decrease Whitefly (Bemisia tabaci MED) Densities on Cotton in China. Insects. 2020; 11(1):57. https://doi.org/10.3390/insects11010057
Chicago/Turabian StyleZhang, Xiaoming, Marco Ferrante, Fanghao Wan, Nianwan Yang, and Gábor L. Lövei. 2020. "The Parasitoid Eretmocerus hayati Is Compatible with Barrier Cropping to Decrease Whitefly (Bemisia tabaci MED) Densities on Cotton in China" Insects 11, no. 1: 57. https://doi.org/10.3390/insects11010057