Importance of Antixenosis and Antibiosis Resistance to the Cabbage Whitefly (Aleyrodes proletella) in Brussels Sprout Cultivars


Abstract
:1. Introduction
- Do cabbage whiteflies show preferences to certain Brussels sprout cultivars (i.e., antixenotic effects)?
- Are CWF fitness parameters affected when developing on different Brussels sprout cultivars (i.e., antibiotic effects)?
- Are there differences in the glucosinolate content among Brussels sprout cultivars, and how is this related to CWF preferences for cultivars?
2. Materials and Methods
2.1. Plant and Insect Culture
2.2. Screening Experiments (Antixenosis)
2.3. No-Choice Experiments (Antibiosis)
2.4. Field Trial
2.5. Glucosinolate Analysis
2.6. Statistics
3. Results
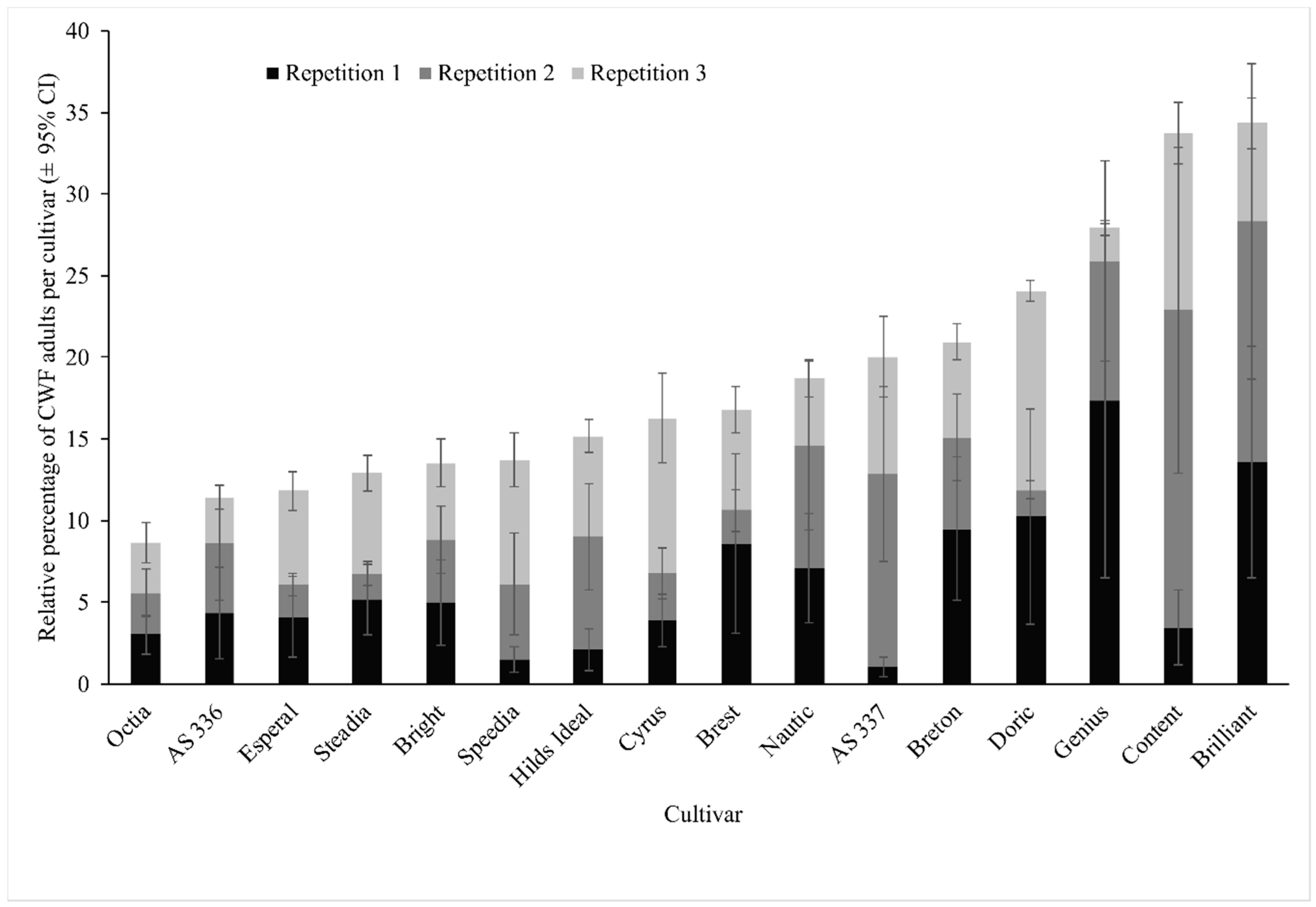
3.1. Screening Experiments (Antixenosis)
3.2. No-Choice Experiments (Antibiosis)
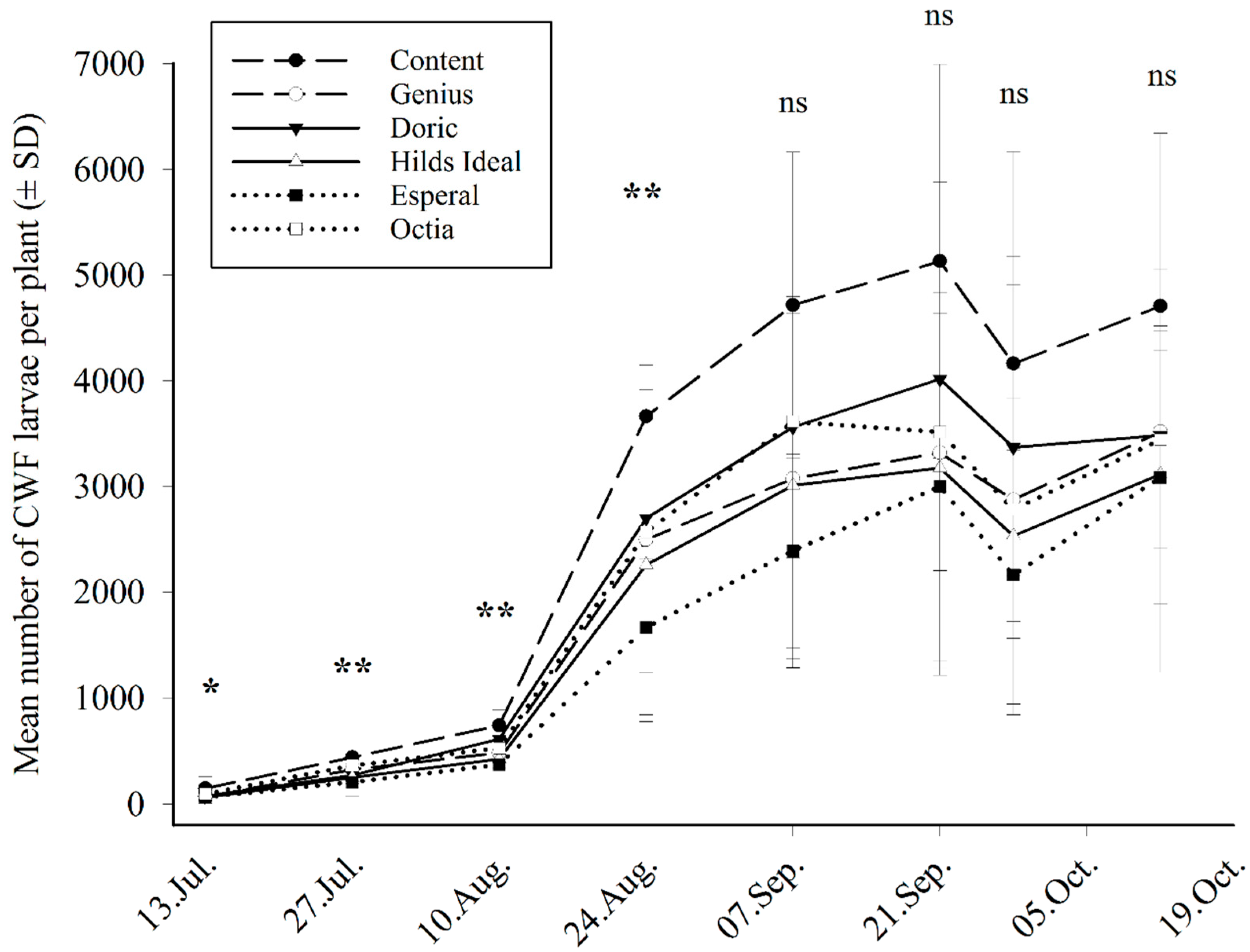
3.3. Field Trial
3.4. Glucosinolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Richter, E.; Hirthe, G. Effect of the cabbage whitefly Aleyrodes proletella on quality and yield of Brussels sprouts. In Julius-Kühn-Archiv, Proceedings of the 58 Deutsche Pflanzenschutztagung, Braunschweig, Germany, 10–14 September 2012; Julius Kühn-Institut: Braunschweig, Germany, 2012; Volume 438, pp. 351–352. [Google Scholar]
- Böckmann, E.; Richter, E. Influence of Aleyrodes proletella on yield and quality of Brussels sprouts. In Julius-Kühn-Archiv, Proceedings of the 60 Deutsche Pflanzenschutztagung, Halle, Germany, 20–23 September 2016; Julius Kühn-Institut: Braunschweig, Germany, 2016; Volume 454, p. 421. [Google Scholar]
- Hüweler, L.; Reineke, A.; Ludwig, M.; Hondelmann, P.; Meyhöfer, R. Relevance of winter rape for colonisation of cabbage by cabbage whitefly: Studying population genetics with microsatellite markers. In Julius-Kühn-Archiv, Proceedings of the 59 Deutsche Pflanzenschutztagung, Freiburg, Germany, 23–26 September 2014; Julius Kühn-Institut: Braunschweig, Germany, 2014; Volume 447, pp. 267–268. [Google Scholar]
- Ludwig, M.; Schlinkert, H.; Meyhöfer, R. Wind-modulated landscape effects on colonization of Brussels sprouts by insect pests and their syrphid antagonists. Agric. For. Entomol. 2017, 20, 1–9. [Google Scholar] [CrossRef]
- Ludwig, M.; Ludwig, H.; Conrad, C.; Dahms, T.; Meyhöfer, R. Cabbage whiteflies colonise Brassica vegetables primarily from distant, upwind source habitats. Entomol. Exp. Appl. 2019, 167, 713–721. [Google Scholar] [CrossRef] [Green Version]
- Askoul, K.; Richter, E.; Vidal, S.; Lusebrink, I. Life History Parameters of Aleyrodes proletella (Hemiptera: Aleyrodidae) on Different Host Plants. J. Econ. Entomol. 2018, 112, 457–464. [Google Scholar] [CrossRef]
- Richter, E.; Hirthe, G. Efficacy of drench and spray applications of insecticides to control cabbage whitefly Aleyrodes proletella. IOBC/WPRS Bull. 2014, 107, 151–156. [Google Scholar]
- Saucke, H.; Schultz, B.; Wedemeyer, R.; Liebig, N.; Zimmermann, O.; Katz, P. Biotechnische Regulierung der Kohlmottenschildlaus in Kohlgemüse–Sachstand und Perspektiven. Gesunde Pflanz. 2011, 63, 183–189. [Google Scholar] [CrossRef]
- Richter, E.; Hirthe, G. Hibernation and migration of Aleyrodes proletella in Germany. IOBC/WPRS Bull. 2014, 107, 143–149. [Google Scholar]
- Matyjaszczyk, E. “Biorationals” in integrated pest management strategies. J. Plant Dis. Prot. 2018, 125, 523–527. [Google Scholar] [CrossRef] [Green Version]
- Czaja, K.; Góralczyk, K.; Strucinski, P.; Hernik, A.; Korcz, W.; Minorczyk, M.; Łyczewska, M.; Ludwicki, J.K. Biopesticides–towards increased consumer safety in the European Union. Pest Manag. Sci. 2014, 71, 3–6. [Google Scholar] [CrossRef]
- Eigenbrode, S.; Trumble, J.T. Host Plant Resistance to Insects in Integrated Pest Management in Vegetable Crops. J. Agric. Entomol. 1994, 11, 201–224. [Google Scholar]
- Sharma, H.C.; Ortiz, R. Host plant resistance to insects: An eco-friendly approach for pest management and environmental conservation. J. Environ. Biol. 2002, 23, 111–135. [Google Scholar]
- Smith, C.M. Plant Resistance to Arthropods; Springer: Dordrecht, The Netherlands, 2005; pp. 19–100. [Google Scholar]
- Ramsey, A.; Ellis, P. Resistance in wild Brassicas to the cabbage whitefly, Aleyrodes proletella. Acta Hortic. 1996, 407, 507–514. [Google Scholar] [CrossRef]
- Vosman, B. A phenomics and genomics approach to the use of landraces and crop wild relatives for crop improvement. Crop Wild Relat. 2012, 8, 11–12. [Google Scholar]
- Pelgrom, K.T.B.; Broekgaarden, C.; Voorrips, R.E.; Bas, N.; Visser, R.G.F.; Vosman, B. Host plant resistance towards the cabbage whitefly in Brassica oleracea and its wild relatives. Euphytica 2015, 202, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.M.; Clement, S.L. Molecular bases of plant resistance to arthropods. Annu. Rev. Entomol. 2012, 57, 309–328. [Google Scholar] [CrossRef]
- Muñiz, M.; Nebreda, M. Differential variation in host preference of Aleyrodes proletella (L.) on several cauliflower cultivars. IOBC/WPRS Bull. 2003, 26, 49–52. [Google Scholar]
- Hirthe, G.; Jakobs, M. Sortenwahl hat Einfluss auf die Schädigung von Rosenkohl durch die Kohlmottenschildlaus. Info-Blatt für den Gartenbau Mecklenburg-Vorpommern 2013, 22, 320–329. [Google Scholar]
- Iheagwam, E.U. Influence of cabbage Brassica oleracea varieties and temperature on population variables of the cabbage whitefly Aleyrodes brassicae. Oikos 1981, 36, 233–237. [Google Scholar] [CrossRef]
- Nebreda, M.; Nombela, G.; Muñiz, M. Comparative Host Suitability of Some Brassica Cultivars for the Whitefly, Aleyrodes proletella (Homoptera: Aleyrodidae). Environ. Entomol. 2005, 34, 205–209. [Google Scholar] [CrossRef]
- Broekgaarden, C.; Riviere, P.; Steenhuis, G.; del sol Cuenca, M.; Kos, M.; Vosman, B. Phloem-specific resistance in Brassica oleracea against the whitefly Aleyrodes proletella. Entomol. Exp. Appl. 2012, 142, 153–164. [Google Scholar] [CrossRef]
- Iheagwam, E.U. The relationship betwen weight of insect, age, hardness and nitrogen content of cabbage leaves and fecundity of the cabbage whitefly, Aleyrodes brassicae Wlk. (Homoptera, Aleyrodidae). Z. Angew. Entomol. 1981, 91, 349–354. [Google Scholar] [CrossRef]
- Hopkins, R.; van Dam, N.M.; van Loon, J.J.A. Role of Glucosinolates in Insect-Plant Relationships and Multitrophic Interactions. Annu. Rev. Entomol. 2009, 54, 57–83. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, M.; Zrenner, R.; Krumbein, A.; Glatt, H.-R.; Schreiner, M. Genotypic variation of the glucosinolate profile in Pak Choi (Brassica rapa ssp. chinensis). J. Agric. Food Chem. 2013, 61, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Butler, C.G. On the Ecology of Aleurodes brassicae Walk. (Hemiptera). Trans. R. Entomol. Soc. Lond. 1938, 87, 291–311. [Google Scholar] [CrossRef]
- Iheagwam, E.U. Influence of host plant (Brassica species) and temperature on population increase of the cabbage whitefly Aleyrodes brassicae. Ann. Appl. Biol. 1980, 95, 273–278. [Google Scholar] [CrossRef]
- Poelman, E.H.; Van Loon, J.J.A.; Van Dam, N.M.; Vet, L.E.M.; Dicke, M. Herbivore-induced plant responses in Brassica oleracea prevail over effects of constitutive resistance and result in enhanced herbivore attack. Ecol. Entomol. 2010, 35, 240–247. [Google Scholar] [CrossRef]
- Staley, J.T.; Stewart-Jones, A.; Pope, T.W.; Wright, D.J.; Leather, S.R.; Hadley, P.; Rossiter, J.T.; Van Emden, H.F.; Poppy, G.M. Varying responses of insect herbivores to altered plant chemistry under organic and conventional treatments. Proc. R. Soc. B Biol. Sci. 2010, 277, 779–786. [Google Scholar] [CrossRef]
- Valladares, G.; Lawton, J.H. Host-Plant Selection in the Holly Leaf-Miner: Does Mother Know Best? J. Anim. Ecol. 1991, 60, 227–240. [Google Scholar] [CrossRef]
- Diaz-Montano, J.; Reese, J.C.; Schapaugh, W.T.; Campbell, L.R. Characterization of antibiosis and antixenosis to the soybean aphid (Hemiptera: Aphididae) in several soybean genotypes. J. Econ. Entomol. 2006, 99, 1884–1889. [Google Scholar] [CrossRef]
- Mayhew, P.J. Herbivore host choice and optimal bad motherhood. Trends Ecol. Evol. 2001, 16, 165–167. [Google Scholar] [CrossRef]
- Hufnagel, M.; Schilmiller, A.L.; Ali, J.; Szendrei, Z. Choosy mothers pick challenging plants: Maternal preference and larval performance of a specialist herbivore are not linked. Ecol. Entomol. 2017, 42, 33–41. [Google Scholar] [CrossRef]
- Lombaert, E.; Boll, R.; Lapchin, L. Dispersal strategies of phytophagous insects at a local scale: Adaptive potential of aphids in an agricultural environment. BMC Evol. Biol. 2006, 6, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekbom, B.; Rumei, X. Sampling and Spatial Patterns of Whiteflies. In Whiteflies: Their Bionomics, Pest Status and Management; Gerling, D., Ed.; Intercept Ltd.: Andover, UK, 1990; pp. 107–121. [Google Scholar]
- Rechner, O.; Poehling, H.-M. UV exposure induces resistance against herbivorous insects in broccoli. J. Plant Dis. Prot. 2014, 121, 125–132. [Google Scholar] [CrossRef]
- Su, Q.; Chen, G.; Mescher, M.C.; Peng, Z.; Xie, W.; Wang, S.; Wu, Q.; Liu, J.; Li, C.; Wang, W.; et al. Whitefly aggregation on tomato is mediated by feeding-induced changes in plant metabolites that influence the behaviour and performance of conspecifics. Funct. Ecol. 2018, 32, 1180–1193. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.J.; Chu, C. Insect Clip Cages Rapidly Alter Photosynthetic Traits of Leaves. Crop Sci. 1999, 39, 1896–1899. [Google Scholar] [CrossRef]
- White, J.A.; Whitham, T.G. Associational susceptibility of cottonwood to a box elder herbivore. Ecology 2000, 81, 1795–1803. [Google Scholar] [CrossRef]
- Santolamazza-Carbone, S.; Velasco, P.; Soengas, P.; Cartea, M.E. Bottom-up and top-down herbivore regulation mediated by glucosinolates in Brassica oleracea var. acephala. Oecologia 2014, 174, 893–907. [Google Scholar] [CrossRef] [Green Version]
- Elbaz, M.; Halon, E.; Malka, O.; Malitsky, S.; Blum, E.; Aharoni, A.; Morin, S. Asymmetric adaptation to indolic and aliphatic glucosinolates in the B and Q sibling species of Bemisia tabaci (Hemiptera: Aleyrodidae). Mol. Ecol. 2012, 21, 4533–4546. [Google Scholar] [CrossRef]
- Newton, E.L.; Bullock, J.M.; Hodgson, D. Temporal consistency in herbivore responses to glucosinolate polymorphism in populations of wild cabbage (Brassica oleracea). Oecologia 2010, 164, 689–699. [Google Scholar] [CrossRef]
- Moyes, C.L.; Collin, H.A.; Britton, G.; Raybould, A.F. Glucosinolates and differential herbivory in wild populations of Brassica oleracea. J. Chem. Ecol. 2000, 26, 2625–2641. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; Kliebenstein, D.J.; Chiem, A.; Morrill, E.; Mills, N.J.; Kremen, C. Chemically mediated tritrophic interactions: Opposing effects of glucosinolates on a specialist herbivore and its predators. J. Appl. Ecol. 2011, 48, 880–887. [Google Scholar] [CrossRef]
- Van Doorn, H.E. Development of Vegetables with Improved Consumer Quality: A Case Study in Brussels Sprouts. Ph.D. Thesis, Wageningen Universiteit, Wageningen, The Netherlands, 29 September 1999. [Google Scholar]
- Drewnowski, A.; Gomez-Carneros, C. Bitter taste, phytonutrients, and the consumer: A review. Am. J. Clin. Nutr. 2000, 72, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Vosman, B.; Pelgrom, K.T.B.; Sharma, G.; Broekgaarden, C.; Pritchard, J.K.; May, S.; Adobor, S.; Castellanos-Uribe, M.; van Kaauwen, M.; Finkers, R.; et al. Using Phenomics and Genomics to Unlock Landrace and Wild Relative Diversity for Crop Improvement. In Enhancing Crop Genepool Use: Capturing Wild Relative and Landrace Diversity for Crop Improvement; Maxted, N., Dulloo, M.E., Ford-Lloyd, B.V., Eds.; CABI: Boston, MA, USA, 2016; pp. 1–9. [Google Scholar]
- Chen, S.; Petersen, B.L.; Olsen, C.E.; Schulz, A.; Halkier, B.A. Long-distance phloem transport of glucosinolates in Arabidopsis. Plant Physiol. 2001, 127, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kissen, R.; Rossiter, J.T.; Bones, A.M. The “mustard oil bomb”: Not so easy to assemble? Localization, expression and distribution of the components of the myrosinase enzyme system. Phytochem. Rev. 2009, 8, 69–86. [Google Scholar] [CrossRef]
- Mewis, I.; Tokuhisa, J.G.; Schultz, J.C.; Appel, H.M.; Ulrichs, C.; Gershenzon, J. Gene expression and glucosinolate accumulation in Arabidopsis thaliana in response to generalist and specialist herbivores of different feeding guilds and the role of defense signaling pathways. Phytochemistry 2006, 67, 2450–2462. [Google Scholar] [CrossRef]
- Brader, G.; Mikkelsen, M.D.; Halkier, B.A.; Tapio Palva, E. Altering glucosinolate profiles modulates disease resistance in plants. Plant J. 2006, 46, 758–767. [Google Scholar] [CrossRef]
- Neugart, S.; Baldermann, S.; Hanschen, F.S.; Klopsch, R.; Wiesner-Reinhold, M.; Schreiner, M. The intrinsic quality of brassicaceous vegetables: How secondary plant metabolites are affected by genetic, environmental, and agronomic factors. Sci. Hortic. 2018, 233, 460–478. [Google Scholar] [CrossRef]
- Rosa, E. Daily variation in glucosinolate concentrations in the leaves and roots of cabbage seedlings in two constant temperature regimes. J. Sci. Food Agric. 1997, 73, 364–368. [Google Scholar] [CrossRef]
- UNECE. STANDARD FFV-08 Concerning the Marketing and Commercial Quality Control of Brussels Sprouts, 2017 Edition. Available online: http://www.unece.org/fileadmin/DAM/trade/agr/standard/standard/fresh/FFV-Std/English/08_BrusselsSprouts.pdf (accessed on 16 October 2019).
- Zehnder, G.; Gurr, G.M.; Kühne, S.; Wade, M.R.; Wratten, S.D.; Wyss, E. Arthropod pest management in organic crops. Annu. Rev. Entomol. 2007, 52, 57–80. [Google Scholar] [CrossRef] [Green Version]
- Lammerts van Bueren, E.T.; Jones, S.S.; Tamm, L.; Murphy, K.M.; Myers, J.R.; Leifert, C.; Messmer, M.M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS-Wageningen J. Life Sci. 2011, 58, 193–205. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Struik, P.C.; van Eekeren, N.; Nuijten, E. Towards resilience through systems-based plant breeding. A review. Agron. Sustain. Dev. 2018, 38, 1–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IFOAM. The IFOAM Norms for Organic Production and Processing. Available online: https://www.ifoam.bio/sites/default/files/ifoam_norms_july_2014_edits_2019.pdf (accessed on 26 November 2019).



{kind=link}
{kind=link}
| Cultivar | Total Developmental Time (Days) | Mortality (%) | Fecundity Rate | Life-Time Fertility | Female Weight (mg) | Longevity (Days) |
|---|---|---|---|---|---|---|
| Octia | 28.0 ± 3.35 ac | 27 ± 10 ac | 3.1 ± 1.3 a | 153.5 ± 88.4 a | 0.074 ± 0.018 ab | 63 ± 24 a |
| Esperal | 25.0 ± 2.98 b | 25 ± 8.0 ac | 3.5 ± 0.5 a | 134.8 ± 71.5 a | 0.085 ± 0.015 cd | 53 ± 30 a |
| Bright | 26.6 ± 2.49 ab | 41 ± 20 ab | 3.6 ± 1.7 a | 95.3 ± 52.4 a | 0.078 ± 0.019 abc | 48 ± 18 a |
| Speedia | 28.0 ± 2.95 ac | 39 ± 13 ab | 3.0 ± 0.8 a | 138.2 ± 118.5 a | 0.079 ± 0.013 abc | 64 ± 43 a |
| Hilds Ideal | 27.7 ± 2.70 ac | 19 ± 7.0 c | 3.3 ± 0.7 a | 116.6 ± 53.6 a | 0.091 ± 0.012 d | 49 ± 29 a |
| Doric | 27.6 ± 2.69 ac | 33 ± 15 abc | 2.5 ± 0.9 a | 91.4 ± 60.2 a | 0.071 ± 0.016 a | 53 ± 26 a |
| Genius | 26.6 ± 2.06 a | 44 ± 20 b | 3.1 ± 1.5 a | 93.8 ± 85.4 a | 0.080 ± 0.014 bc | 47 ± 24 a |
| Brilliant | 28.5 ± 4.08 c | 27 ± 14 ac | 2.8 ± 0.7 a | 117.3 ± 59.9 a | 0.075 ± 0.015 ab | 56 ± 24 a |
| Content | 26.7 ± 1.95 a | 37 ± 12 ab | 3.3 ± 0.9 a | 123.8 ± 62.6 a | 0.091 ± 0.012 d | 57 ± 23 a |
| Grand mean | 27.01 ± 2.82 | 32.0 ± 15.6 | 3.08 ± 1.06 | 115.8 ± 70.3 | 0.081 ± 0.015 | 53.8 ± 25.6 |
| Cultivar | Mortality (%) | Fecundity Rate | Degree of Contamination |
|---|---|---|---|
| Octia | 79 ± 27 ab | 2.8 ± 1.4 a | 3.3 ± 0.6 a |
| Esperal | 89 ± 20 b | 3.2 ± 1.5 a | 1.4 ± 0.5 c |
| Hilds Ideal | 88 ± 20 b | 2.9 ± 1.4 a | 3.3 ± 0.6 a |
| Doric | 80 ± 22 ab | 3.4 ± 1.6 a | 4.3 ± 0.5 b |
| Genius | 92 ± 12 ab | 1.6 ± 1.1 a | 2.3 ± 0.7 d |
| Content | 68 ± 29 a | 3.4 ± 1.6 a | 3.4 ± 0.9 a |
| Grand mean | 81.7 ± 24 | 3.03 ± 1.5 | 2.95 ± 1.1 |
| GS Concentration (µmol/g Dry Mass) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Upper Level Leaves | Mid–Level Leaves | |||||||
| Cultivar | Total GS | Indole GS | Alkenyl GS | Methylsulfinyl GS | Total GS | Indole GS | Alkenyl GS | Methylsulfinyl GS |
| Octia | 55.90 ± 32.40 a | 29.99 ± 18.21 a | 10.35 ± 7.94 b | 15.56 ± 8.10 ab | 11.60 ± 20.18 b | 7.06 ± 12.54 a | 1.17 ± 1.84 b | 3.37 ± 5.82 c |
| Hilds Ideal | 60.46 ± 11.19 a | 30.07 ± 6.94 a | 11.99 ± 5.44 b | 18.40 ± 5.71 a | 37.45 ± 14.93 a | 15.38 ± 8.47 a | 6.06 ± 4.78 ab | 16.01 ± 8.14 a |
| Doric | 65.11 ± 25.85 a | 35.11 ± 19.85 a | 18.81 ± 5.01 a | 11.19 ± 2.72 b | 22.68 ± 13.91 ab | 9.47 ± 6.08 a | 6.89 ± 4.86 a | 6.32 ± 4.66 bc |
| Genius | 62.88 ± 19.41 a | 35.97 ± 18.55 a | 11.67 ± 5.36 b | 15.24 ± 2.80 ab | 32.79 ± 23.46 a | 14.80 ± 12.40 a | 6.54 ± 5.38 a | 11.46 ± 6.46 ab |
| Content | 61.57 ± 19.65 a | 32.74 ± 14.22 a | 10.63 ± 4.08 b | 18.19 ± 3.20 a | 29.19 ± 16.10 ab | 12.32 ± 7.49 a | 6.12 ± 4.92 ab | 10.75 ± 4.63 ab |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hondelmann, P.; Paul, C.; Schreiner, M.; Meyhöfer, R. Importance of Antixenosis and Antibiosis Resistance to the Cabbage Whitefly (Aleyrodes proletella) in Brussels Sprout Cultivars. Insects 2020, 11, 56. https://doi.org/10.3390/insects11010056
Hondelmann P, Paul C, Schreiner M, Meyhöfer R. Importance of Antixenosis and Antibiosis Resistance to the Cabbage Whitefly (Aleyrodes proletella) in Brussels Sprout Cultivars. Insects. 2020; 11(1):56. https://doi.org/10.3390/insects11010056
Chicago/Turabian StyleHondelmann, Peter, Christina Paul, Monika Schreiner, and Rainer Meyhöfer. 2020. "Importance of Antixenosis and Antibiosis Resistance to the Cabbage Whitefly (Aleyrodes proletella) in Brussels Sprout Cultivars" Insects 11, no. 1: 56. https://doi.org/10.3390/insects11010056