Tracking Short-Range Attraction and Oviposition of European Grapevine Moths Affected by Volatile Organic Compounds in a Four-Chamber Olfactometer



Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Volatile Organic Compounds (VOCs)
2.3. Electroantennography (EAG)
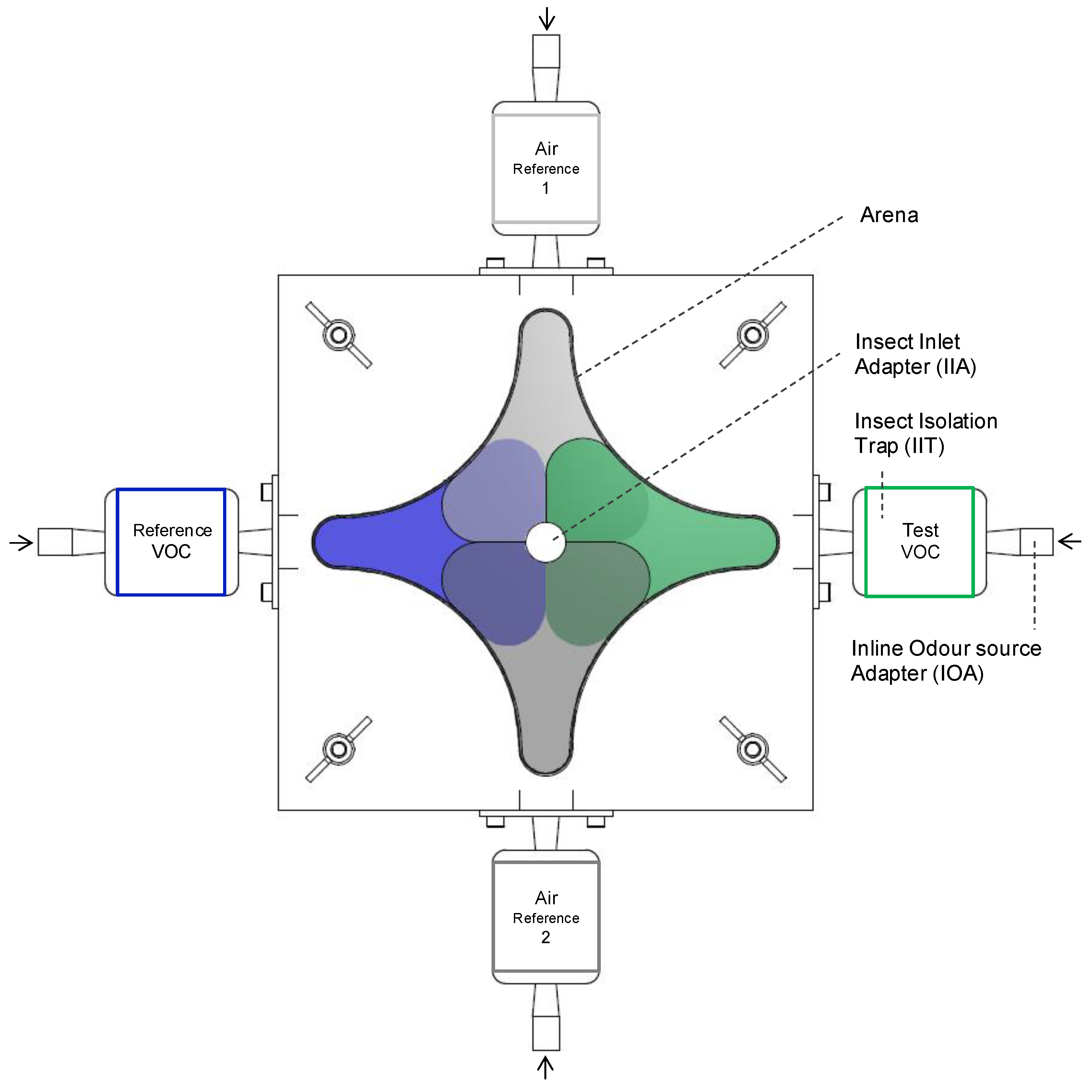
2.4. Four-Chamber Olfactometer Assays
2.5. Video Tracking System
2.6. Oviposition Bioassay
2.7. Statistical Analyses
3. Results
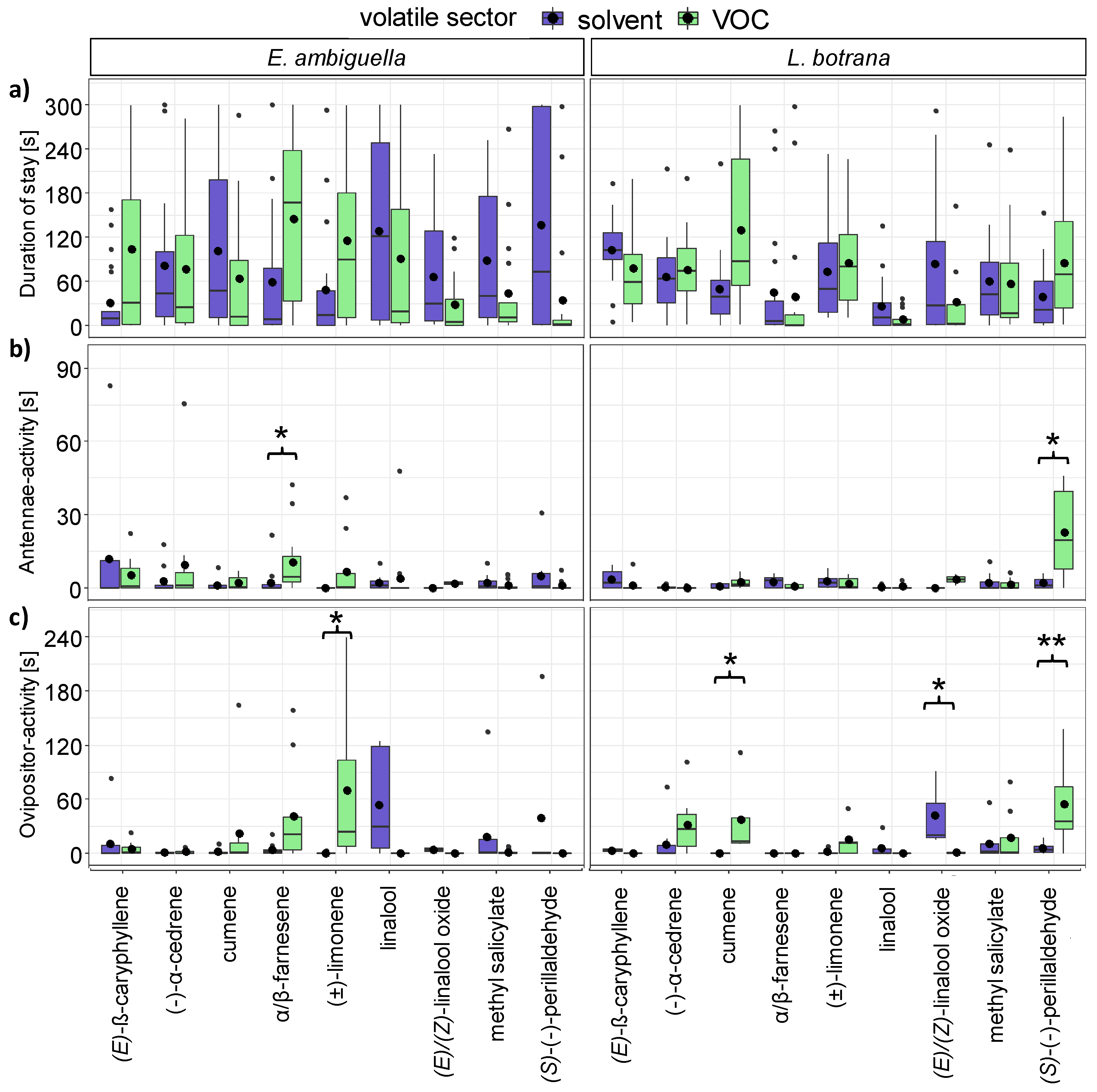
3.1. Perception of VOCs by EAG
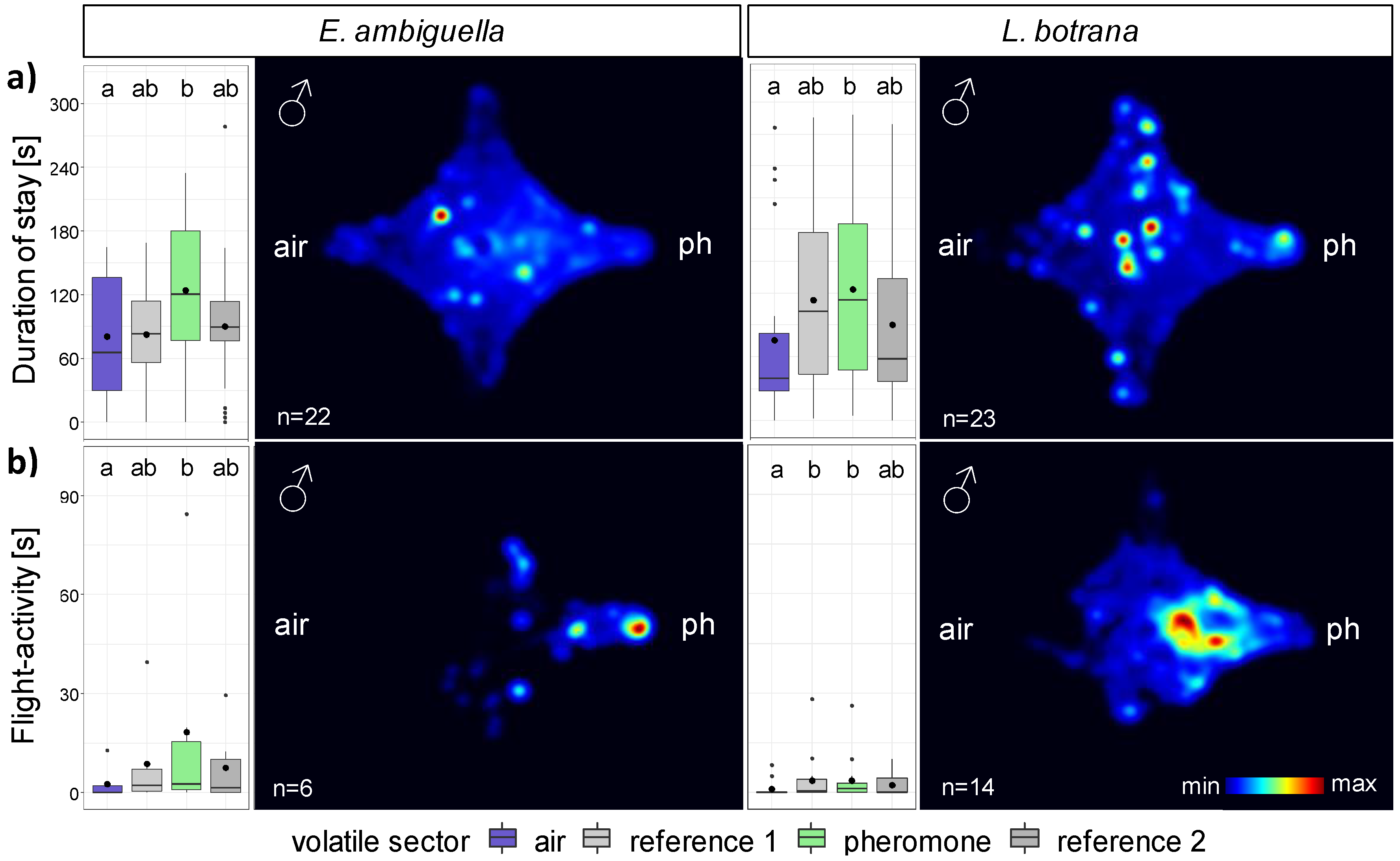
3.2. Responsiveness of Moths in the Olfactometer
3.2.1. Males
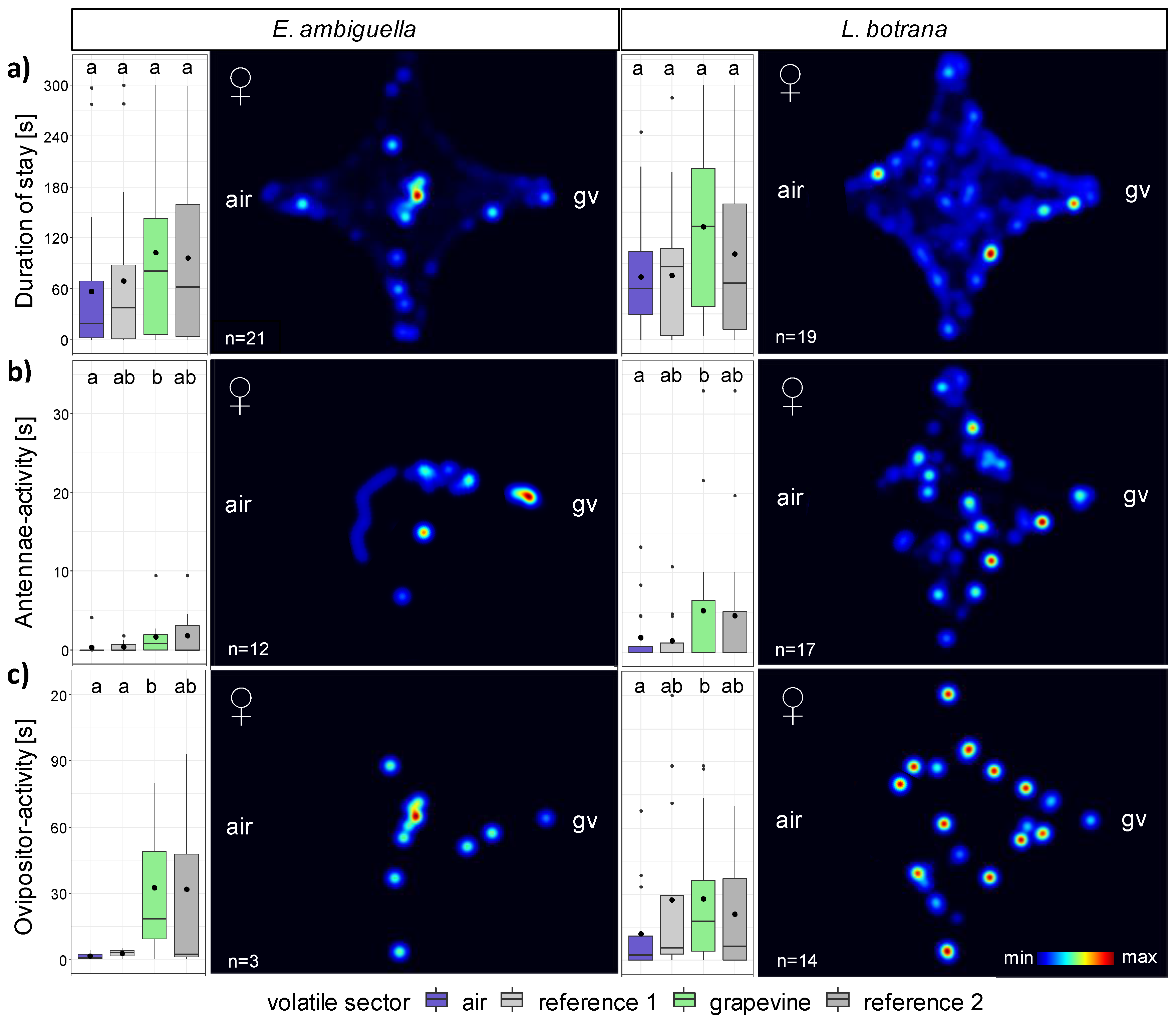
3.2.2. Females
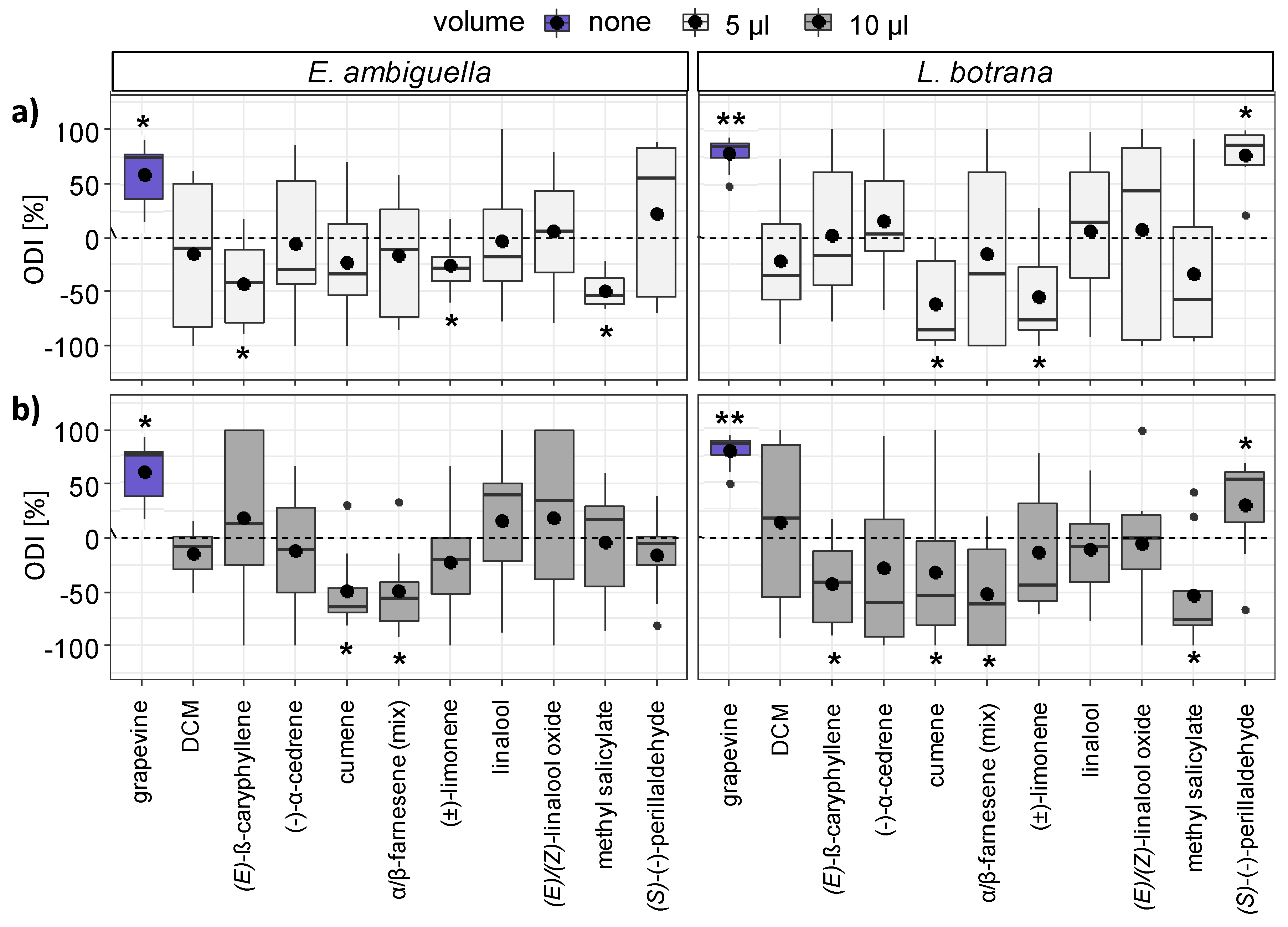
3.3. Short-Range Attraction of Females by VOCs
3.4. Oviposition Induced by VOCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Tasin et al., 2007 | Anfora et al., 2009 | Tasin et al., 2010 | Rid et al., 2019 | ||
|---|---|---|---|---|---|---|
| L * | B * | PS 33 * | GM * | DM * | Grapevine * | |
| Monoterpenes | ||||||
| (E)/(Z)-linalool oxide (furanoid) | ◦ | ● | ◦ | ● | ● | ● |
| (E)/(Z)-linalool oxide (pyranoid) | ◦ | ◦ | ◦ | ◦ | ● | ◦ |
| (±)-limonene | ◦ | ◦ | ● | ◦ | ◦ | ● |
| linalool | ◦ | ● | ● | ● | ● | ● |
| Sesquiterpenes | ||||||
| (E)-β-caryophyllene | ● | ● | ● | ● | ● | ● |
| (E)-β-farnesene | ● | ◦ | ◦ | ● | ◦ | ● |
| (E,E)-α-farnesene | ◦ | ● | ● | ● | ● | ● |
| (−)-α-cedrene | ◦ | ◦ | ◦ | ◦ | ◦ | ● |
| Benzenoids | ||||||
| methyl salicylate | ◦ | ● | ● | ● | ● | ● |
| cumene | ◦ | ◦ | ◦ | ◦ | ◦ | ● |
References
- Knolhoff, L.M.; Heckel, D.G. Behavioral Assays for Studies of Host Plant Choice and Adaptation in Herbivorous Insects. Annu. Rev. Entomol. 2014, 59, 263–278. [Google Scholar] [CrossRef]
- Pavan, F.; Stefanelli, G.; Villani, A.; Cargnus, E. Influence of Grapevine Cultivar on the Second Generations of Lobesia botrana and Eupoecilia ambiguella. Insects 2018, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilligan, T.M.; Epstein, M.E.; Passoa, S.C.; Powell, J.A.; Sage, O.C.; Brown, J.W. Discovery of Lobesia Botrana ([Denis & Schiffermuller]) in California: An Invasive Species New to North America (Lepidoptera: Tortricidae). Proc. Entomol. Soc. Wash. 2011, 113, 14–30. [Google Scholar] [CrossRef]
- Varela, L.G.; Smith, R.J.; Cooper, M.; Hoenisch, R.W. European grapevine moth, Lobesia botrana, in Napa Valley vineyards. Pract. Winery Vineyard 2010, 2010, 1–5. [Google Scholar]
- Gonzalez, M. Lobesia botrana: Polilla da la uva. Rev. Enol. 2010, 2, 2–5. [Google Scholar]
- Lucchi, A.; Sambado, P.; Royo, A.B.J.; Bagnoli, B.; Benelli, G. Lobesia botrana males mainly fly at dusk: Video camera-assisted pheromone traps and implications for mating disruption. J. Pest Sci. 2018, 91, 1327–1334. [Google Scholar] [CrossRef]
- Stellwaag, F. Neue Foschungen über Mottenflug, Eiablage und Eidauer der Traubenwickler. Der Dtsch. Weinbau 1943, 22, 203–206. [Google Scholar]
- Balachowsky, A.; Mensil, L. Les insectes nuisibles aux plantes cultivées. Paris 1935, 1, 137. [Google Scholar]
- Savopoulou-Soultani, Μ.; Stavridis, D.G.; Tzanakakis, Μ.Ε. Development and reproduction of Lobesia botrana on vine and olive inflorescences. Entomol. Hell. 2017, 8, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Thiery, D.; Moreau, J. Relative performance of European grapevine moth (Lobesia botrana) on grapes and other hosts. Oecologia 2005, 143, 548–557. [Google Scholar] [CrossRef]
- Roditakis, N.E. Effectiveness of Bacillus thuringiensis Berliner var. Kurstaki on the Grape Berry Moth Lobesia botrana Den. and Shiff. (Lepidoptera, Tortricidae) under Field and Laboratory Conditions in Crete. Entomol. Hell. 1986, 4, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Stellwaag, F. Die Weinbauinsekten Der Kulturländer; Paul Parey: Berlin, Germany, 1928. [Google Scholar]
- Broumas, T.; Stavraki, H.; Souliotis, C.; Sarafis, N. Observations on the Biology of Grape Moth Lobesia Botrana De. et. Schiff in Kavala, Greece; A.A. In Proceedings of the Meeting of the EC Expert’s Group, Thessaloniki, Greece, 6–8 October 1987; Balkema: Roterdam, The Netherlands, 1989; pp. 57–62. [Google Scholar]
- Moschos, T. Yield loss quantification and assessment of economic injury level for the anthophagous generation of the European grapevine moth Lobesia botrana Den. et Schiff. (Lepidoptera: Tortricidae). Int. J. Pest Manag. 2005, 51, 81–89. [Google Scholar] [CrossRef]
- Delbac, L.; Thiery, D. Damage to grape flowers and berries by Lobesia botrana larvae (Denis & Schiffernuller) (Lepidoptera: Tortricidae), and relation to larval age. Aust. J. Grape Wine Res. 2016, 22, 256–261. [Google Scholar] [CrossRef]
- Fermaud, M.; Lemenn, R. Association of Botrytis cinerea with Grape Berry Moth Larvae. Phytopathology 1989, 79, 651–656. [Google Scholar] [CrossRef]
- Roehrich, R.; Carles, J.P.; Tresor, C.; Devathaire, M.A. Experiments of Pheromonal Control on the Grape Vine Moths Lobesia botrana and Eupoecilia ambiguella. Ann. Zool. Ecol. Anim. 1979, 11, 659–675. [Google Scholar]
- Ioriatti, C.; Guzzon, R.; Anfora, G.; Ghidoni, F.; Mazzoni, V.; Villegas, T.R.; Dalton, D.T.; Walton, V.M. Drosophila suzukii (Diptera: Drosophilidae) Contributes to the Development of Sour Rot in Grape. J. Econ. Entomol. 2018, 111, 283–292. [Google Scholar] [CrossRef]
- Barata, A.; Santos, S.C.; Malfeito-Ferreira, M.; Loureiro, V. New Insights into the Ecological Interaction Between Grape Berry Microorganisms and Drosophila Flies During the Development of Sour Rot. Microb. Ecol. 2012, 64, 416–430. [Google Scholar] [CrossRef]
- Schmidt-Busser, D.; von Arx, M.; Connetable, S.; Guerin, P.M. Identification of host-plant chemical stimuli for the European grape berry moth Eupoecilia ambiguella. Physiol. Entomol. 2011, 36, 101–110. [Google Scholar] [CrossRef]
- Tasin, M.; Anfora, G.; Ioriatti, C.; Carlin, S.; De Cristofaro, A.; Schmidt, S.; Bengtsson, M.; Versini, G.; Witzgall, P. Antennal and behavioral responses of grapevine moth Lobesia botrana females to volatiles from grapevine. J. Chem. Ecol. 2005, 31, 77–87. [Google Scholar] [CrossRef]
- von Arx, M.; Schmidt-Busser, D.; Guerin, P.M. Host plant volatiles induce oriented flight behaviour in male European grapevine moths, Lobesia botrana. J. Insect Physiol. 2011, 57, 1323–1331. [Google Scholar] [CrossRef]
- Vacas, S.; Alfaro, C.; Zarzo, M.; Navarro-Llopis, V.; Primo, J. Effect of sex pheromone emission on the attraction of Lobesia botrana. Entomol. Exp. Appl. 2011, 139, 250–257. [Google Scholar] [CrossRef]
- Tasin, M.; Backman, A.C.; Bengtsson, M.; Varela, N.; Ioriatti, C.; Witzgall, P. Wind tunnel attraction of grapevine moth females, Lobesia botrana, to natural and artificial grape odour. Chemoecology 2006, 16, 87–92. [Google Scholar] [CrossRef]
- Tasin, M.; Lucchi, A.; Ioriatti, C.; Mraihi, M.; De Cristofaro, A.; Boger, Z.; Anfora, G. Oviposition Response of the Moth Lobesia botrana to Sensory Cues from a Host Plant. Chem. Senses 2011, 36, 633–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rid, M.; Markheiser, A.; Hoffmann, C.; Gross, J. Waxy bloom on grape berry surface is one important factor for oviposition of European grapevine moths. J. Pest Sci. 2018, 91, 1225–1239. [Google Scholar] [CrossRef]
- Maher, N.; Thiery, D. Distribution of chemo- and mechanoreceptors on the tarsi and ovipositor of female European grapevine moth, Lobesia botrana. Entomol. Exp. Appl. 2004, 110, 135–143. [Google Scholar] [CrossRef]
- Tasin, M.; Backman, A.C.; Coracini, M.; Casado, D.; Ioriatti, C.; Witzgall, P. Synergism and redundancy in a plant volatile blend attracting grapevine moth females. Phytochemistry 2007, 68, 203–209. [Google Scholar] [CrossRef]
- Tasin, M.; Backman, A.C.; Anfora, G.; Carlin, S.; Ioriatti, C.; Witzgall, P. Attraction of Female Grapevine Moth to Common and Specific Olfactory Cues from 2 Host Plants. Chem. Senses 2010, 35, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Anfora, G.; Tasin, M.; De Cristofaro, A.; Ioriatti, C.; Lucchi, A. Synthetic Grape Volatiles Attract Mated Lobesia botrana Females in Laboratory and Field Bioassays. J. Chem. Ecol. 2009, 35, 1054–1062. [Google Scholar] [CrossRef]
- Thiery, D.; Gabel, B. Inter-Specific Avoidance of Egg-Associated Semiochemicals in 4 Tortricids. Experientia 1993, 49, 998–1001. [Google Scholar] [CrossRef]
- Markheiser, A.; Rid, M.; Biancu, S.; Gross, J.; Hoffmann, C. Physical factors influencing the oviposition behaviour of European grapevine moths Lobesia botrana and Eupoecilia ambiguella. J. Appl. Entomol. 2018, 142, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Cattaneo, A.M.; Bengtsson, J.M.; Borgonovo, G.; Bassoli, A.; Anfora, G. Response of the European grapevine moth Lobesia botrana to somatosensory-active volatiles emitted by the non-host plant Perilla frutescens. Physiol. Entomol. 2014, 39, 229–236. [Google Scholar] [CrossRef]
- Rid, M.; Markheiser, A.; Stein, S.; Hoffmann, C.; Gross, J. Volatiles of several grapevine cultivars emitted at different phenological stages linked to discriminatory ability of grapevine moths. J. Plant Dis. Protect. 2019, 126, 115–127. [Google Scholar] [CrossRef]
- Cattaneo, A.M.C. Response of the tortricid pest Lobesia botrana to volatiles emitted by the non-host plant Perilla frutescens. Chem. Senses 2014, 39, 91. [Google Scholar]
- Rid, M.; Mesca, C.; Ayasse, M.; Gross, J. Apple Proliferation Phytoplasma Influences the Pattern of Plant Volatiles Emitted Depending on Pathogen Virulence. Front. Ecol. Evol. 2016, 3, 152. [Google Scholar] [CrossRef] [Green Version]
- RCoreTeam. R: A Language and Environment for Staistical Computing. Available online: https://www.R-project.org (accessed on 15 September 2019).
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, AKA Least-Squares Means. R Package Version 1.3.4. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 15 September 2019).
- Maher, N.; Thiery, D. A bioassay to evaluate the activity of chemical stimuli from grape berries on the oviposition of Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 2004, 94, 27–33. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis. Available online: https://ggplot2.tidyverse.org (accessed on 15 September 2019).
- Götz, B. Neue Apparate zum Studium der Insektenphysiologie. Sonderdr. Die Umschau 1941, 49, 779–781. [Google Scholar]
- Prokopy, R.J.; Owens, E.D. Visual Detection of Plants by Herbivorous Insects. Annu. Rev. Entomol. 1983, 28, 337–364. [Google Scholar] [CrossRef]
- Gabel, B.; Thiery, D. Oviposition response of Lobesia botrana females to long-chain free fatty acids and esters from its eggs. J. Chem. Ecol. 1996, 22, 161–171. [Google Scholar] [CrossRef]
- Gabel, B.; Thiery, D.; Suchy, V.; Marionpoll, F.; Hradsky, P.; Farkas, P. Floral Volatiles of Tanacetum vulgare L Attractive to Lobesia botrana Den-Et-Schiff Females. J. Chem. Ecol. 1992, 18, 693–701. [Google Scholar] [CrossRef]
- Schoonhoven, L.M. Insect-plant relationships: The whole is more than the sum of its parts. Entomol. Exp. Appl. 2005, 115, 5–6. [Google Scholar] [CrossRef]
- Likhayo, P.W.; Hodges, R.J. Field monitoring Sitophilus zeamais and Sitophilus oryzae (Coleoptera: Curculionidae) using refuge and flight traps baited with synthetic pheromone and cracked wheat. J. Stored Prod. Res. 2000, 36, 341–353. [Google Scholar] [CrossRef]
- Nielsen, M.C.; Sansom, C.E.; Larsen, L.; Worner, S.P.; Rostas, M.; Chapman, R.B.; Butler, R.C.; de Kogel, W.J.; Davidson, M.M.; Perry, N.B.; et al. Volatile compounds as insect lures: Factors affecting release from passive dispenser systems. N. Zeal. J. Crop Hort. Sci. 2019, 47, 208–223. [Google Scholar] [CrossRef]
- Moreau, J.; Rahme, J.; Benrey, B.; Thiery, D. Larval host plant origin modifies the adult oviposition preference of the female European grapevine moth Lobesia botrana. Naturwissenschaften 2008, 95, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Birgucu, A.K.; Turanli, F.; Gumus, E.; Guzel, B.; Karsavuran, Y. The effect of grape cultivars on oviposition preference and larval survival of Lobesia botrana Den. & Schiff. (Lepidoptera: Tortricidae). Fresen. Environ. Bull. 2015, 24, 33–38. [Google Scholar]
- Hurtrel, B.; Thiery, D. Modulation of flight activity in Lobesia botrana Den. & Schiff. (Lepidoptera: Tortricidae) females studied in a wind tunnel. J. Insect Behav. 1999, 12, 199–211. [Google Scholar] [CrossRef]
- von Arx, M.; Schmidt-Busser, D.; Guerin, P.M. Plant Volatiles Enhance Behavioral Responses of Grapevine Moth Males, Lobesia botrana to Sex Pheromone. J. Chem. Ecol. 2012, 38, 222–225. [Google Scholar] [CrossRef]
- Galet, P. Les Maladies Et Les Parasites De La Vigne; Galet, P., Ed.; Impr. du Paysan du Midi: Montpellier, France, 1982. [Google Scholar]
- Silva, D.; Curkovic, T.; Ceballos, R. Behavioral and antennal responses of Lobesia botrana (Lepidoptera: Tortricidae) to volatiles from the non-host plant Schinus molle L. (Anacardiaceae). Chil. J. Agric. Res. 2019, 79, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Thiery, D.; Gabel, B.; Farkas, P.; Pronier, V. Identification of an Oviposition-Regulating Pheromone in the European Grapevine Moth, Lobesia botrana (Lepidoptera, Tortricidae). Experientia 1992, 48, 697–699. [Google Scholar] [CrossRef]
- Hatanaka, A.; Kajiwara, T.; Kenji, M. The biogeneration of green odor by green leaves and it’s physiological functions—Present, past and future. Z. Für Nat. C 1995, 50, 467–472. [Google Scholar] [CrossRef]
- Lin, Y.W.; Qasim, M.; Hussain, M.; Akutse, K.S.; Avery, P.B.; Dash, C.K.; Wang, L.D. The Herbivore-Induced Plant Volatiles Methyl Salicylate and Menthol Positively affect Growth and Pathogenicity of Entomopathogenic Fungi. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.M.; Lima, M.C.H.P.; Araujo, H.C.; Esteves, R.L.; Kaplan, M.A.C. Evaluation of the volatile components and the seasonal variation of the methyl salicylate from Stifftia chrysantha Mikan by HS-SPME/GC-MS. Bol. Lat. Caribe Plantas Med. Aromat. 2012, 11, 413–419. [Google Scholar]
- James, D.G. Field evaluation of herbivore-induced plant volatiles as attractants for beneficial insects: Methyl salicylate and the green lacewing, Chrysopa nigricornis. J. Chem. Ecol. 2003, 29, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Snoeren, T.A.L.; Mumm, R.; Poelman, E.H.; Yang, Y.; Pichersky, E.; Dicke, M. The Herbivore-Induced Plant Volatile Methyl Salicylate Negatively Affects Attraction of the Parasitoid Diadegma semiclausum. J. Chem. Ecol. 2010, 36, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.W.; Park, K.C. Methyl salicylate, a soybean aphid-induced plant volatile attractive to the predator Coccinella septempunctata. J. Chem. Ecol. 2005, 31, 1733–1746. [Google Scholar] [CrossRef]
- Tasin, M.; Betta, E.; Carlin, S.; Gasperi, F.; Mattivi, F.; Pertot, I. Volatiles that encode host-plant quality in the grapevine moth. Phytochemistry 2011, 72, 1999–2005. [Google Scholar] [CrossRef]
- Ulland, S.; Ian, E.; Mozuraitis, R.; Borg-Karlson, A.K.; Meadow, R.; Mustaparta, H. Methyl salicylate, identified as primary odorant of a specific receptor neuron type, inhibits oviposition by the moth Mamestra brassicae L. (Lepidoptera, noctuidae). Chem. Senses 2008, 33, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, O.R.; Hutchins, R.F. Attraction of Newly Hatched Codling Moth Larvae (Laspeyresia pomonella) to Synthetic Stereo-Isomers of Farnesene. J. Insect Physiol. 1973, 19, 723–727. [Google Scholar] [CrossRef]
- Sutherland, O.R.; Hutchins, R.F. Alpha-Farnesene, a Natural Attractant for Codling Moth Larvae. Nature 1972, 239, 170. [Google Scholar] [CrossRef]
- Hern, A.; Dorn, S. Sexual dimorphism in the olfactory orientation of adult Cydia pomonella in response to alpha-farnesene. Entomol. Exp. Appl. 1999, 92, 63–72. [Google Scholar] [CrossRef]
- Wearing, C.H.; Hutchins, R.F. Alpha-Farnesene, a Naturally Occurring Oviposition Stimulant for Codling Moth, Laspeyresia pomonella. J. Insect Physiol. 1973, 19, 1251–1256. [Google Scholar] [CrossRef]
- Yan, F.; Bengtsson, M.; Makranczy, G.; Lofqvist, J. Roles of alpha-farnesene in the behaviors of codling moth females. Z. Naturforsch. C 2003, 58, 113–118. [Google Scholar] [CrossRef] [PubMed]





| Compound | E. ambiguella | L. botrana | ||||||
|---|---|---|---|---|---|---|---|---|
| n | Response [mV] ± SD | p-Value 1 | n | Response [mV] ± SD | p-Value 1 | |||
| (E)-β-caryophyllene | 3 × 5 | −1.14 ± 0.22 | 9.30 × 10−12 | *** | 3 × 5 | −0.63 ± 0.23 | 3.30 × 10−9 | *** |
| (−)-α-cedrene | 3 × 5 | −0.63 ± 0.22 | 2.35 × 10−7 | *** | 3 × 5 | −0.41 ± 0.18 | 1.13 × 10−3 | ** |
| cumene | 3 × 5 | −0.67 ± 0.35 | 1.39 × 10−8 | *** | 3 × 5 | −0.37 ± 0.10 | 5.85 × 10−8 | *** |
| α/β-farnesene | 3 × 5 | −1.45 ± 0.61 | 9.94 × 10−7 | *** | 3 × 5 | −1.12 ± 0.40 | 7.58 × 10−10 | *** |
| (+)-limonene | 3 × 5 | −0.82 ± 0.39 | 1.79 × 10−7 | *** | 3 × 5 | −0.58 ± 0.24 | 1.69 × 10−8 | *** |
| (−)-limonene | 3 × 5 | −0.96 ± 0.35 | 1.25 × 10−7 | *** | 3 × 5 | −0.82 ± 0.26 | 6.47 × 10−8 | *** |
| linalool | 3 × 5 | −1.88 ± 0.46 | 8.82 × 10−14 | *** | 3 × 5 | −1.29 ± 0.41 | 1.35 × 10−10 | *** |
| (E)/(Z)-linalool oxide (furanoid) | 3 × 5 | −1.40 ± 0.51 | 2.77 × 10−11 | *** | 3 × 5 | −0.69 ± 0.22 | 1.89 × 10−7 | *** |
| (E)/(Z)-linalool oxide (pyranoid) | 3 × 5 | −0.98 ± 0.67 | 1.68 × 10−5 | *** | 3 × 5 | −0.80 ± 0.26 | 1.62 × 10−10 | *** |
| methyl salicylate | 3 × 5 | −1.10 ± 0.43 | 1.07 × 10−8 | *** | 3 × 5 | −1.06 ±0.46 | 2.39 × 10−7 | *** |
| DCM | 3 × 50 | −0.44 ± 0.26 | 3 × 50 | −0.36 ± 0.18 | ||||
| air | 3 × 50 | 0.48 ± 0.34 | 3 × 50 | 0.38 ± 0.22 | ||||
| Species | Behavior | n | FPM in Sector (Mean ± SE) [n] | DPM in Sector (Mean ± SE) [s] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Air | Reference 1 (Air) | Pher | Reference 2 (Air) | Air | Reference 1 (Air) | Pher | Reference 2 (Air) | |||
| EA | stay | 22 | 41.4 ± 6.3 | 55.6 ± 9.9 | 47.3 ± 6.9 | 58.2 ± 10.4 | 80.5 ± 12.2 | 82.8 ± 9.5 | 124.2 ± 14.9 | 90.8 ± 13.0 |
| flight | 6 | 5.2 ± 3.6 | 9.6 ± 6.7 | 7.8 ± 4.9 | 10.7 ± 7.6 | 2.6 ± 2.1 | 8.8 ± 6.3 | 18.3 ± 13.6 | 7.5 ± 4.8 | |
| LB | stay | 23 | 30.7 ± 7.0 | 29.0 ± 4.2 | 31.4 ± 4.7 | 37.6 ± 7.4 | 75.9 ± 16.6 | 113.2 ± 17.0 | 123.8 ± 16.9 | 90.2 ± 16.2 |
| flight | 14 | 0.5 ± 0.4 | 1.6 ± 0.7 | 1.4 ± 0.5 | 2.0 ± 1.1 | 1.0 ± 0.7 | 3.7 ± 2.0 | 3.7 ± 1.9 | 2.3 ± 0.9 | |
| Species | Behavior | n | FPM in Sector (Mean ± SE) [n] | DPM in Sector (Mean ± SE) [s] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Air | Reference 1 (Air) | Grape | Reference 2 (Air) | Air | Reference 1 (Air) | Grape | Reference 2 (Air) | |||
| EA | stay | 21 | 32.8 ± 9.8 | 25.3 ± 9.1 | 35.5 ± 10.5 | 25.8 ± 7.4 | 42.7 ± 19.1 | 45.5 ± 19.0 | 82.3 ± 25.4 | 72.6 ± 22.7 |
| antennae | 12 | 0.3 ± 0.3 | 0.6 ± 0.3 | 1.6 ± 0.5 | 1.0 ± 0.4 | 0.3 ± 0.3 | 0.4 ± 0.2 | 1.7 ± 0.8 | 1.8 ± 0.9 | |
| ovipositor | 3 | 0.7 ± 0.3 | 1.0 ± 0.6 | 4.7 ± 2.9 | 1.3 ± 0.9 | 1.6 ± 1.2 | 2.7 ± 1.5 | 32.7 ± 24.1 | 31.8 ± 30.6 | |
| LB | stay | 19 | 11.9 ± 2.5 | 11.2 ± 2.4 | 28.0 ± 12.1 | 11.5 ± 3.0 | 73.8 ± 15.4 | 75.4 ± 17.6 | 132.6 ± 22.1 | 100.5 ± 21.8 |
| antennae | 17 | 0.7 ± 0.3 | 0.6 ± 0.2 | 1.4 ± 0.5 | 1.1 ± 0.5 | 1.9 ± 0.9 | 1.5 ± 0.7 | 5.3 ± 2.2 | 4.6 ± 2.2 | |
| ovipositor | 14 | 2.4 ± 0.7 | 3.1 ± 0.8 | 3.2 ± 0.8 | 2.2 ± 0.6 | 12.1 ± 5.4 | 27.5 ± 10.2 | 27.9 ± 8.6 | 21.0 ± 7.5 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markheiser, A.; Rid, M.; Biancu, S.; Gross, J.; Hoffmann, C. Tracking Short-Range Attraction and Oviposition of European Grapevine Moths Affected by Volatile Organic Compounds in a Four-Chamber Olfactometer. Insects 2020, 11, 45. https://doi.org/10.3390/insects11010045
Markheiser A, Rid M, Biancu S, Gross J, Hoffmann C. Tracking Short-Range Attraction and Oviposition of European Grapevine Moths Affected by Volatile Organic Compounds in a Four-Chamber Olfactometer. Insects. 2020; 11(1):45. https://doi.org/10.3390/insects11010045
Chicago/Turabian StyleMarkheiser, Anna, Margit Rid, Sandra Biancu, Jürgen Gross, and Christoph Hoffmann. 2020. "Tracking Short-Range Attraction and Oviposition of European Grapevine Moths Affected by Volatile Organic Compounds in a Four-Chamber Olfactometer" Insects 11, no. 1: 45. https://doi.org/10.3390/insects11010045