1. Introduction
More than 60% of the termite species of the family Termitidae rely on soil-feeding [
1]; among them, some are considered as wood–soil interface feeders and the others as true soil-feeders (“group III” and “group IV”, respectively, in [
2]). The trophic niches of those species, besides the fact that they feed on organic material near the end of the humification gradient, are not well known.
The species of the mound-building genera
Thoracotermes (four species) and
Cubitermes (67 valid taxa listed in [
3]) are known as true soil-feeders; these genera and some others of the same subfamily Cubitermitinae are good candidates for tackling the problem of trophic niche differentiation.
It has already been mentioned that two species of
Cubitermes can frequently coexist in the same biotope: Williams [
4] observed that
C. ugandensis Fuller frequently shares the same biotope near Kampala, Uganda, with either
C. orthognathus (Emerson) or
C. testaceus Williams, suggesting that they might occupy somewhat different ecological niches. Bouillon & Vincke [
5] suggested that the
Cubitermes’ diets might be linked with the morphologies of their enteric valves and, indeed, when looking at Williams’ data,
C. ugandensis on the one hand has a valve of the
fungifaber pattern whereas either
C. orthognathus or
C. testaceus, on the other hand, have valves of the
sankurensis pattern [
6]. The same kind of association was found at Bondoé, Republic of Central Africa: in some grassy savannas, two species previously identified as
C. ugandensis and
C. sankurensis Wasmann, coexist with high densities (more than 1200 mounds per hectare) [
7].
Ji et al. [
8] showed that
Cubitermes orthognathus can partly assimilate radiolabeled synthetic humic acids but they did not mention any difference with
C. umbratus Williams or
Thoracotermes macrothorax Sjöstedt. Similarly, no difference was observed between
C. fungifaber (Sjöstedt),
C. heghi Sjöstedt, and
Th. macrothorax in their stable isotope signatures [
9].
This paper relates some preliminary results that were obtained at Bondoé, in November 2010; unfortunately, the civil war in the Republic of Central Africa prevented us, until now, to extend this research.
2. Material and Methods
The observations and sampling occurred in a grassy savannah at Bondoé, Republic of Central Africa (5°10′ N, 17°44′ E); the soil is shallow on a lateritic crust and can be occasionally flooded during the rainy season, but local small beaches offer deeper soil and many
Cubitermes nests are located on those patches (more details in [
7]).
Two nests, located on patches of relatively deep soil (about 30 cm), were chosen, the one built by
C. aff. ugandensis, the other by
C. aff. sankurensis. These species were identified as
C. ugandensis and
C. sankurensis in [
7], but an ongoing revision and an ongoing phylogeny study of
Cubitermes suggest that they belong to nearby, somewhat different species. The nest made by
C. aff. sankurensis was 50 cm high, 25 cm in diameter, and had a single cap 45 cm in diameter; it is therefore evaluated to house about 35,000 individuals [
10]. An annular trench, 25 cm deep, was dug around its base; the nest was examined 24 and 48 h later. The nest of
C. aff. ugandensis was 30 cm high, 30 cm in diameter, and had two caps of about 40 cm in diameter; it is thus evaluated to contain about 30,000 individuals. This nest was also surrounded by an annular trench, 25 cm deep, and examined 24 and 48 h later.
Four other mounds, with a preference for recent nests, were identified to contain either C. aff. sankurensis (two nests) or C. aff. ugandensis (two nests). Recent mounds were chosen because they were expected to contain, each, only one species. Nevertheless, one of the C. aff. sankurensis mounds was somewhat older and appeared to house also a society of Adaiphrotermes sp.
A part of each of the nests was then sorted and samples of 20 to 30 workers, about 10 soldiers and 10 to 15 large larvae (and/or nymphs of alates) were collected and preserved in vials containing dry silica gel. Some nest fragments (about 10 g dry mass) were sampled and preserved the same way. The Adaiphrotermes sp. and the walls surrounding their galleries were also sampled. In the laboratory, the heads of the workers and the soldiers were cut off their bodies and preserved separately with dry silica gel.
The samples were sent to the Kompetenzzentrum Stabile Isotope, Büsgen-Institut, Georg-August-Universität Göttingen for analysis of the C and N stable isotopes. The international standards for nitrogen and carbon are atmospheric nitrogen and Vienna Pee Dee Belemnite, respectively.
3. Results and Discussion
3.1. Depth of Foraging
The C. aff. ugandensis nest, around which a trench had been dug, showed, 24 and 48 h later, two covered galleries that were coming out of the nest at depths of 2 and 4 cm (therefore, near the soil surface) whereas the C. aff. sankurensis nest showed three covered galleries that were coming out of the nest at depths of 9, 20, and 21 cm, respectively. This preliminary result suggests that C. aff. ugandensis would forage near the surface of the soil whereas C. aff. sankurensis would forage in deeper layers.
Even if it is based on two single observations, this is consistent with Williams’ observations near Kampala, Uganda: in that case,
C. ugandensis coexists with
C. testaceus (that has the same kind of enteric valve as
C. sankurensis) and, there also,
C. ugandensis tends to forage near the soil surface whereas
C. testaceus does it “more randomly from the top to the lower parts of the topsoil” [
4], p. 117.
It sounds logical that C. aff. ugandensis, whose workers and soldiers are much larger and possibly also much stronger than either C. sankurensis or C. testaceus, outcompetes the smaller species from the upper layers of soil which are richer in organic matter.
3.2. Stable Isotopes
3.2.1. δ13C
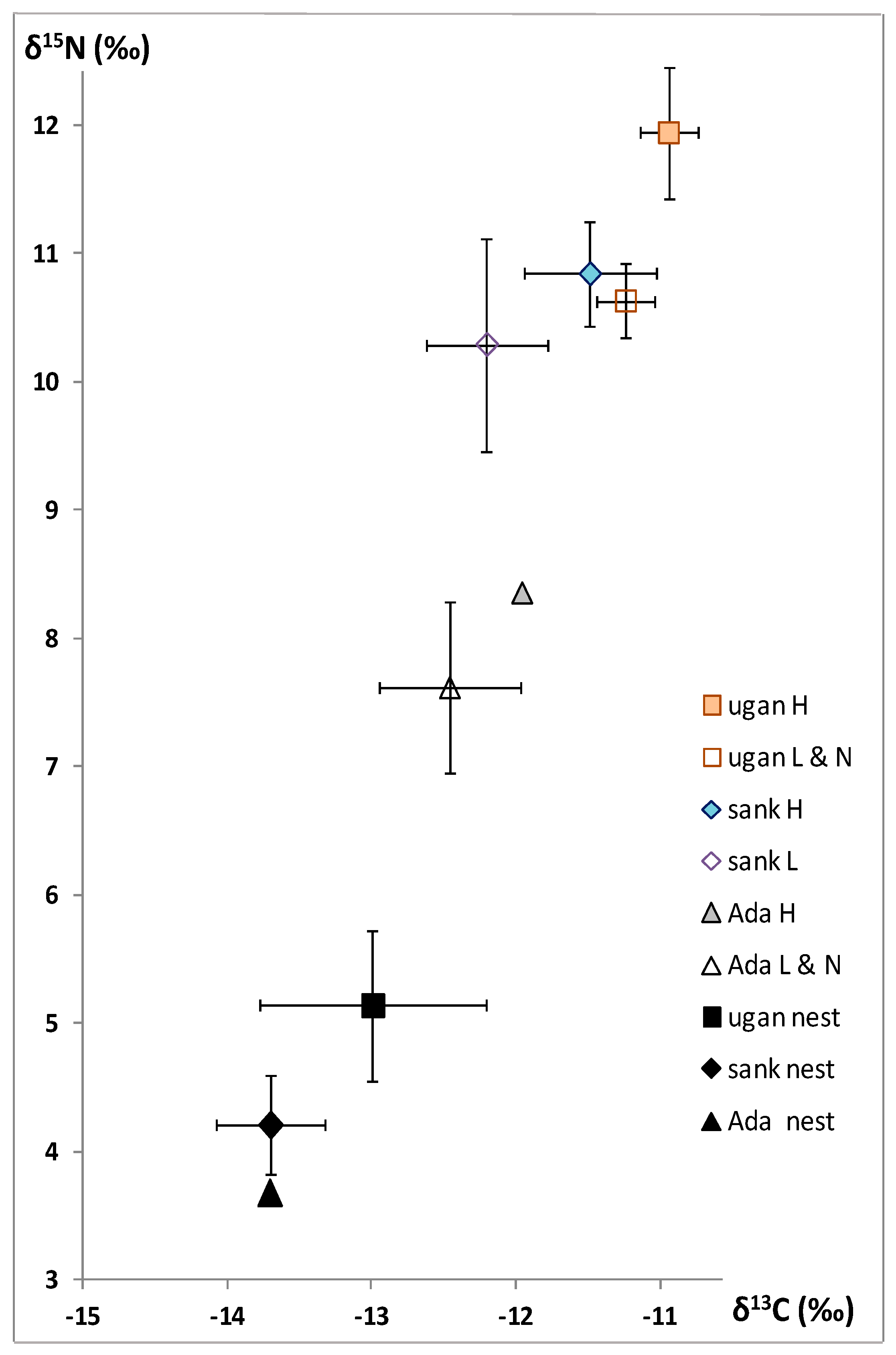
In all the samples, the high δ
13C values (between −13 and −10 in termites; between −14 to −12 in nest material) characterize soil-feeding and a grass food source [
11,
12], in this case, rather organic matter derived from grass (
Figure 1).
In the worker heads, soldier heads, larvae, and alate nymphs, a tendency emerges:
C. aff. ugandensis >
C. aff. sankurensis >
Adaiphrotermes sp., significantly different between
aff.. ugandensis and
aff.. sankurensis (
p < 0.05,
t-test). The same tendency is observed in the nest material, at least between
C. aff. ugandensis and
C. aff. sankurensis, but
Adaiphrotermes established in a mound of
C. aff. sankurensis did not change the δ
13C value (
Figure 1). Within each
Cubitermes species, there is no difference between workers and soldiers (which are fed the same soil as workers) and their values have therefore been pooled in
Figure 1; the larvae and alate nymphs (which are saliva-fed) display lower δ
13C.
3.2.2. δ15N
As expected for humivorous species, high δ15N values (between 7 and 12.5) were observed; actually between 9.5 and 12.5 in the true soil-feeders Cubitermes spp., and less high values (between 7 and 8.5) in the wood-soil interface feeder Adaiphrotermes sp.
In the worker and soldier heads, as well as larvae and alate nymphs, clear and consistent differences emerge:
C. aff. ugandensis >
C. aff. sankurensis >>
Adaiphrotermes sp., significant between
aff.. ugandensis and
aff.. sankurensis (
p < 0.01,
t-test). The same tendency is observed in nest material, at least between
C. aff. ugandensis and
C. aff. sankurensis, but
Adaiphrotermes, established in a mound of
C. aff. sankurensis, shows the same δ
15N value (
Figure 1). The same tendency was observed in the abdomens whose δ
13C and δ
15N are lower than in the heads; however, these results are not shown in
Figure 1 and difficult to interpret because we do not know which proportions of C or N correspond to termite tissues and to gut content.
In both
Cubitermes species, there is no difference between workers and soldiers and their values have therefore been pooled in
Figure 1; the larvae and alate nymphs display some lower δ
15N.
The difference between
Adaiphrotermes sp. (feeding group III), on the one hand, and both
Cubitermes species (feeding group IV), on the other hand, was expected, as it was already observed in Thailand and Cameroon for feeding groups III and IV [
11,
13]. However, since δ
15N and δ
13C are higher in deeper layers than in leaf litter and fermentation layers (more mineralized organic matter) [
12,
13,
14,
15], it sounds contradictory that
C. aff. ugandensis (which forages in upper layers) shows higher δ
15N (and probably also δ
13C) in workers and soldiers, when compared with
C. aff. sankurensis (which forages in deeper layers).
4. Conclusions
The best possible explanation is that these species feed on different organic materials highlighting, for the first time, a trophic niche differentiation between two sympatric true soil feeders bearing two different enteric valve patterns. However, 15N and 13C signatures rely on several factors: exact nature of crude food, termite gut microbiome functions, 15N and 13C signatures of feeding saliva, etc. The “Bondoé bispecific model” provides in this scope promising future research perspectives.
Author Contributions
G.J. designed the study. S.P.M.W. and G.J. collected the material and contributed significantly to the manuscript and approved the final version.
Funding
This study was funded by the Coopération Universitaire au Développement, Belgium.
Acknowledgments
The authors thank J. Deligne for a critically reading of a previous version and two anonymous reviewers for their stimulating comments.
Conflicts of Interest
We declare we have no competing interests.
References
- Noirot, C. From wood- to humus-feeding: An important trend in termite evolution. In Biology and Evolution of Social Insects; Billen, J., Ed.; Leuven University Press: Leuven, Belgium, 1992; pp. 109–117. [Google Scholar]
- Donovan, S.E.; Eggleton, P.; Bignell, D.E. Gut content analysis and a new feeding group classification of termites (Isoptera). Ecol. Entomol. 2001, 26, 356–366. [Google Scholar] [CrossRef]
- Krishna, K.; Grimaldi, D.A.; Krishna, V.; Engel, M.S. Treatise of the Isoptera of the world. Bull. Am. Mus. Nat. Hist. 2013, 377, 1–2704. [Google Scholar] [CrossRef]
- Williams, R.M.C. The East African termites of the genus Cubitermes (Isoptera: Termitidae). Trans. R. Entomol. Soc. Lond. 1966, 118, 73–118. [Google Scholar] [CrossRef]
- Bouillon, A.; Vincke, P.P. Valvule entérique et révision du genre Cubitermes Wasmann. Cubitermes bugeserae sp. nov. (Isoptera, Termitidae). Revue de Zoologie et de Botanique Africaines 1971, 84, 269–280. [Google Scholar]
- Josens, G.; Deligne, J. Species groups in the genus Cubitermes (Isoptera: Termitidae) defined on the basis of enteric valve morphology. Eur. J. Taxon. 2019, in press. [Google Scholar]
- Wango, S.P.; Josens, G. Comparison of nest shapes and densities of two sympatric species of Cubitermes (Isoptera: Termitidae: Termitinae) as clues for the study of their population dynamics. Afr. Zool. 2011, 46, 156–168. [Google Scholar] [CrossRef]
- Ji, R.; Kappler, A.; Brune, A. Transformation and mineralization of synthetic 14C-labeled humic model compounds by soil-feeding termites. Soil Biol. Biochem. 2000, 32, 1281–1291. [Google Scholar] [CrossRef]
- Tayasu, I.; Abe, T.; Eggleton, P.; Bignell, D.E. Nitrogen and carbon isotope ratios in termites (Isoptera): An indicator of trophic habit along the gradient from wood-feeding to soil-feeding. Ecol. Entomol. 1997, 22, 343–351. [Google Scholar] [CrossRef]
- Josens, G.; Soki, K. Relation between termite numbers and the size of their mounds. Insectes Sociaux 2010, 57, 303–316. [Google Scholar] [CrossRef]
- TayasuI, I. Use of carbon and nitrogen isotope ratios in termite research. Ecol. Res. 1998, 13, 377–387. [Google Scholar] [CrossRef]
- Schwartz, D.; Mariotti, A.; Lanfranchi, R.; Guillet, B. 13C/12C ratios of soil organic matter as indicators of vegetation changes in the Congo. Geoderma 1986, 39, 97–103. [Google Scholar] [CrossRef]
- Tayasu, I.; Nakamura, T.; Oda, H.; Hyodo, F.; Takematsu, Y.; Abe, T. Termite ecology in a dry evergreen forest in Thailand in terms of stable (d13C and d15N) and radio (14C, 137Cs and 210Pb) isotopes. Ecol. Res. 2002, 17, 195–206. [Google Scholar] [CrossRef]
- Tiunov, A.V. Stable Isotopes of Carbon and Nitrogen in Soil Ecological Studies. Biol. Bull. 2007, 34, 395–407. [Google Scholar] [CrossRef]
- Potapov, A.M.; Tiunov, A.V.; Scheu, S. Uncovering trophic positions and food resources of soil animals using bulk natural stable isotope composition. Biol. Rev. 2018, 94, 37–59. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).


{kind=link}
