Temperature-Mediated Variations in Behavior and Mortality Caused by Non-Repellent Insecticides in Subterranean Termites (Blattodea: Rhinotermitidae)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Organism
2.2. Chemicals
2.3. Bioassay
2.4. Observation of Behavioral Symptoms
2.5. Statistical Analysis
3. Results
3.1. Mortality
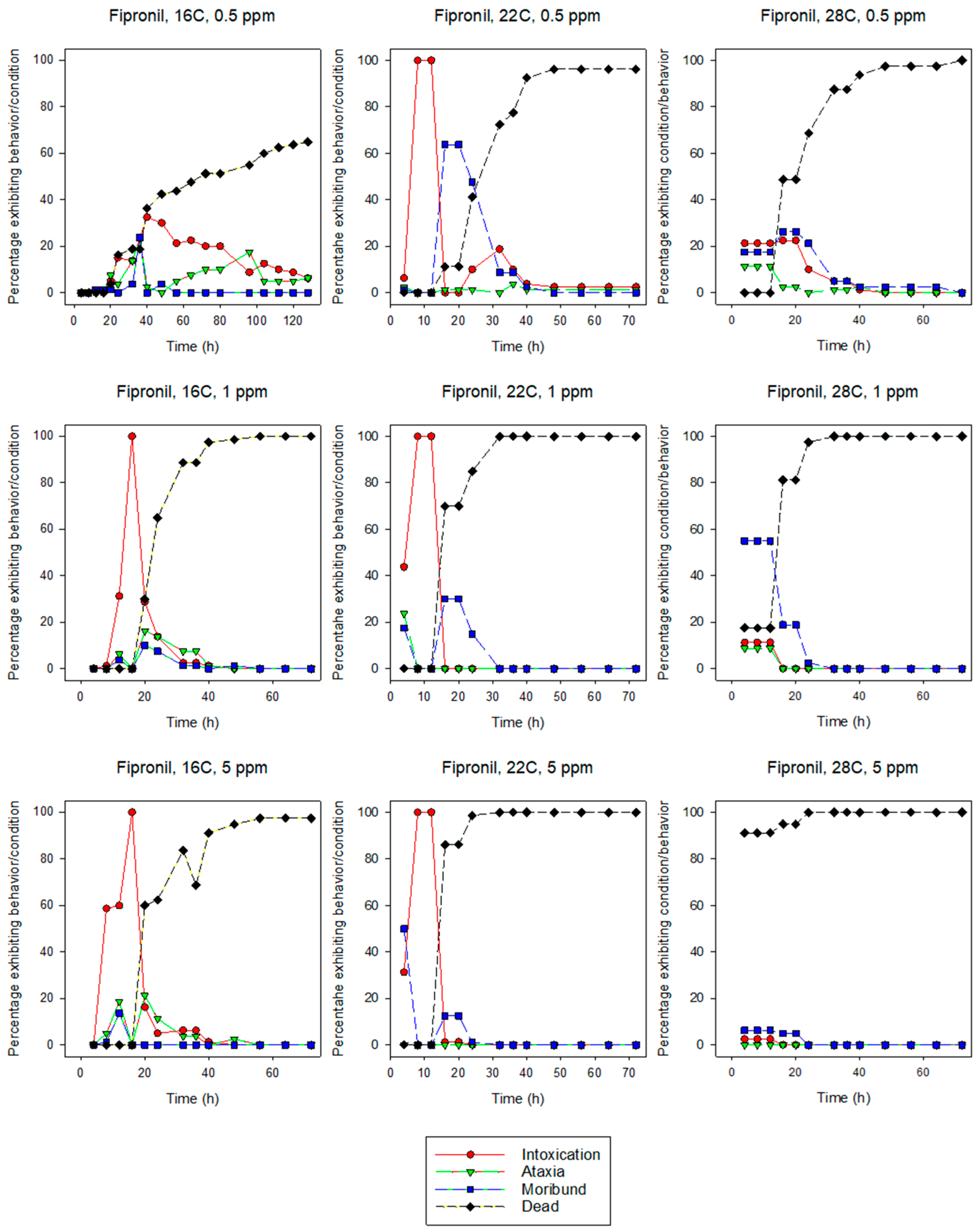
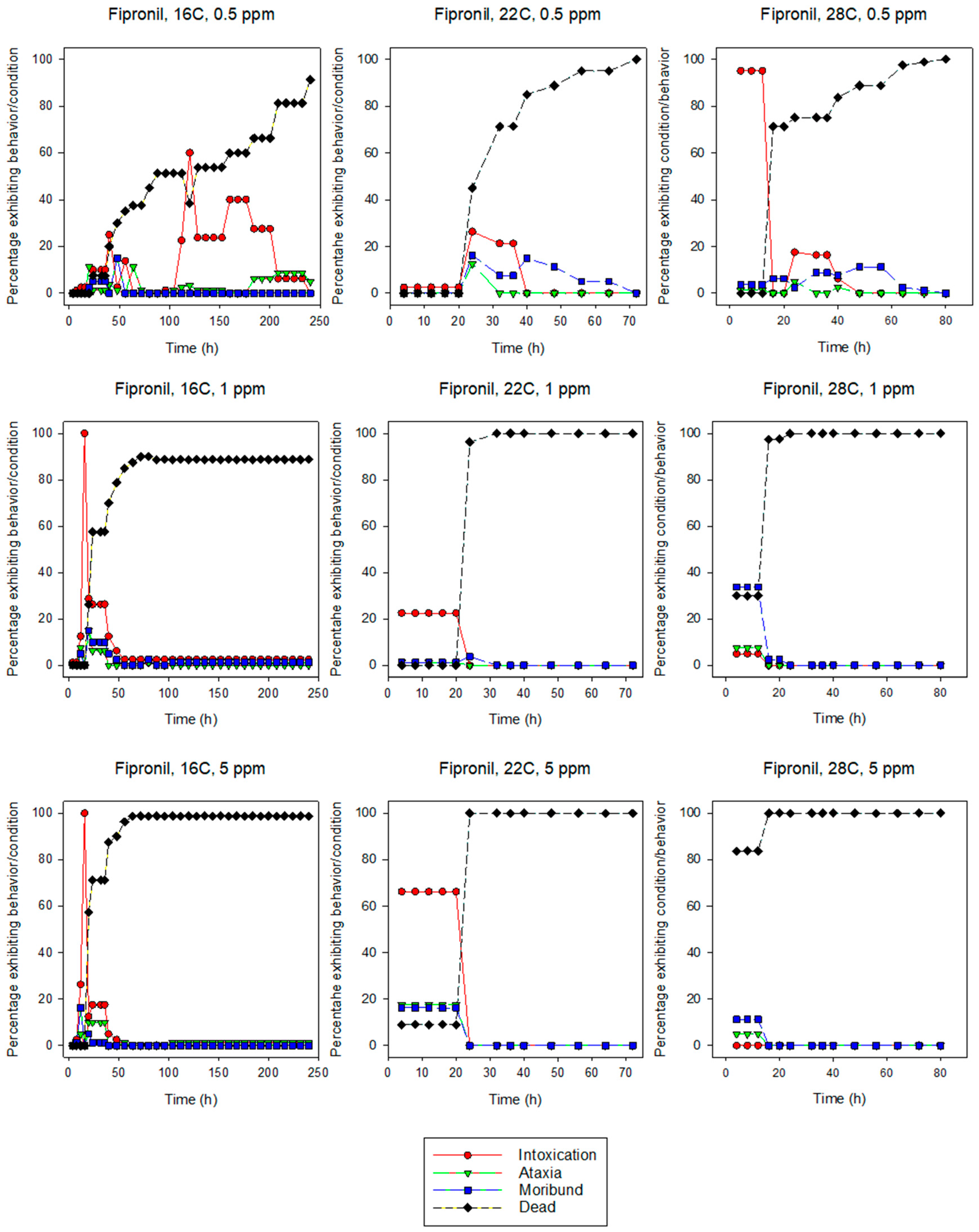
3.1.1. Fipronil
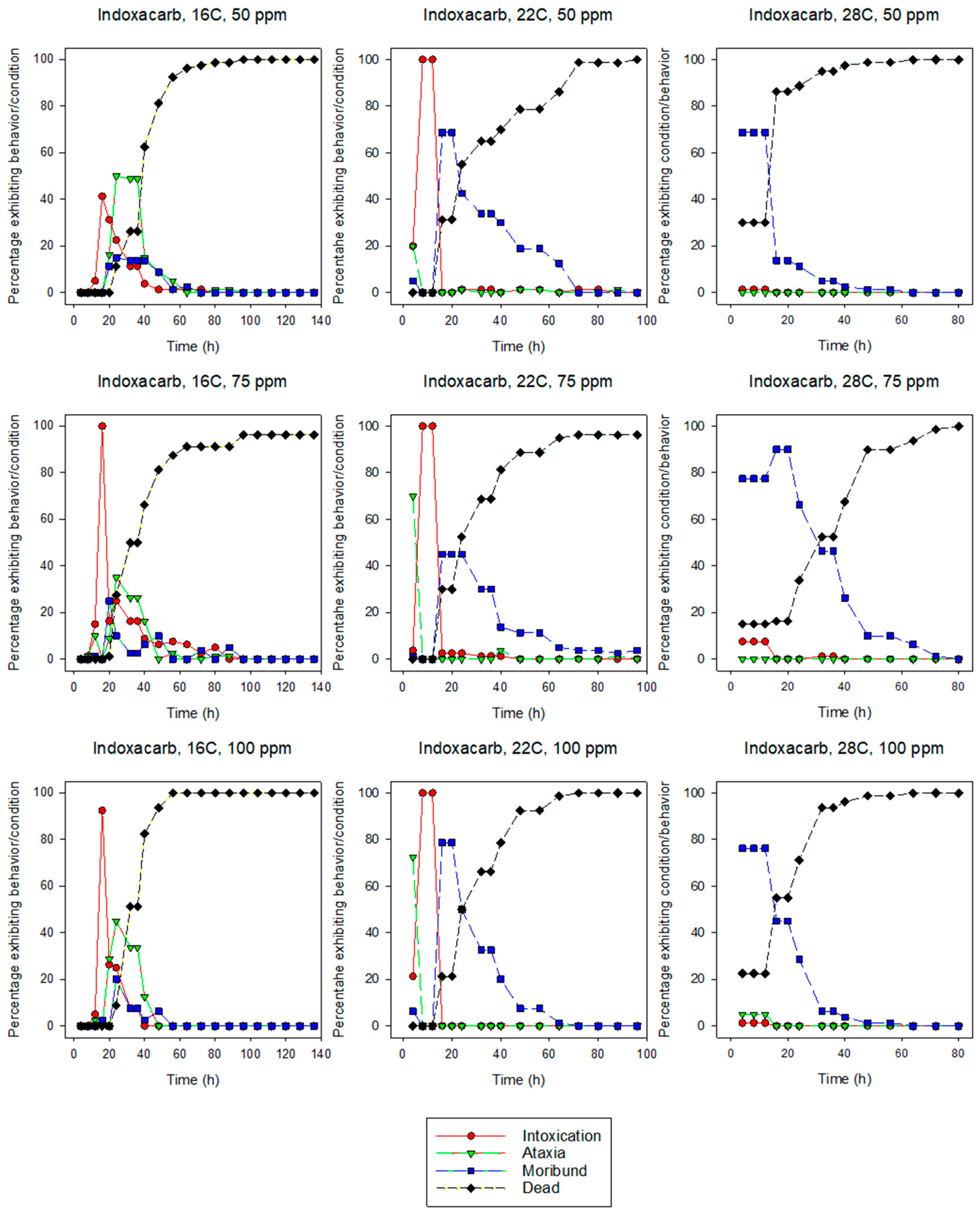
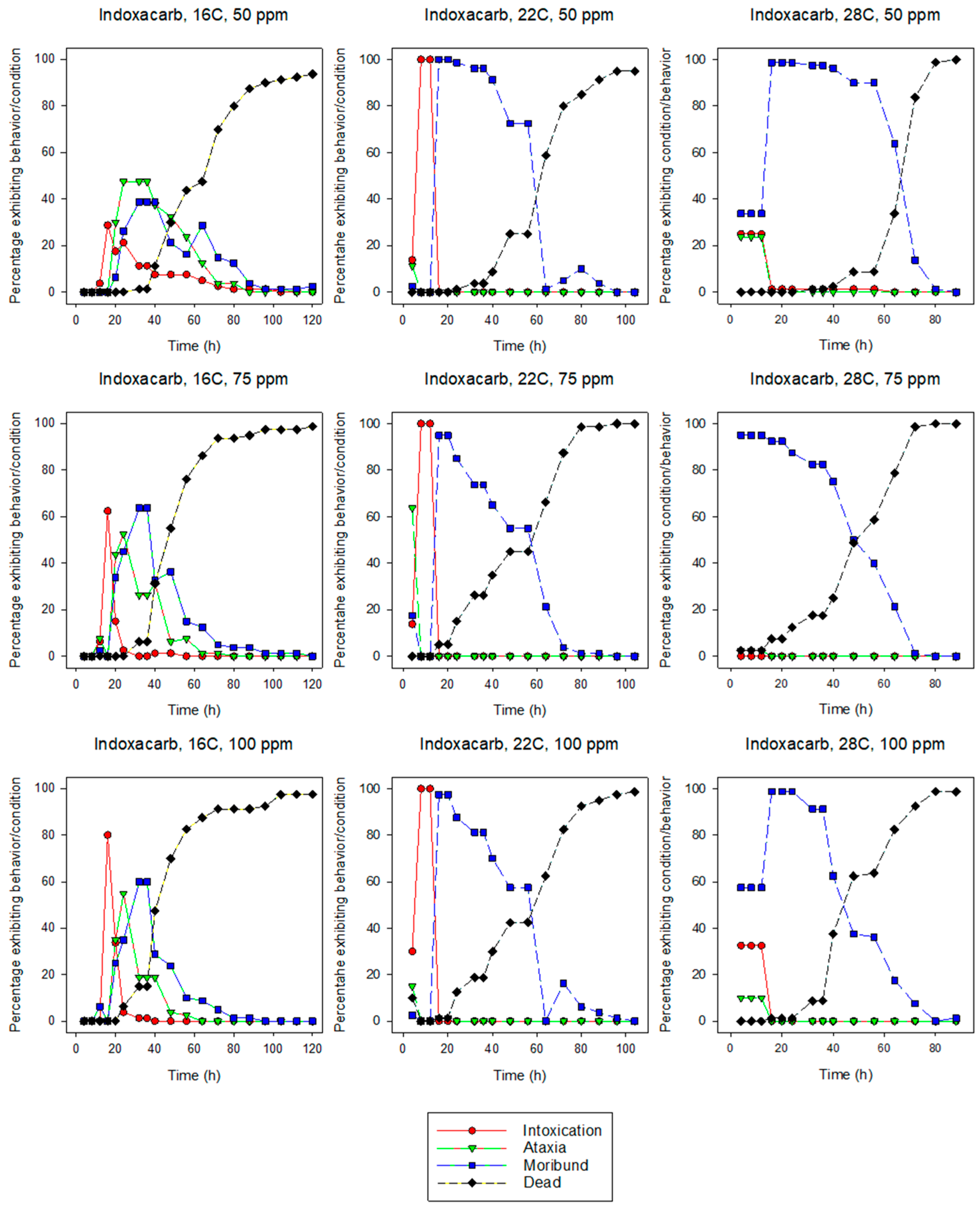
3.1.2. Indoxacarb
3.2. Behavior
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Su, N.-Y.; Scheffrahn, R.H. Termites as pest of buildings. In Termites: Evolution, Sociality, Symbiosis, and Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 437–453. [Google Scholar]
- Hu, X.P. Liquid insecticides: Their role in subterranean termite management. In Urban Pest Management: An Environmental Perspective; Dhang, P., Ed.; CAB International: Cambridge, MA, USA, 2011; pp. 114–132. [Google Scholar]
- Hu, X.P.; Hickman, B. Exterior perimeter plus limited interior treatments with fipronil as an IPM option for subterranean termite management. Int. Pest Control 2006, 48, 200–203. [Google Scholar]
- Hu, X.P.; Song, D.; Presley, W. Horizontal and vertical transfer of fipronil within functional colonies. In Proceedings of the 2006 National Conference on Urban Entomology, Raleigh, NC, USA, 21–24 May 2006; pp. 39–44. [Google Scholar]
- Quarcoo, F.Y.; Appel, A.G.; Hu, X.P. Effects of indoxacarb concentration and exposure time on onset of abnormal behaviors, morbundity, and death in the eastern subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2010, 103, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Indrayani, Y.; Yoshimura, T.; Yanase, Y.; Fujii, Y.; Imamura, Y. Evaluation of the temperature and relative humidity preferences of the western dry-wood termite Incisitermes minor (Hagen) using acoustic emission (AE) monitoring. J. Wood Sci. 2007, 53, 76–79. [Google Scholar] [CrossRef]
- Strack, B.H.; Myles, T.G. Behavioral responses of the eastern subterranean termite to falling temperatures (Isoptera: Rhinotermitidae). Proc. Entomol. Soc. Ont. 1997, 128, 13–17. [Google Scholar]
- Rudolph, D.; Glocke, B.; Rathenow, S. On the role of the different humidity parameters for the survival distribution and ecology of various termite species. Socibiology 1990, 17, 129–140. [Google Scholar]
- Appel, A.G.; Reierson, D.A.; Rust, M.K. Comparative water relations and temperature sensitivity of cockroaches. Comp. Biochem. Physiol. 1983, 74, 357–361. [Google Scholar] [CrossRef]
- Sponsler, R.C.; Appel, A.G. Temperature tolerances of the Formosan and eastern subterranean termites (Isoptera: Rhinotermitidae). J. Therm. Biol. 1991, 16, 41–44. [Google Scholar] [CrossRef]
- Hu, X.P.; Appel, A.G. Seasonal variation of critical thermal limits and temperature tolerance in two subterranean termites (Isoptera: Rhinotermitidae). Environ. Entomol. 2004, 33, 197–205. [Google Scholar] [CrossRef]
- Woodrow, R.J.; Grace, J.K. Thermal tolerances of four termite species (Isoptera: Rhinotermitidae, Kalotermitidae). Sociobiology 1998, 32, 17–25. [Google Scholar]
- Smith, J.L.; Rust, M.K. Influence of temperature on tunneling, feeding rates, and oxygen requirements of the western subterranean termite, Reticulitermes hesperus (Isoptera: Rhinotermitidae). Sociobiology 1993, 21, 225–236. [Google Scholar]
- Smith, J.L.; Rust, M.K. Effect of relative humidity and temperature on the survival of Reticulitermes hesperus (Isoptera: Rhinotermitidae). Sociobiology 1993, 21, 217–224. [Google Scholar]
- Esenther, G.R. Subterranean termite studies in southern Ontario. Can. Entomol. 1969, 100, 827–834. [Google Scholar] [CrossRef]
- Cabrera, B.J.; Kamble, S.T. Effects of decreasing thermophotoperiod on the eastern subterranean termite (Isoptera: Rhinotermitidae). Environ. Entomol. 2001, 30, 166–171. [Google Scholar] [CrossRef]
- Hu, X.P.; Song, D. Behavioral responses of two subterranean termite species (Isoptera: Rhinotermitidae) to instant freezing or chilling temperatures. Environ. Entomol. 2007, 36, 1450–1456. [Google Scholar] [CrossRef]
- Hadley, N.F. Adaptational biology of desert scorpions. J. Arachnol. 1974, 2, 11–23. [Google Scholar]
- Bryson, R.A.; Hare, F.K. Climates of North America. In World Survey of Climatology, Volume 2; Landsberg, H.E., Ed.; Elsevier: Amsterdam, The Netherlands, 1974; p. 420. [Google Scholar]
- Delaplane, K.S. Foraging and feeding behaviors of the Formosan subterranean termite (Isoptera: Rhinotermitidae). Sociobiology 1991, 19, 101–114. [Google Scholar]
- Wiltz, B.A. Effects of temperature and humidity on survival of Coptotermes formosanus and Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiology 2012, 59, 381–394. [Google Scholar] [CrossRef]
- Fei, H.; Henderson, G. Effects of temperature, directional aspects, light conditions and termite species on subterranean termite activity (Isoptera: Rhinotermitidae). Environ. Entomol. 2004, 33, 242–248. [Google Scholar] [CrossRef]
- King, E.G., Jr.; Spink, W.L. Laboratory studies on the biology of the Formosan subterranean termite with primary emphasis on young colony development. Ann. Entomol. Soc. Am. 1974, 67, 953–958. [Google Scholar] [CrossRef]
- Honigsberg, B.M. Protozoa associated with termites and their role in digestion. In Biology of Termites; Krishna, K., Weesner, F.M., Eds.; Academic Press: New York, NY, USA, 1970; pp. 1–36. [Google Scholar]
- Fei, H.; Henderson, G. Formosan subterranean termite (Isoptera: Rhinotermitidae) wood consumption and worker survival as affected by temperature and soldier proportion. Environ. Entomol. 2002, 31, 509–514. [Google Scholar] [CrossRef]
- Satpute, N.S.; Deshmukh, S.D.; Rao, N.G.V.; Tikar, S.N.; Moharil, M.P.; Nimbalkar, S.A. Temperature-dependent variation in toxicity of insecticides against Earias vitella (Lepidoptera: Noctuidae). J. Econ. Entomol. 2007, 100, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Silver, K.S.; Soderlund, D.M. Action of pyrazoline-type insecticides at neuronal target sites. Pest. Biochem. Physiol. 2005, 81, 136–143. [Google Scholar] [CrossRef]
- Sheets, J.J.; Karr, L.L.; Dripps, J.E. Kinetics of uptake, clearance, transfer and metabolism of hexaflumuron by eastern subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2000, 93, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Campbell, N.A.; Reece, J.B.; Urry, L.A.; Cain, M.L.; Wasserman, S.A.; Minorsky, P.V.; Jackson, R.B. Biology, 8th ed.; Pearson Benjamin Cummings: San Francisco, CA, USA, 2009; p. 1267. [Google Scholar]
- Spomer, N.A.; Kamble, S.T. Temperature effect on kinetics of uptake, transfer and clearance of [14C] Noviflumuron in eastern subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2006, 99, 134–140. [Google Scholar] [CrossRef]
- Spomer, N.A.; Kamble, S.T. Effect of temperature on noviflumuron performance against the eastern subterranean termite (Isoptera: Rhinotermitidae). Sociobiology 2005, 46, 335–348. [Google Scholar]
- U.S. Environmental Protection Agency. Indoxacarb Pesticide Fact Sheet, Office of Prevention, Pesticides and Toxic Substances (7505C); U.S. Environmental Protection Agency: Washington, DC, USA, 2000.
- Lainé, L.; Wright, D. The life cycle of Reticulitermes spp. (Isoptera: Rhinotermitidae): What do we know? Bull. Entomol. Res. 2003, 93, 267–278. [Google Scholar] [CrossRef]
- Quarcoo, F.Y.; Appel, A.G.; Hu, X.P. Descriptive study of non-repellent insecticide-induced behaviors in Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiology 2010, 55, 217–227. [Google Scholar]
- SAS Institute. SAS User’s Guide, Version 8.2; SAS Institute: Cary, NC, USA, 2001. [Google Scholar]
- Smythe, R.V.; Williams, L.H. Feeding and survival of two subterranean termite species at constant temperatures. Ann. Entomol. Soc. Am. 1972, 65, 226–229. [Google Scholar] [CrossRef]
- Cao, R.; Su, N.-Y. Tunneling and food transportation activity of four subterranean termite species (Isoptera: Rhinotermitidae) at various temperatures. Ann. Entomol. Soc. Am. 2014, 107, 696–701. [Google Scholar] [CrossRef]
- Hu, X.P. Evaluation of efficacy and nonrepellency of indoxacarb and fipronil-treated soil at various concentrations and thicknesses against two subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2005, 98, 509–517. [Google Scholar] [CrossRef]
- Quarcoo, F.Y.; Hu, X.P.; Appel, A.G. Effects of non-repellent termiticides on the tunneling and walking ability of the eastern subterranean termite (Isoptera: Rhinotermitidae). Pest Manag. Sci. 2012, 68, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.G. Effects of temperature on insecticides toxicity. In Reviews in Pesticide Toxicology; Roe, R.M., Kuhr, R.J., Eds.; Toxicology Communications Inc.: Raleigh, NC, USA, 1995; Volume 3, pp. 111–135. [Google Scholar]
- Scharf, M.E.; Siefried, B.D.; Meinke, L.J.; Chandler, L.D. Fipronil metabolism, oxidative sulfone formation and toxicity among organophosphate- and carbamate-resistant and susceptible western corn rootworm populations. Pest Manag. Sci. 2000, 56, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Su, N.-Y.; Tamashiro, M.; Yates, J.R.; Haverty, M.I. Effect of behavior on the evaluation of insecticides for prevention of remedial control of the Formosan subterranean termite. J. Econ. Entomol. 1982, 75, 188–193. [Google Scholar] [CrossRef]
- Shelton, T.G. Distance of the repellency of dead Reticulitermes flavipes (Isoptera: Rhinotermitidae) nestmates. J. Entomol. Sci. 2014, 49, 221–227. [Google Scholar] [CrossRef]
- Shelton, T.G.; Grace, J.K. Effects of exposure duration on transfer of nonrepellent termiticides among workers of Coptotermes formosanus Shiraki (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2003, 96, 456–460. [Google Scholar] [CrossRef]
- Saran, R.K.; Rust, M.K. Toxicity, uptake, and transfer efficiency of fipronil in western subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2007, 100, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Osbrink, W.L.A.; Cornelius, M.L.; Lax, A.R. Effect of imidacloprid soil treatments on the occurrence of Formosan subterranean termites, Coptotermes formosanus Shiraki (Isoptera: Rhinotermitidae), in independent monitors. J. Econ. Entomol. 2005, 98, 2160–2168. [Google Scholar] [CrossRef]
- Aguilar, J.; Monaenkova, D.; Linevich, V.; Savoie, W.; Dutta, B.; Kuan, H.-S.; Betterton, M.D.; Goodisman, M.A.D.; Goldman, D.I. Collective clog control: Optimizing traffic flow in confined biological and robophysical excavation. Science 2018, 361, 672–677. [Google Scholar] [CrossRef]
- Henderson, G. Liquid learning. Pest Contr. Tech. 2003, 31, 48–59. [Google Scholar]
- Song, D.; Hu, X.P. Effects of dose, donor-recipient interaction time and ratio on fipronil transmission among the Formosan subterranean termite nestmates (Isoptera: Rhinotermitidae). Sociobiology 2006, 48, 237–246. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (ppm) | Temperature (°C) | LT50 (h) | 95% FL (h) | χ2 | Slope ± SE |
|---|---|---|---|---|---|
| 0.5 | 16 | 84.92 | 71.47–99.88 | 91.893 | 1.77 ± 9.18 |
| 0.5 | 22 | 26.84 | 24.66–28.95 | 138.107 | 5.79 ± 0.49 |
| 0.5 | 28 | 20.14 | 18.65–21.58 | 181.186 | 5.45 ± 0.40 |
| 1.0 | 16 | 23.65 | 22.73–24.55 | 257.079 | 8.90 ± 0.55 |
| 1.0 | 22 | 16.68 | 15.78–17.54 | 105.292 | 8.89 ± 0.87 |
| 1.0 | 28 | 11.36 | 9.89–12.81 | 108.114 | 4.17 ± 0.40 |
| 5.0 | 16 | 22.97 | 17.80–27.51 | 96.070 | 2.85 ± 0.29 |
| 5.0 | 22 | 15.14 | 14.57–15.68 | 119.117 | 13.36 ± 1.22 |
| 5.0 | 28 | 0.96 | 0.37–1.72 | 56.049 | 1.60 ± 0.21 |
| Concentration (ppm) | Temperature (°C) | LT50 (h) | 95% FL (h) | χ2 | Slope ± SE |
|---|---|---|---|---|---|
| 0.5 | 16 | 98.78 | 83.65–115.78 | 69.564 | 2.46 ± 0.29 |
| 0.5 | 22 | 29.12 | 27.33–30.83 | 135.573 | 7.54 ± 0.65 |
| 0.5 | 28 | 19.20 | 16.34–21.90 | 87.952 | 3.96 ± 0.42 |
| 1.0 | 16 | 31.73 | 26.31–37.10 | 194.653 | 1.96 ± 0.14 |
| 1.0 | 22 | 23.16 | 22.92–23.39 | 3.61 × 105 | 114.44 ± 0.00 |
| 1.0 | 28 | 8.57 | 7.28–9.84 | 94.090 | 3.79 ± 0.39 |
| 5.0 | 16 | 22.48 | 17.80–26.97 | 78.932 | 3.30 ± 0.37 |
| 5.0 | 22 | 21.91 | 21.55–22.01 | 0.000 | 212.93 ± 0.00 |
| 5.0 | 28 | 2.02 | 1.06–2.98 | 51.732 | 2.14 ± 0.30 |
| Concentration (ppm) | Temperature (°C) | LT50 (h) | 95% FL (h) | χ2 | Slope ± SE |
|---|---|---|---|---|---|
| 50.0 | 16 | 43.52 | 42.30–44.75 | 292.324 | 9.60 ± 0.56 |
| 50.0 | 22 | 17.33 | 12.56–22.26 | 90.377 | 1.99 ± 0.21 |
| 50.0 | 28 | 6.42 | 4.47–8.41 | 114.016 | 1.87 ± 0.18 |
| 75.0 | 16 | 23.31 | 16.44–30.50 | 79.284 | 1.80 ± 0.20 |
| 75.0 | 22 | 16.65 | 11.45–22.03 | 90.470 | 1.67 ± 0.18 |
| 75.0 | 28 | 16.91 | 12.26–21.72 | 92.779 | 1.94 ± 0.20 |
| 100.0 | 16 | 20.52 | 14.44–27.01 | 65.450 | 2.19 ± 0.28 |
| 100.0 | 22 | 16.91 | 12.29–21.71 | 86.538 | 2.10 ± 0.23 |
| 100.0 | 28 | 7.57 | 5.88–9.30 | 155.352 | 2.03 ± 0.16 |
| Concentration (ppm) | Temperature (°C) | LT50 (h) | 95% FL (h) | χ2 | Slope ± SE |
|---|---|---|---|---|---|
| 50.0 | 16 | 69.1 | 63.94–74.16 | 286.697 | 4.32 ± 0.26 |
| 50.0 | 22 | 39.62 | 26.40–54.52 | 44.193 | 1.85 ± 0.28 |
| 50.0 | 28 | 37.69 | 23.22–54.89 | 31.559 | 2.10 ± 0.37 |
| 75.0 | 16 | 29.94 | 20.21–40.62 | 51.207 | 2.03 ± 0.28 |
| 75.0 | 22 | 28.33 | 19.28–38.21 | 52.846 | 2.06 ± 0.28 |
| 75.0 | 28 | 27.14 | 18.49–36.55 | 54.090 | 2.04 ± 0.28 |
| 100.0 | 16 | 28.34 | 19.52–37.83 | 60.040 | 1.93 ± 0.25 |
| 100.0 | 22 | 31.25 | 21.04–42.45 | 50.172 | 1.97 ± 0.28 |
| 100.0 | 28 | 25.28 | 17.90–33.38 | 67.110 | 2.08 ± 0.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quarcoo, F.Y.; Hu, X.P.; Appel, A.G. Temperature-Mediated Variations in Behavior and Mortality Caused by Non-Repellent Insecticides in Subterranean Termites (Blattodea: Rhinotermitidae). Insects 2019, 10, 37. https://doi.org/10.3390/insects10020037
Quarcoo FY, Hu XP, Appel AG. Temperature-Mediated Variations in Behavior and Mortality Caused by Non-Repellent Insecticides in Subterranean Termites (Blattodea: Rhinotermitidae). Insects. 2019; 10(2):37. https://doi.org/10.3390/insects10020037
Chicago/Turabian StyleQuarcoo, Franklin Y., Xing Ping Hu, and Arthur G. Appel. 2019. "Temperature-Mediated Variations in Behavior and Mortality Caused by Non-Repellent Insecticides in Subterranean Termites (Blattodea: Rhinotermitidae)" Insects 10, no. 2: 37. https://doi.org/10.3390/insects10020037