Consumption Rate of Lichens by Constrictotermes cyphergaster (Isoptera): Effects of C, N, and P Contents and Ratios


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Collection and Sampling Procedures
2.3. Lichen Species Determination
2.4. Macronutrient Quantification
2.5. Lichen Consumption in the Laboratory
2.6. Data Analysis
3. Results
3.1. Lichens Consumed by C. cyphergaster in the Field
3.2. Quantification of Lichen Consumption in the Laboratory
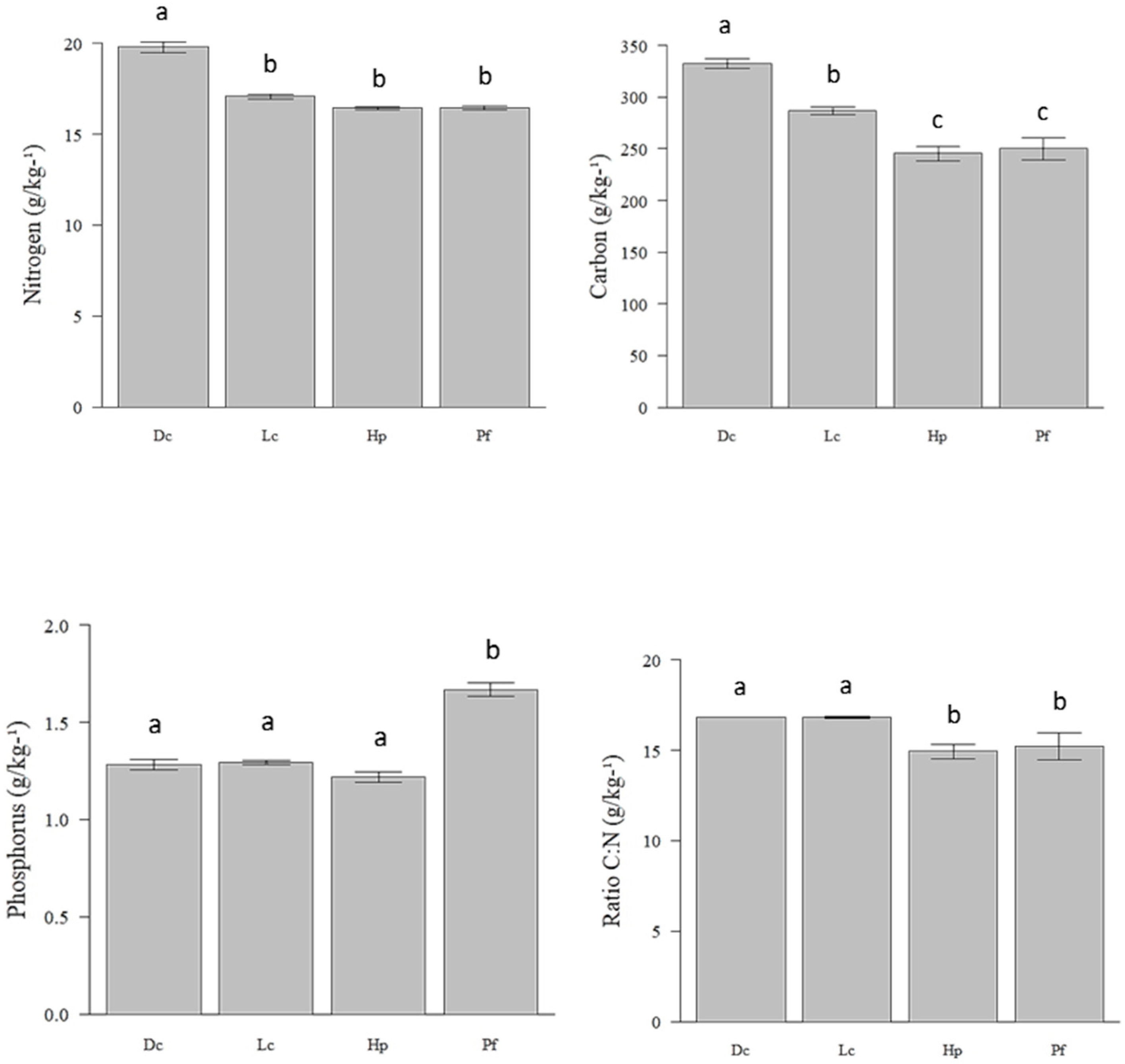
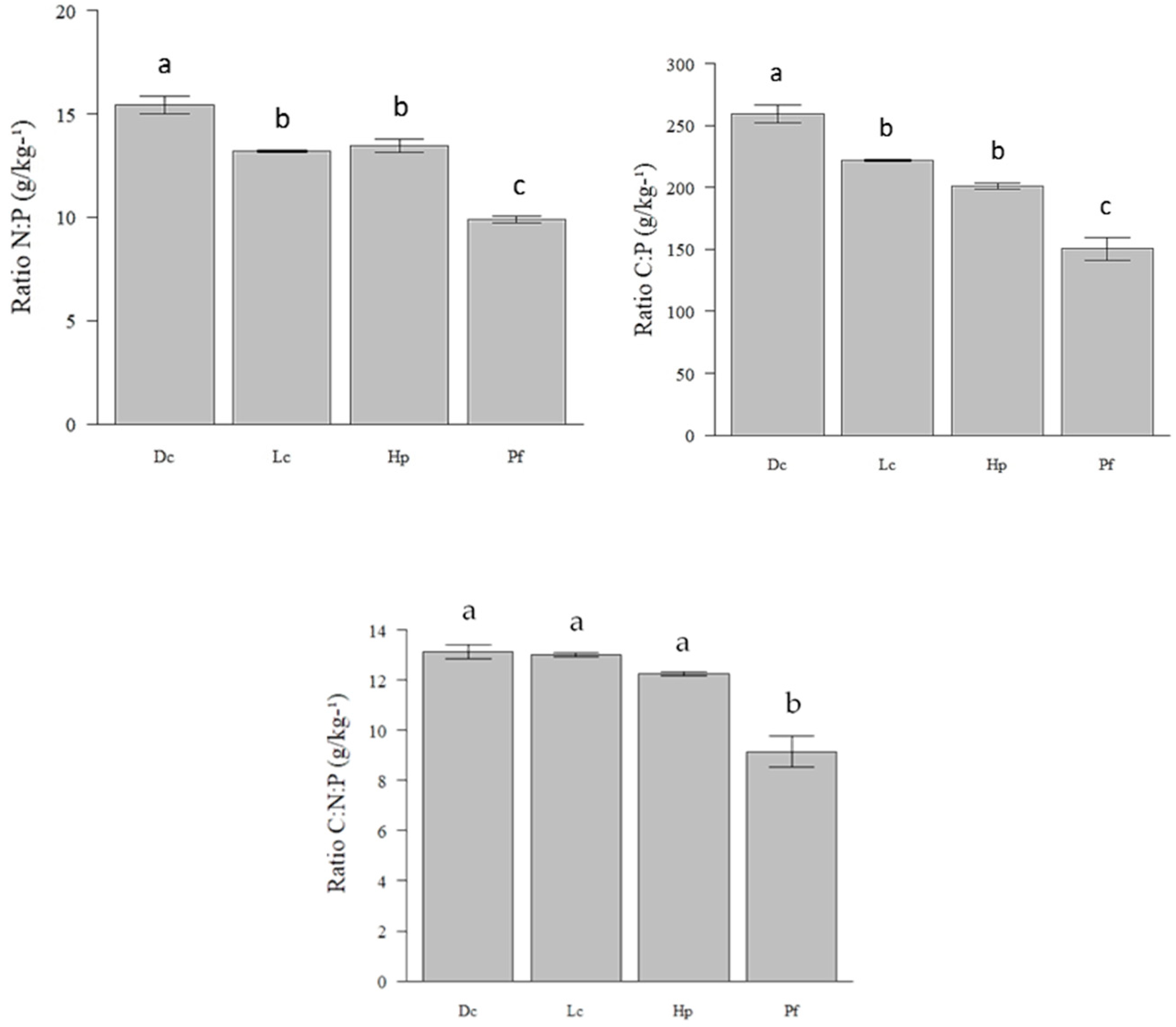
3.3. Concentration of Nutrients in Lichens and Their Effect on Consumption
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wood, T.G.; Sands, W.A. The role of termites in ecosystems. In Production Ecology of Ants and Termites; Brian, M.V., Ed.; Cambridge University Press: Cambridge, UK, 1978; pp. 245–292. [Google Scholar]
- Bignell, D.E.; Eggleton, P. Termites in ecosystems. In Termites: Evolution, Sociality, Symbiosis, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 363–387. [Google Scholar]
- Brauman, A. Effect of gut transit and mound deposit on soil organic matter transformations in the soil feeding termite: A review. Eur. J. Soil Sci. Eur. 2000, 36, 117–125. [Google Scholar] [CrossRef]
- Wood, T.G. Food and feeding habits of termites. In Production Ecology of Ants and Termites; Brian, M.V., Ed.; Cambridge University Press: Cambridge, UK, 1978; pp. 55–80. [Google Scholar]
- Matsumoto, T.; Abe, T. The role of termites in an equatorial rain forest ecosystem of west Malaysia. Oecologia 1979, 38, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Becker, G. Rearing of termites and testing methods used in the laboratory. In Biology of Termites; Krishna, K., Weesner, F.M., Eds.; Academic Press: New York, NY, USA; London, UK, 1969; pp. 351–385. [Google Scholar]
- Lenz, M.; Williams, E.R. Influence of container, matrix volume and group size on survival and feeding activity in species of Coptotermes and Nasutitermes (Isoptera: Rhinotermitidae, Termitidae). Mater. Org. 1980, 15, 25–46. [Google Scholar]
- Lenz, M.; Barret, R.A.; Miller, L.R. The capacity of colonies of Coptotermes acinaciformis acinaciformis from Australia to produce neotenics (Isoptera: Rhinotermitidae). Sociobiology 1986, 11, 237–244. [Google Scholar]
- Lenz, M.; Amburgey, T.L.; Zi-Rong, D.; Mauldin, J.K.; Preston, A.F.; Rudolph, D.; Williams, E.R. Interlaboratory studies on termite wood decay fungi associations. 2. Response of termites to Gloeophyllum trabeum grown on different species of wood (Isoptera: Mastotermitidae, Termopsidae, Rhinotermitidae, Termitidae). Sociobiology 1991, 18, 203–254. [Google Scholar]
- Bustamante, N.C.R.; Martius, C. Nutritional preferences of wood-feeding termites inhabiting floodplain forests of the Amazon River, Brazil. Acta Amazon. 1998, 28, 301–307. [Google Scholar] [CrossRef]
- Botch, P.S.; Brennan, C.L.; Judd, T.M. Seasonal Effects of Calcium and Phosphates on the Feeding Preference of the Termite Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiology 2010, 55, 489–498. [Google Scholar]
- Vasconcellos, A.; Moura, F.M.S. Wood litter consumption by three species of Nasutitermes termites in an area of the Atlantic Coastal Forest in Northeastern Brazil. J. Insect Sci. 2010. [Google Scholar] [CrossRef]
- Barca, R.R.B.; Lucena, E.F.; Vasconcellos, A. Nest Population Structure and Wood Litter Consumption by Microcerotermes indistinctus (Isoptera) in a Seasonally Dry Tropical Forest, Northeastern Brazil. Insects 2018, 9, 97. [Google Scholar] [CrossRef]
- Waller, D.A.; Jones, C.G.; La Fage, J.P. Measuring Wood preference in termites. Entomol. Esp. Appl. 1990, 56, 117–123. [Google Scholar] [CrossRef]
- Curtis, A.D.; Waller, D.A. Variation in rates of nitrogen fixation in termites: Response to dietary nitrogen in the field and laboratory. Physiol. Entomol. 1997, 22, 303–309. [Google Scholar] [CrossRef]
- Waller, D.A.; Curtis, A.D. Effects of Sugar-Treated Foods on Preference and Nitrogen Fixation in Reticulitermes flavipes (Kollar) and Reticulitermes virginicus (Banks) (Isoptera: Rhinotermitidae). Ann. Entomol. Soc. Am. 2003, 96, 81–85. [Google Scholar] [CrossRef]
- Noirot, C. From wood- to humus feeding: An important trend in termite evolution. In Biology and Evolution of Social Insects; Billen, J., Ed.; Leuven University Press: Leuven, Belgium, 1992; pp. 107–119. [Google Scholar]
- Mathews, A.G.A. Studies on Termites from the Mato Grosso State, Brazil; Academia Brasileira de Ciências: Rio de Janeiro, Brazil, 1977. [Google Scholar]
- Donovan, S.E.; Eggleton, E.; Bignell, D.E. Gut content analysis and a new feeding group classification of termites. Ecol. Entomol. 2001, 26, 356–366. [Google Scholar] [CrossRef]
- Nalepa, C.A. Nourishment and the origin of termite eusociality. In Nourishment and Evolution in Insect Societies; Hunt, J.H., Nalepa, C.A., Eds.; Westview Press: Oxford, UK, 1994; pp. 57–104. [Google Scholar]
- Higashi, M.; Abe, T.; Thomas, P.; Burns, T.P. Carbon—nitrogen balance and termite ecology. Proc. R. Soc. Land. B 1992, 249, 303–308. [Google Scholar] [CrossRef]
- Judd, T.M. Cues Used by Subterranean Termites During Foraging and Food Assessment. In Termites and Sustainable Management: Biology, Social Behaviour and Economic Importance; Khan, A., Ahmad, W., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 159–180. [Google Scholar]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef]
- Singer, F.D. Physiological and evolutionary ecology of acquiring nutrients and energy. In Ecology in Action; Singer, F.D., Ed.; Cambridge University Press: Cambridge, UK, 2016; pp. 86–112. [Google Scholar]
- Matsumoto, T. The role of termites in an equatorial rain forest ecosystem of west Malaysia: Population density, biomass, carbon, nitrogen and calorific content and respiration rate. Oecologia 1976, 22, 153–178. [Google Scholar] [CrossRef] [PubMed]
- La Fage, J.P.; Nutting, W.L. Nutrient dynamics of termites. In Production Ecology of Ants and Termites; Brian, M.V., Ed.; Cambridge University Press: Cambridge, UK, 1978; pp. 165–232. [Google Scholar]
- Prestwich, G.D.; Bentley, B.L.; Carpenter, E.J. Nitrogen sources for Neotropical nasute termites: Fixation and selective foraging. Oecologia 1980, 46, 397–401. [Google Scholar] [CrossRef]
- Miura, T.; Matsumoto, T. Diet and nest material of the processional termite Hospitalitermes, and cohabitation of Termes (Isoptera, Termitidae) on Borneo Island. Insectes Soc. 1997, 44, 267–275. [Google Scholar] [CrossRef]
- Spears, B.M.; Veckert, D.N. Survival and food consumption by the desert termite Gnathamitermes tubiformans in relation to dietary nitrogen source and levels. Environ. Entomol. 1979, 5, 1022–1025. [Google Scholar] [CrossRef]
- Sands, W.A. The association of termites and fungi. In Biology of Termites; Krishna, K., Weesner, F.M., Eds.; Academic Press: London, UK, 1969; pp. 495–519. [Google Scholar]
- Duryea, M.L.; Huffman, J.B.; Osbrink, W. Will subterranean termites consume landscape mulches? J. Arboric. 1999, 25, 143–150. [Google Scholar]
- Souza, J.H.D.; Menezes, E.D.L.A.; Mauri, R.; Menezes, E.B. Suscetibilidade de cinco essencias florestais a Coptotermes gestroi. Rev. Árvore 2009, 33, 1043–1050. [Google Scholar] [CrossRef]
- Kirker, G.; Blodgett, A.; Arango, R.; Lebow, P.; Clausen, C. The role of extractives in naturally durable wood species. Int. Biodeterior. Biodegrad. 2013, 82, 53–58. [Google Scholar] [CrossRef]
- Owoyemi, J.M.; Olaniran, O.S. Natural resistance of ten selected Nigerian wood species to subterranean termites attack. Int. J. Biol. Sci. 2014, 1, 35–39. [Google Scholar]
- Sattar, A.; Naeem, M.; Ehsan-Ul-Haq, A. Potential phagostimulants for the subterranean termite, Microtermes obesi (Blattodea: Termitidae). J. Biodivers. Bioprospect. Dev. 2015. [Google Scholar] [CrossRef]
- Dussutour, A.; Simpson, S.J. Carbohydrate regulation in relation to colony growth in ants. J. Exp. Biol. 2008, 211, 2224–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, A. Applying optimal foraging theory to assess nutrient availability ratios for ants. Ecology 2002, 83, 1935–1944. [Google Scholar] [CrossRef]
- Nonacs, P. Less growth with more food: How insect-prey availability changes colony demographics in the ant, Camponotus floridanus. J. Insect Physiol. 1991, 37, 891–898. [Google Scholar] [CrossRef]
- Grover, C.D.; Kay, A.D.; Monson, J.A.; Marsh, T.C.; Holway, D.A. Linking nutrition and behavioural dominance: Carbohydrate scarcity limits aggression and activity in Argentine ants. Proc. R. Soc. Lond. 2007, 274, 2951–2957. [Google Scholar] [CrossRef]
- Collins, N.M. Observations on the foraging activity of Hospitalitermes umbrinus (Haviland), (Isoptera: Termitidae) in the Gunong Mulu National Park, Sarawak. Ecol. Entomol. 1979, 4, 231–238. [Google Scholar] [CrossRef]
- Miura, T.; Matsumoto, T. Open-Air Litter Foraging in the Nasute Termite Longipeditermes longipes (Isoptera: Termitidae). J. Insect Behav. 1998, 11, 179–189. [Google Scholar] [CrossRef]
- Martius, C.; Amelung, W.; Garcia, M.V.B. The Amazonian forest termite (Isoptera: Termitidae) (Constrictotermes cavifrons) feeds on microepiphytes. Sociobiology 2000, 35, 379–383. [Google Scholar]
- Roisin, Y.; Pasteels, J.M. The Nasute Termites (Isoptera: Nasutitermitinae) of Papua New Guinea. Invertebr. Taxon. 1996, 10, 507–616. [Google Scholar] [CrossRef]
- Barbosa-Silva, A.M.; Silva, A.C.; Pereira, E.C.G.; Buril, M.L.L.; Silva, N.H.; Cáceres, M.E.S.; Aptroot, A.; Bezerra-Gusmão, M.A. Lichen Richness Consumed by Constrictotermes cyphergaster in Semi-arid Region of Brazil. Sociobiology 2018, in press. [Google Scholar]
- Nash, T.H., III. Lichen Biology; Cambridge University Press: Cambridge, UK, 2008; 486p. [Google Scholar]
- Vasconcellos, A.; Araújo, V.F.P.; Moura, F.M.S.; Bandeira, A.G. Biomass and population structure of Constrictotermes cyphergaster (Silvestri) (Isoptera: Termitidae) in the dry forest of Caatinga, Northeastern Brazil. Neotrop. Entomol. 2007, 36, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Moura, F.M.S.; Vasconcellos, A.; Araújo, V.F.P.; Bandeira, A.G. Consumption of Vegetal Organic Matter by Constrictotermes cyphergaster (Isoptera, Termitidae, Nasutitermitinae) in an Area of Caatinga, Northeastern Brazil. Sociobiology 2008, 51, 181–189. [Google Scholar]
- Moura, F.M.S.; Vasconcellos, A.; Araújo, V.F.P.; Bandeira, A.G. Feeding Habit of Constrictotermes cyphergaster (Isoptera, Termitidae) in an Area of Caatinga, Northeast. Brazil. Sociobiology 2006, 48, 1–6. [Google Scholar]
- Simpson, S.J.; Raubenheimer, D. The Nature of Nutrition: A Unifying Framework from Animal Adaptation to Human Obesity; Princeton University Press: Princeton, NJ, USA, 2012; p. 239. [Google Scholar]
- Governo do estado da Paraíba. Atlas Geográfico da Paraíba; Grafset: João Pessoa, Brazil, 1985; 100p.
- Núcleo de Meteorologia Aplicada. Atlas Climatológico da Paraíba; Universidade Federal da Paraíba: Campina Grande, Brazil, 1987; 143p. [Google Scholar]
- Cáceres, M.E.S. Corticolous, Crustose and Microfoliose Lichens of Northeastern Brazil; Libri Botanici: Berchtesgaden, Germany, 2007; 168p. [Google Scholar]
- Lücking, R.; Rivas-Plata, E. Clave y Guía Ilustrada Para Géneros de Graphidaceae. Glalia 2008, 1, 1–39. [Google Scholar]
- Marbach, B. Corticole und Lignicole Arten der Flechtengattung Buellia Sensu Lato in den Subtropen und Tropen; Bibliotheca Lichenologica: Berlin, Stuttgart, 2000; 384p. [Google Scholar]
- Nylander, W. Circa novum in studio lichenum criterium chemicum. Flora 1866, 49, 198–201. [Google Scholar]
- Allison, L.E. Organic carbon. In Methods of Soil Analysis, Chemical and Microbiological Properties; Black, C.A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1367–1378. [Google Scholar]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis; Black, C.A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1149–1178. [Google Scholar]
- Tedesco, J.M.; Gianello, C.; Bissani, C.A.; Bohnen, H.; Volkweiss, S.J. Análises de Solo, Plantas e Outros Materiais; Boletim Técnico, Universidade Federal do Rio Grande do Sul: Porto Alegre, Brazil, 1995; 174p. [Google Scholar]
- Lenz, M.; Becker, G.; Garcia, M. Zur Eignung von verschiedenen Subtraten und zu ihrer Auswahl fur die Prufung mit Rhinotermitiden. Mater. Org. 1976, 11, 121–144. [Google Scholar]
- Bustamante, N.C.R. Preferências Alimentares de 5 Espécies de Cupins Nasutitermes Dudley, 1890 (Termitidae, Isoptera) por 7 Espécies de Madeira de Várzea na Amazônia Central. Master’s Thesis, Universidade Federal do Amazonas, INPA, Manaus, Brazil, 1993. [Google Scholar]
- Vasconcellos, A.; Bandeira, A.G. Avaliação do consumo de madeira por espécies de Nasutitermes e Microcerotermes (Insecta, Isoptera, Termitidae). Rev. Nordestina Biol. 2000, 14, 17–24. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Cáceres, M.E.S.; Lucking, R.; Rambold, G. Corticolous microlichens in Northeastern Brazil: Habitat differentiation between coastal Mata Atlântica, Caatinga and Brejos de Altitude. Bryologist 2008, 111, 98–117. [Google Scholar] [CrossRef]
- Silva, A.C. Dinâmica do Forrageio de Constrictotermes cyphergaster (Blattaria, Isoptera) em um Ecossistema Semiárido do Nordeste Brasileiro. Master’s Thesis, Universidade Federal da Paraíba (UFPB), Paraiba, Brazil, 2017; 70p. [Google Scholar]
- Machida, M.; Kitade, O.; Miura, T.; Matsumoto, T. Nitrogen recycling through proctodeal trophallaxis in the Japanese damp-wood termite Hodotermopsis japonica (Isoptera, Termopsidae). Insectes soc. 2001, 48, 52–56. [Google Scholar] [CrossRef]
- Yamada, A.; Inoque, T.; Noda, S.; Hongoh, Y.; Ohkuma, M. Evolutionary trend phylogenetic diversity of nitrogen fixation genes in the gut community of wood-feeding termites. Mol. Ecol. 2007, 16, 3768–3777. [Google Scholar] [CrossRef] [PubMed]
- Dhanarajan, G. Cannibalism and necrophagy in a subterranean termite (Reticulitermes lucifagus var. santonensis). Malay. Nat. J. 1978, 31, 237–251. [Google Scholar]
- Potrikus, C.J.; Breznak, J.A. Uric Acid-Degrading Bacteria in Guts of Termites Reticulitermes flavipes (Kollar). Appl. Environ. Microbiol. 1980, 40, 117–124. [Google Scholar] [PubMed]
- Potrikus, C.J.; Breznak, J.A. Gut bacteria recycle uric acid nitrogen in termites: A strategy for nutrient conservation. Proc. Natl. Acad. Sci. USA 1981, 78, 4601–4605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002; 464p. [Google Scholar]
- Rückamp, D.; Amelung, W.; Theisz, N.; Bandeira, A.G.; Martius, C. Phosphorus forms in Brazilian termite nests and soils: Relevance of feeding guild and ecosystems. Geoderma 2010, 155, 269–279. [Google Scholar] [CrossRef]
- Noirot, C.; Darlington, J.P.E.C. Termites nests: Architecture, regulation and defence. In Termites: Evolution, Sociality, Symbiosis, Ecology; Abe, T., Higashi, M., Bignell, D.E., Eds.; Kluwer Academic Publications: Dordrecht, The Netherlands, 2000; pp. 121–139. [Google Scholar]
- Janzow, M.P.; Judd, T.M. The Termite Reticulitermes flavipes (Rhinotermitidae: Isoptera) Can Acquire Micronutrients from Soil. Environ. Entomol. 2015, 44, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Buxton, R.D. Changes in the composition and activities of termite communities in relation to changing rainfall. Oecologia 1981, 51, 371–378. [Google Scholar] [CrossRef]
- Lepage, M.; Darlington, J.P.E.C. Population dynamics of termites. In Termites: Evolution, Sociality, Symbiosis, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Academic Plublications: Dordrecht, The Netherlands, 2000; pp. 333–362. [Google Scholar]
- Judd, T.M. The Effects of Water, Season, and Colony Composition on Foraging Preferences of Pheidole ceres [Hymenoptera: Formicidae]. J. Insect Behav. 2005, 18, 781–803. [Google Scholar] [CrossRef]
- Stein, M.B.; Thorvilson, H.G.; Johnson, J.W. Seasonal changes in bait preference by red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae). Fla. Entomol. 1990, 73, 117–123. [Google Scholar] [CrossRef]
- Wallace, B.A.; Judd, T.M. A Test of Seasonal Responses to Sugars in Four Populations of the Termite Reticulitermes flavipes. J. Econ. Entomol. 2010, 103, 2126–2131. [Google Scholar] [CrossRef] [PubMed]
- Cornelius, M.L.; Osbrink, W.L.A. Effect of Seasonal Changes in Soil Temperature and Moisture on Wood Consumption and Foraging Activity of Formosan Subterranean Termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2011, 104, 1024–1030. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Lichen | ¹ Mean ± SE | Range |
|---|---|---|
| Dirinaria confluens | 0.0106 ± 0.0009 a | 0.005–0.015 |
| Pertusaria flavens | 0.0064 ± 0.0007 b | 0.003–0.012 |
| Haematomma persoonii | 0.0067 ± 0.0007 b | 0.003–0.012 |
| Lecanora spp. | 0.0081 ± 0.0004 ab | 0.005–0.012 |
| Total mean consumption | 0.0318 ± 0.0009 | 0.003–0.015 |
| df | Deviance | Resid. df | Resid. dev | F | Pseudo R2 | P (>F) | |
|---|---|---|---|---|---|---|---|
| N | 1 | 0.0050547 | 48 | 0.010547 | 22.6322 | 0.32 | <0.001 |
| C | 1 | 0.0001990 | 47 | 0.010348 | 0.8908 | 0.01 | 0.35 |
| P | 1 | 0.000746 | 46 | 0.010274 | 0.5661 | 0.00 | 0.56 |
| CN | 1 | 0.0036748 | 48 | 0.11927 | 16.4539 | 0.23 | <0.001 |
| C:P | 1 | 0.0009845 | 47 | 0.010943 | 4.4080 | 0.06 | <0.01 |
| N:P | 1 | 0.0006689 | 46 | 0.010274 | 2.9951 | 0.04 | 0.09 |
| C:N:P | 1 | 0.0001860 | 46 | 0.011741 | 0.7444 | 0.00 | 0.39 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbosa-Silva, A.M.; Vasconcellos, A. Consumption Rate of Lichens by Constrictotermes cyphergaster (Isoptera): Effects of C, N, and P Contents and Ratios. Insects 2019, 10, 23. https://doi.org/10.3390/insects10010023
Barbosa-Silva AM, Vasconcellos A. Consumption Rate of Lichens by Constrictotermes cyphergaster (Isoptera): Effects of C, N, and P Contents and Ratios. Insects. 2019; 10(1):23. https://doi.org/10.3390/insects10010023
Chicago/Turabian StyleBarbosa-Silva, Ana M., and Alexandre Vasconcellos. 2019. "Consumption Rate of Lichens by Constrictotermes cyphergaster (Isoptera): Effects of C, N, and P Contents and Ratios" Insects 10, no. 1: 23. https://doi.org/10.3390/insects10010023