Termite Communities along A Disturbance Gradient in a West African Savanna

Abstract
:1. Introduction
2. Materials and Methods
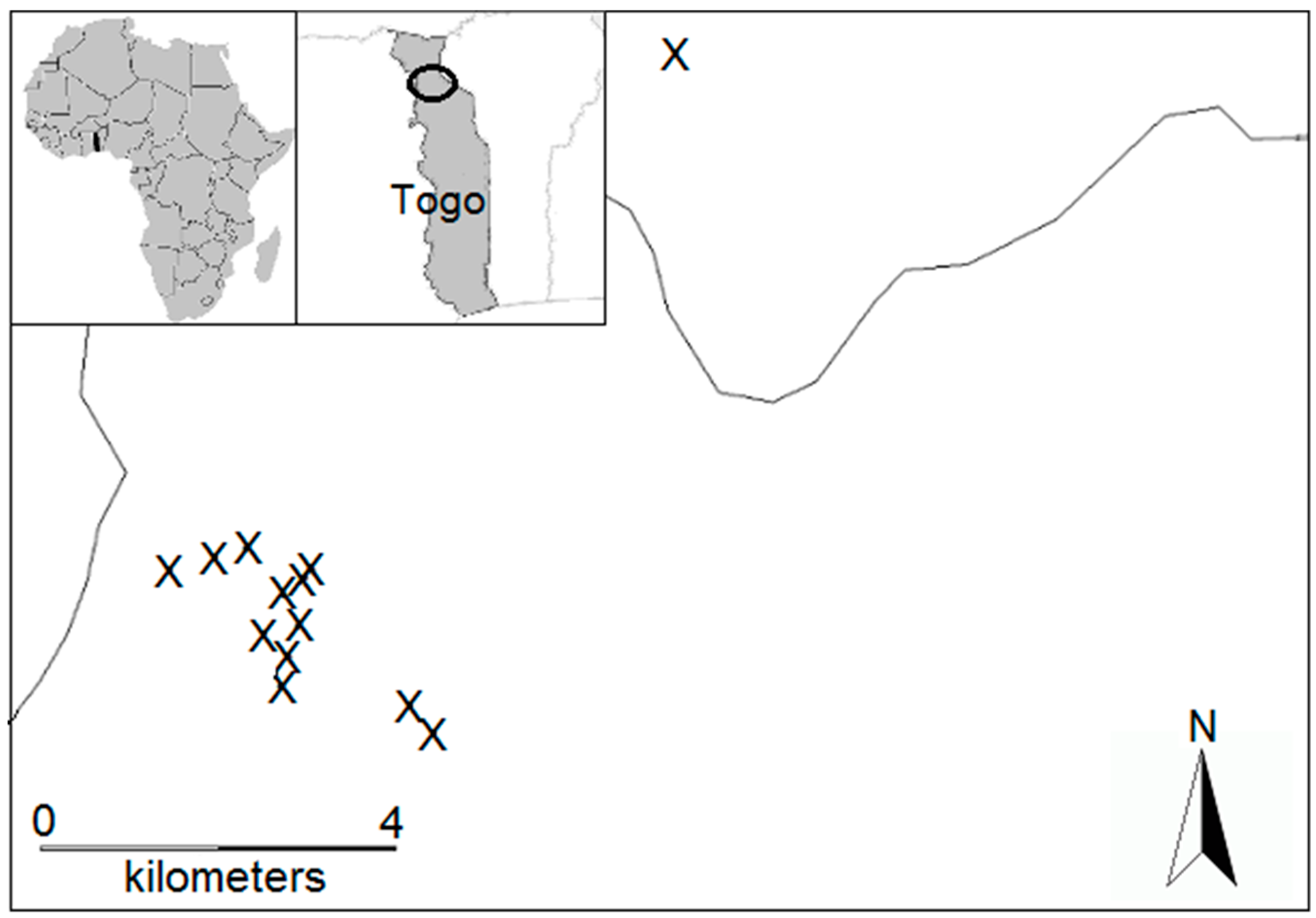
2.1. Termite Sampling
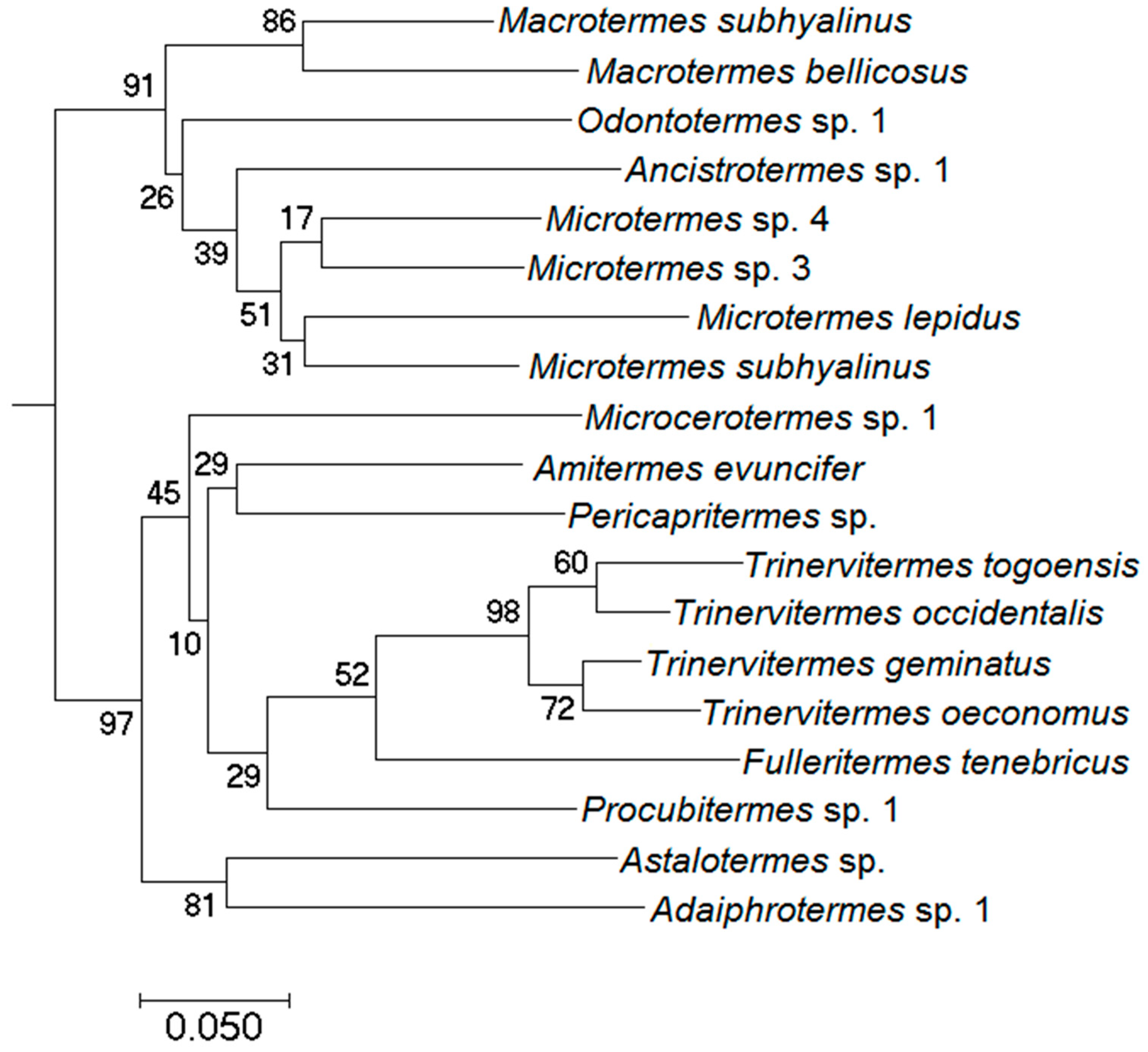
2.2. Genetic Identification and Phylogenetic Analyses
2.3. Phylogenetic Community Structure Analyses
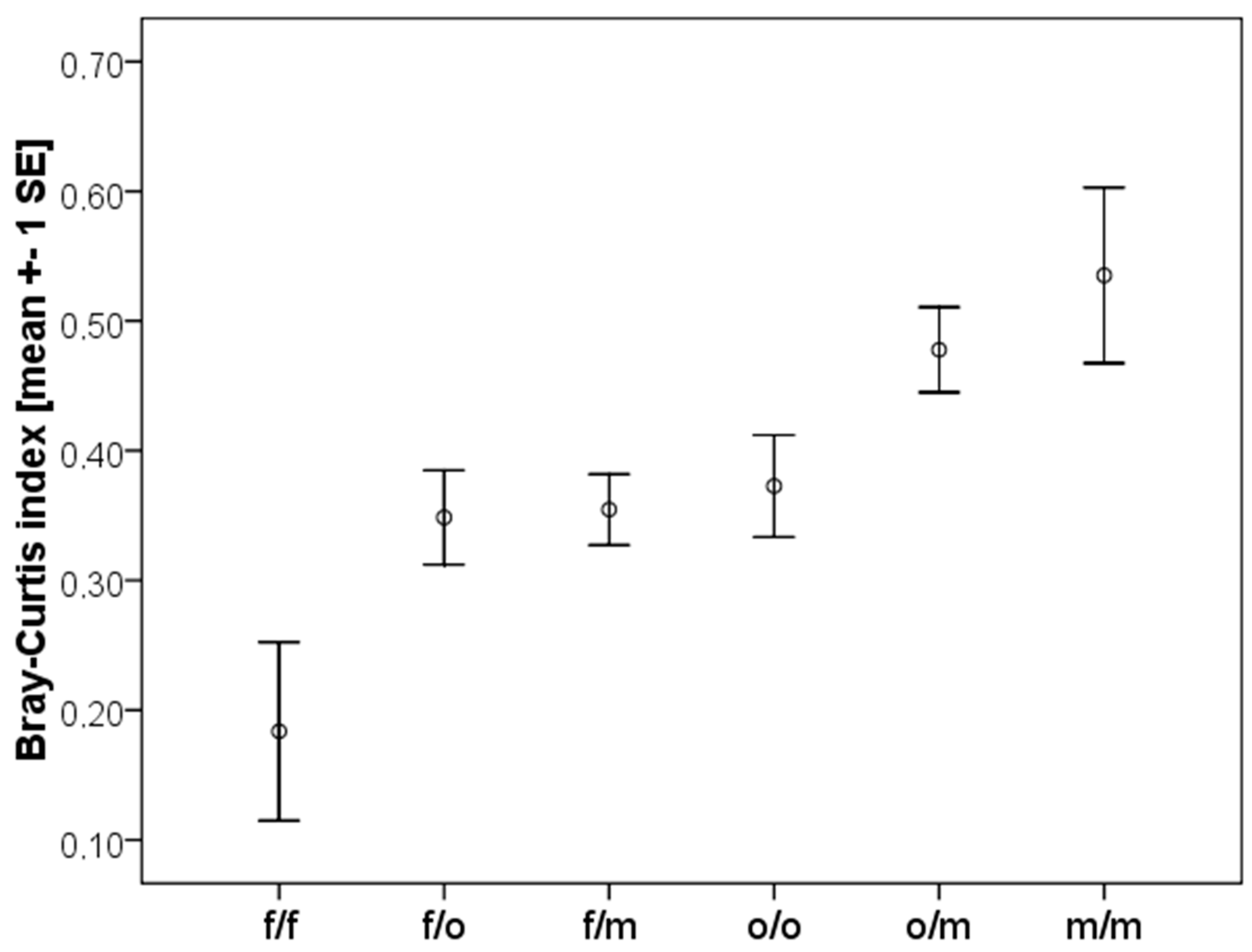
2.4. Similarity between Fallows
2.5. Other Statistical Analyses
3. Results
3.1. Diversity
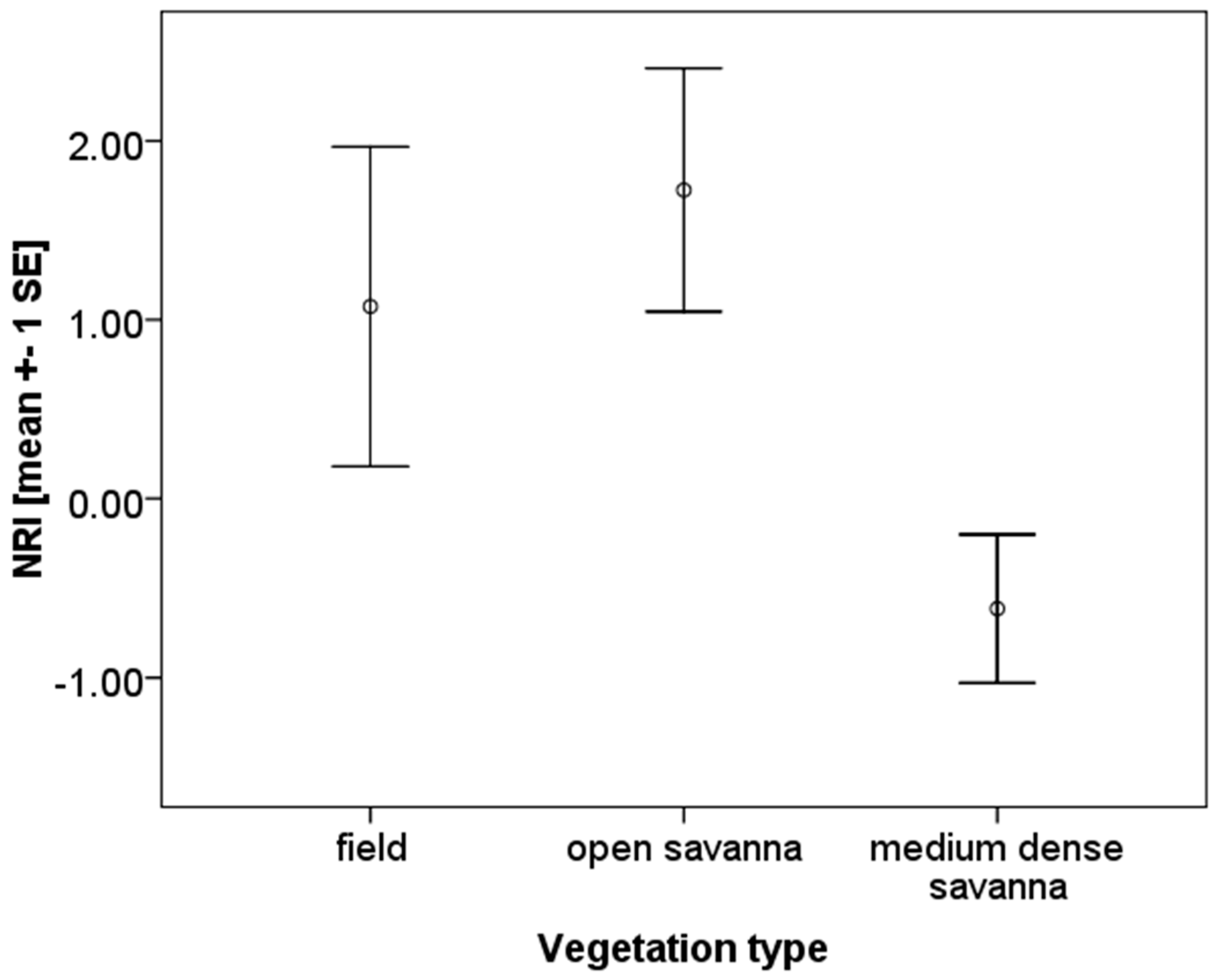
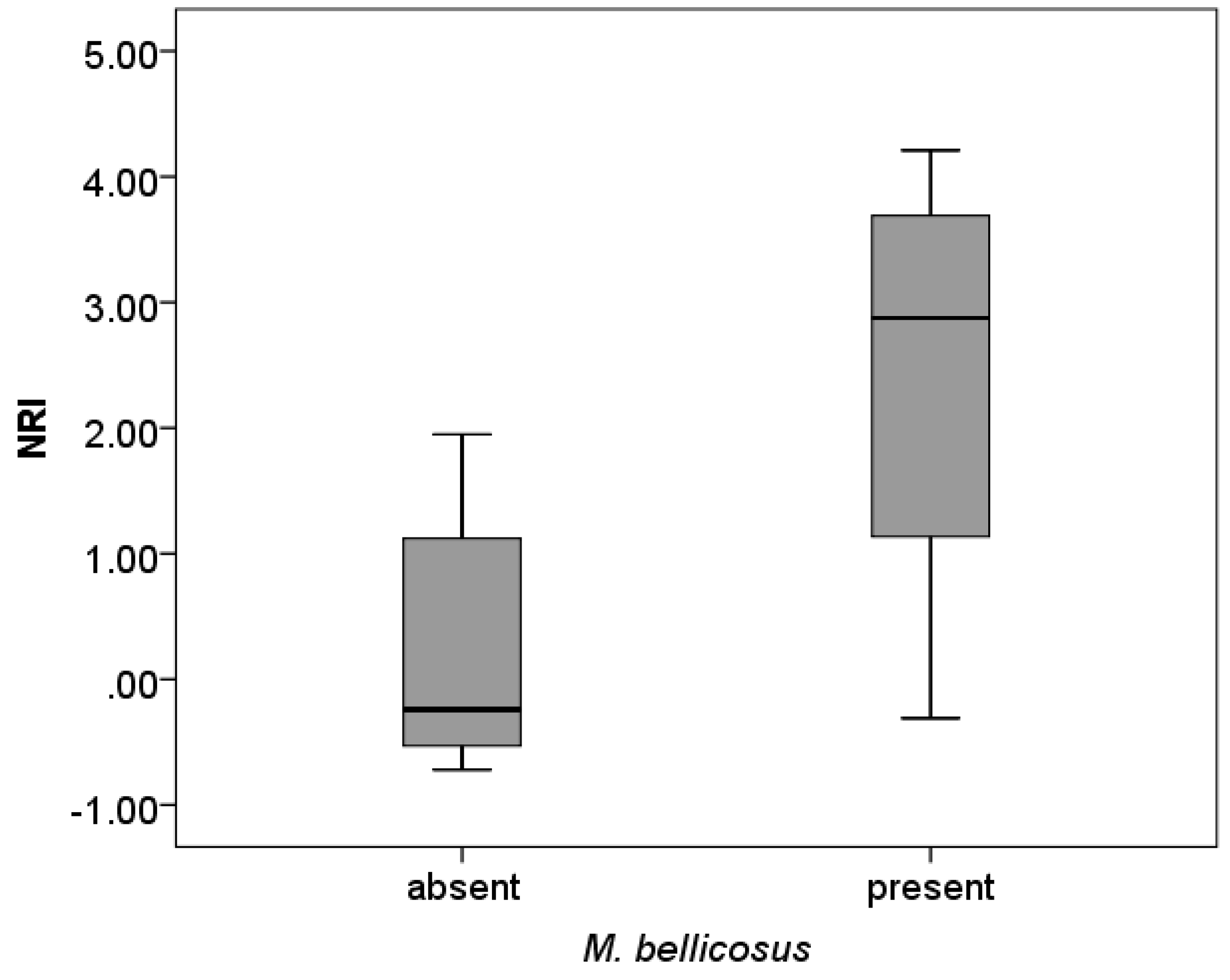
3.2. Phylogenetic Community Structure
3.3. Similarity between Fallows
3.4. Pest Species
4. Discussion
Comparison with Other Community Studies in West Africa
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pringle, R.M.; Doak, D.F.; Brody, A.K.; Jocque, R.; Palmer, T.M. Spatial pattern enhances ecosystem functioning in an African Savanna. PLoS Biol. 2010, 8, e1000377. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.A.; Dawes, T.Z.; Ward, P.R.; Lo, N. Ants and termites increase crop yield in a dry climate. Nat. Commun. 2011, 2, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouland-Lefèvre, C. Termites as pests of agriculture. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2011; pp. 499–517. [Google Scholar]
- Eggleton, P.; Tayasu, I. Feeding groups, lifetypes and the global ecology of termites. Ecol. Res. 2001, 16, 941–960. [Google Scholar] [CrossRef]
- Davies, R.G.; Eggleton, P.; Jones, D.T.; Gathorne-Hardy, F.J.; Hernandez, L.M. Evolution of termite functional diversity: Analysis and synthesis of local ecological and regional influences on local species richness. J. Biogeogr. 2003, 30, 847–877. [Google Scholar] [CrossRef]
- Eggleton, P. An introduction to termites: Biology, taxonomy and functional morphology. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2011; pp. 1–26. [Google Scholar]
- Dosso, K.; Konaté, S.; Aidara, D.; Linsenmair, K.E. Termite diversity and abundance across fire-induced habitat variability in a tropical moist savanna (Lamto, Central Cˆote d’Ivoire). J. Trop. Ecol. 2010, 26, 323–334. [Google Scholar] [CrossRef]
- Dosso, K.; Deligne, J.; Yeó, K.; Konaté, S.; Linsenmair, K.E. Changes in the termite assemblage across a sequence of land-use systems in the rural area around Lamto Reserve in central Côte d’Ivoire. J. Insect Conserv. 2013, 17, 1047–1057. [Google Scholar] [CrossRef]
- Hausberger, B.; Kimpel, D.; van Neer, A.; Korb, J. Uncovering cryptic species diversity of a community in a West African Savanna. Mol. Phylogenet. Evol. 2011, 61, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Hausberger, B.; Korb, J. A phylogenetic community approach for studying termite communities in a West African Savannah. Biol. Lett. 2015. [Google Scholar] [CrossRef] [PubMed]
- Hausberger, B.; Korb, J. The impact of anthropogenic disturbance on assembly patterns of termite communities. Biotropica 2016, 48, 356–364. [Google Scholar] [CrossRef]
- Nobre, T.; Rouland-Lefèvre, C.; Aanen, D.K. Comparative biology of fungus cultivation in termites and ants. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2011; pp. 193–210. [Google Scholar]
- Basu, P. Food preference and interspecific interactions in fungus-growing termite assemblage in a West African Savanna. Curr. Sci. 2011, 100, 77–83. [Google Scholar]
- Korb, J.; Linsenmair, K.E. Resource availability and distribution patterns, indicators of competition between Macrotermes bellicosus and other macro-detritivores in the Comoé National Park, Côte d’Ivoire. Afr. J. Ecol. 2001, 39, 257–265. [Google Scholar] [CrossRef]
- Jones, D.T.; Eggleton, P. Sampling termite assemblages in tropical forests: Testing a rapid biodiversity assessment protocal. J. Appl. Ecol. 2000, 37, 191–203. [Google Scholar] [CrossRef]
- Korb, J.; Linsenmair, K.E. The causes of spatial patterning of mounds of a fungus-cultivating termite: Results from nearest-neighbour analysis and ecological studies. Oecologia 2001, 127, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Vamosi, S.M.; Heard, S.B.; Vamosi, J.C.; Webb, C.O. Emerging patterns in the comparative analysis of phylogenetic community structure. Mol. Ecol. 2009, 18, 572–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, G.C. Keys to the Genera of the African Termites; Ibadan University Press: Ibadan, Nigeria, 1965. [Google Scholar]
- Sands, W.A. A revision of the termite subfamily Nasutitermitinae (Isoptera, Termitidae) from the Ethiopian region. Bull. Br. Museum (Nat. Hist.) Entomol. 1965, 4, 3–172. [Google Scholar]
- Donovan, S.E.; Eggleton, P.; Bignell, D.E. Gut content analysis and a new feeding group classification of termites. Ecol. Entomol. 2001, 26, 356–366. [Google Scholar] [CrossRef]
- Davies, R.G. Feeding group responses of a Neotropical termite assemblage to rain forest fragmentation. Oecologia 2002, 133, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Legendre, F.; Whiting, M.F.; Bordereau, C.; Cancello, E.M.; Evans, T.A.; Grandcolas, P. The phylogeny of termites (Dictyoptera: Isoptera) based on mitochondrial and nuclear markers: Implications for the evolution of the worker and pseudergate castes, and foraging behaviors. Mol. Phylogenet. Evol. 2008, 48, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.O.; Ackerly, D.D.; Kembel, S.W. Phylocom: Sofware for the analysis of phylogenetic community structure and character evolution. Bioinformatics 2008, 24, 2098–2100. [Google Scholar] [CrossRef]
- Chromas, version 2.4.4; Technilysium Pty Ltd.: South Brisbane, Australia, 1998–2016.
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Entsminger, G.L. Swap algorithms in null model analysis. Ecology 2003, 84, 532–535. [Google Scholar] [CrossRef]
- Hardy, O.J. Testing the spatial phylogenetic structure of local communities: Statistical performances of different null models and test statistics on a locally neutral community. J. Ecol. 2008, 96, 914–926. [Google Scholar] [CrossRef]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magurran, A.E. Ecological Diversity and Its Measurement; Springer: Dordrecht, The Netherlands, 1988. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9. User’s Guide and Application. 2013. Available online: http://purl.oclc.org/estimates (accessed on 1 May 2012).
- Wood, T.G.; Smith, R.W.; Johnson, R.A.; Komolafe, P.O. Termite damage and crop loss studies in Nigeria—Pre-harvest losses to yams due to termites and other soil pests. Trop. Pest Manag. 1980, 26, 355–370. [Google Scholar] [CrossRef]
- Cowie, R.H.; Logan, J.W.M.; Wood, T.G. Termite (Isoptera) damage and control in tropical forestry with special reference to Africa and Indo-Malaysia: A review. Bull. Entomol. Res. 1989, 79, 173–184. [Google Scholar] [CrossRef]
- Collins, N.M. Termite damage and crop loss studies in Nigeria—Assessment of damage to upland sugarcane. Trop. Pest Manag. 1984, 30, 26–28. [Google Scholar] [CrossRef]
- Majer, J.D.; Heterick, B.; Gohr, T.; Hughes, E.; Mounsher, L.; Grigg, A. Is thirty-seven years sufficient for full return of the ant biota following restoration? Ecol. Process. 2013, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Sands, W.A. Termite distribution in man-modified habitats in West Africa, with special reference to species segregation in the genus Trinervitermes (Isoptera, Termitidae, Nasutitermitinae). J. Anim. Ecol. 1965, 34, 557–571. [Google Scholar] [CrossRef]
- Sands, W.A. Foraging behaviour and feeding habits in five species of Trinervitermes in West Africa. Entomol. Exp. Appl. 1961, 4, 277–288. [Google Scholar] [CrossRef]
- Joseph, G.S.; Seymour, C.L.; Cumming, G.S.; Cumming, D.H.M.; Mahlangu, Z. Termite mounds as islands: Woody plant assemblages relative to termitarium size and soil properties. J. Veg. Sci. 2013, 24, 702–711. [Google Scholar] [CrossRef]
- Eggleton, P.; Bignell, D.E.; Hauser, S.; Dibog, L.; Norgrove, L.; Madong, B. Termite diversity across an anthopogenic disturbance gradient in the humid forest zone of West Africa. Agric. Ecosyst. Environ. 2002, 90, 189–202. [Google Scholar] [CrossRef]
- Su, N.Y.; Scheffrahn, R.H. Intraspecific and interspecific competition of the Formosan and the Eastern subterranean termite—Evidence from field observations (Isoptera, Rhinotermitidae). Sociobiology 1988, 14, 157–164. [Google Scholar]
- Leponce, M.; Roisin, Y.; Pasteels, J.M. Intraspecific interactions in a community of arboreal nesting termites. (Isoptera: Termitidae). J. Insect Behav. 1996, 9, 799–817. [Google Scholar] [CrossRef]
- Bourguignon, T.; Sobotnik, J.; Lepoint, G.; Martin, J.M.; Roisin, Y. Niche differentiation among neotropical soldierless soil-feeding termites revealed by stable isotope ratios. Soil Biol. Biochem. 2009, 41, 2038–2043. [Google Scholar] [CrossRef]
- Bourguignon, T.; Leponce, M.; Roisin, Y. Beta-diversity of termite assemblages among primary French Guiana rain forests. Biotropica 2011, 43, 473–479. [Google Scholar] [CrossRef]
- Li, H.F.; Lan, Y.C.; Fujisaki, I.; Kanzaki, N.; Lee, H.J.; Su, N.Y. Termite assemblage pattern and niche partitioning in a tropical forest ecosystem. Environ. Entomol. 2015, 44, 546–556. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F | sf | fg | L | M | N | O | P | R | S | T | U | W | 2 | 3 | 5 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trinervitermes occidentalis | Te | N | IIg | 15 | 2 | 2 | 1 | 5 | 5 | 3 | 8 | 9 | 0 | 1 | 0 | 0 |
| Trinervitermes geminatus | Te | N | IIg | 3 | 2 | 4 | 0 | 7 | 6 | 6 | 2 | 13 | 0 | 1 | 0 | 0 |
| Trinervitermes oeconomus | Te | N | IIg | 5 | 6 | 2 | 0 | 2 | 0 | 3 | 1 | 3 | 0 | 4 | 1 | 0 |
| Trinervitermes togoensis | Te | N | IIg | 8 | 2 | 6 | 0 | 6 | 12 | 13 | 0 | 2 | 0 | 1 | 2 | 0 |
| Fulleritermes tenebricus | Te | N | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 |
| Microtermes subhyalinus | Te | M | IIf | 0 | 2 | 1 | 4 | 2 | 3 | 0 | 0 | 0 | 3 | 5 | 0 | 2 |
| Microtermes lepidus | Te | M | IIf | 1 | 0 | 4 | 5 | 1 | 0 | 0 | 6 | 6 | 13 | 2 | 1 | 6 |
| Microtermes sp.3 | Te | M | IIf | 4 | 10 | 3 | 3 | 4 | 0 | 2 | 1 | 0 | 2 | 5 | 5 | 2 |
| Microtermes sp.4 | Te | M | IIf | 1 | 0 | 3 | 8 | 0 | 1 | 0 | 1 | 1 | 4 | 2 | 0 | 2 |
| Macrotermes bellicosus | Te | M | IIf | 0 | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 2 | 0 | 5 | 0 | 18 |
| Macrotermes subhyalinus | Te | M | IIf | 0 | 1 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| Ancistrotermes sp.1 | Te | M | IIf | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 | 7 | 19 | 28 | 10 | 16 |
| Odontotermes sp.1 | Te | M | IIf | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 9 | 9 | 3 | 0 | 0 | 1 |
| Astalotermes sp. | Te | Ap | III | 4 | 2 | 1 | 0 | 0 | 3 | 3 | 0 | 1 | 1 | 0 | 0 | 0 |
| Adaiphrotermes sp.1 | Te | Ap | III | 0 | 0 | 2 | 1 | 1 | 0 | 0 | 1 | 4 | 0 | 0 | 1 | 2 |
| Microcerotermes sp.1 | Te | T | II | 6 | 0 | 11 | 13 | 11 | 17 | 23 | 33 | 15 | 5 | 14 | 15 | 2 |
| Amitermes evuncifer | Te | T | II | 0 | 0 | 0 | 1 | 0 | 3 | 0 | 5 | 0 | 13 | 0 | 5 | 0 |
| Procubitermes sp.1 | Te | T | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| Pericapritermes sp. | Te | T | III | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Number of species | 9 | 10 | 12 | 10 | 9 | 9 | 7 | 11 | 13 | 9 | 13 | 9 | 10 | |||
| Number of encounters | 47 | 30 | 40 | 63 | 39 | 52 | 53 | 68 | 73 | 63 | 70 | 41 | 54 | |||
| Fallow age (years) | 8 | 0 | 6 | 2 | 4 | 10 | 12 | 0 | 10 | 2 | 1 | 1 | 10 | |||
| Vegetation type | 1 | 0 | 2 | 1 | 2 | 2 | 1 | 0 | 1 | 0 | 2 | 1 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schyra, J.; Korb, J. Termite Communities along A Disturbance Gradient in a West African Savanna. Insects 2019, 10, 17. https://doi.org/10.3390/insects10010017
Schyra J, Korb J. Termite Communities along A Disturbance Gradient in a West African Savanna. Insects. 2019; 10(1):17. https://doi.org/10.3390/insects10010017
Chicago/Turabian StyleSchyra, Janine, and Judith Korb. 2019. "Termite Communities along A Disturbance Gradient in a West African Savanna" Insects 10, no. 1: 17. https://doi.org/10.3390/insects10010017