

Transcription Factor ATF3 Participates in DeltaNp63-Mediated Proliferation of Corneal Epithelial Cells
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Construction of ATF3 Promoter Luciferase Vectors
2.3. Analysis of ATF3 Promoter Activity
2.4. Establishment of Expression Vectors and siRNAs
2.5. Transfection with Plasmids or siRNAs
2.6. Infection with ΔNp63 Adenoviral Vectors
2.7. Quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR) Assay
2.8. Western Blot Assays
2.9. Proliferation Assays
2.10. Microarray Analysis
2.11. Statistical Analysis
3. Results
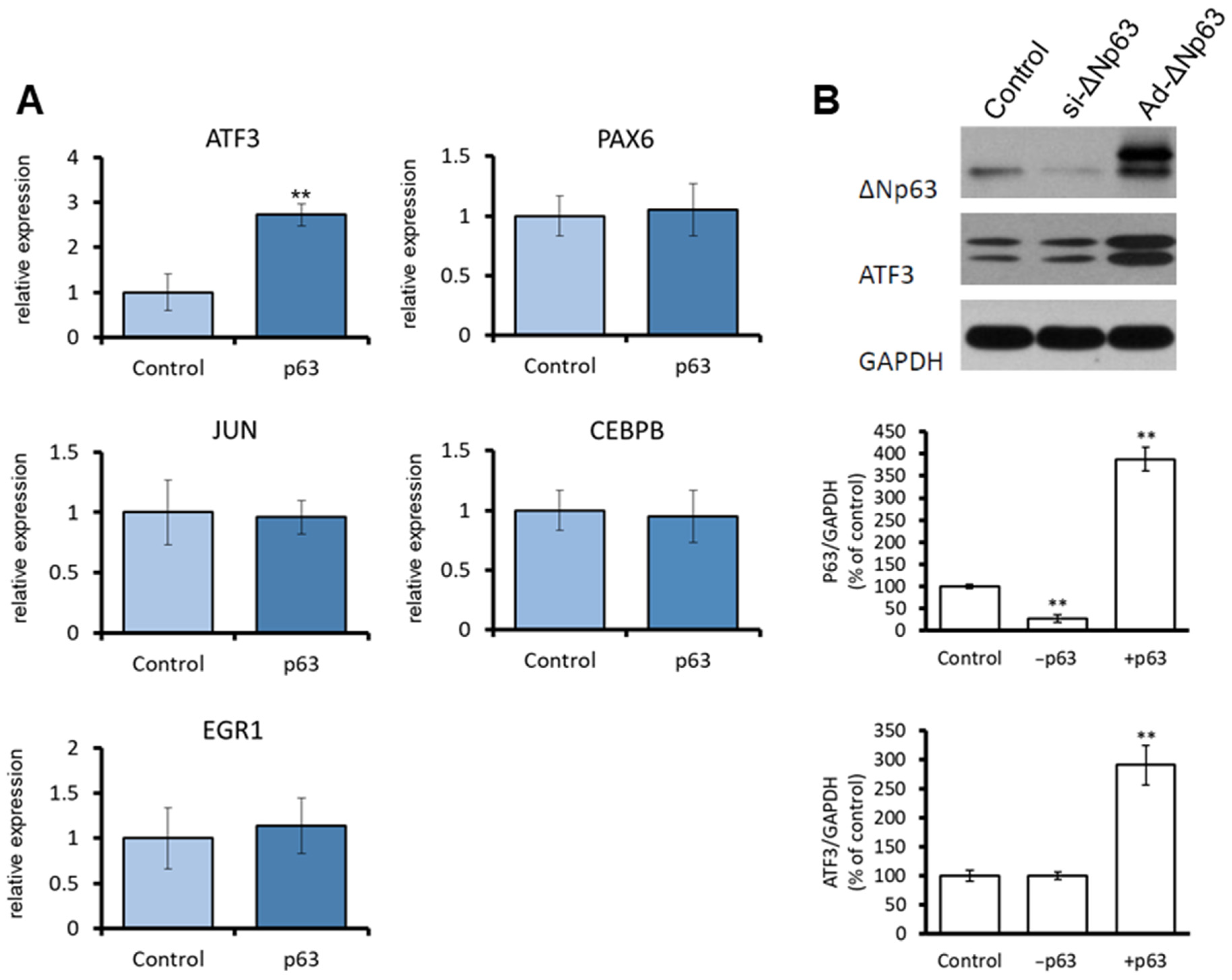
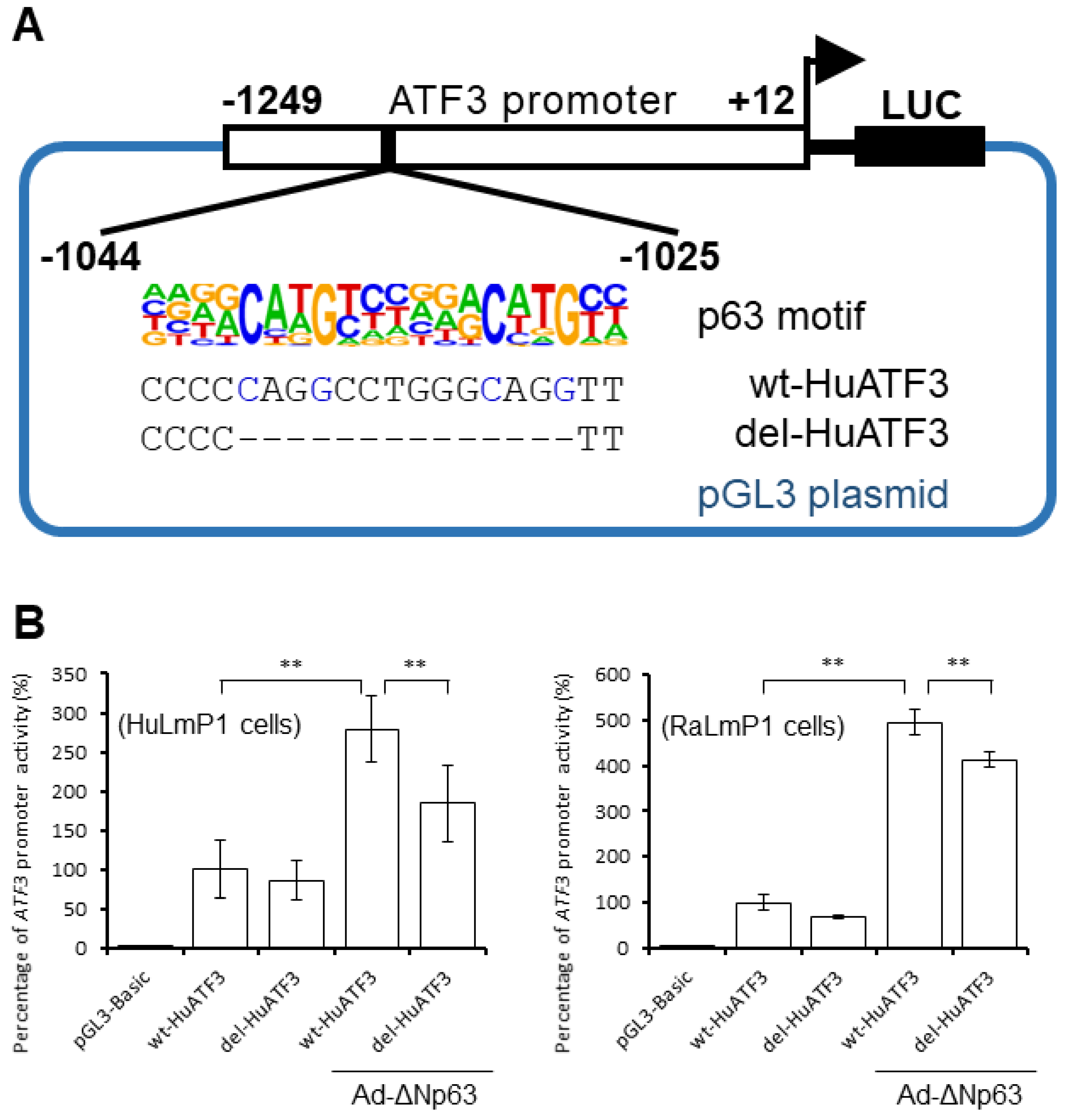
3.1. ΔNp63 Increases ATF3 Expression by Regulating ATF3 Promoter Activity in CECs
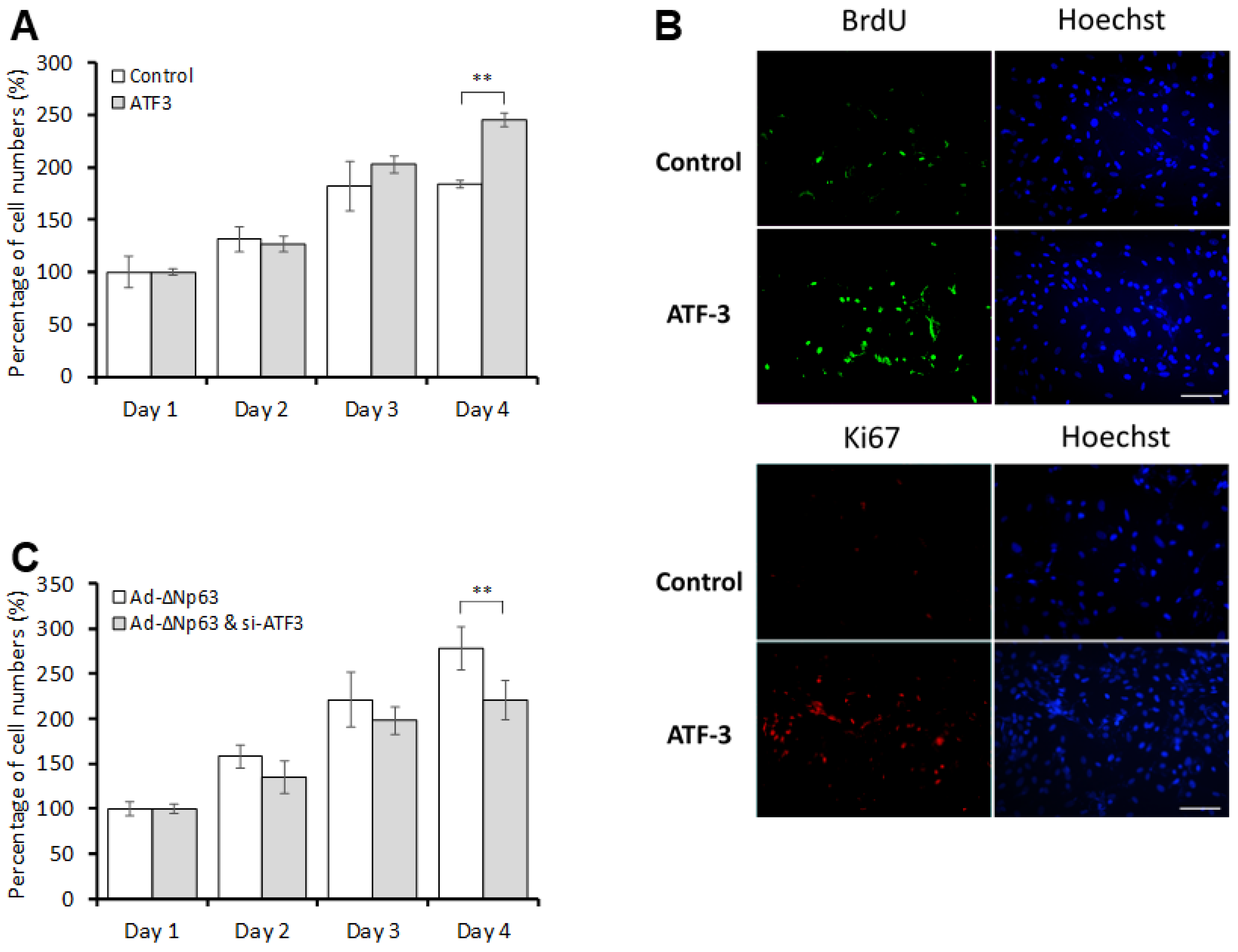
3.2. ΔNp63 Promotes CEC Proliferation in an ATF3-Dependent Manner
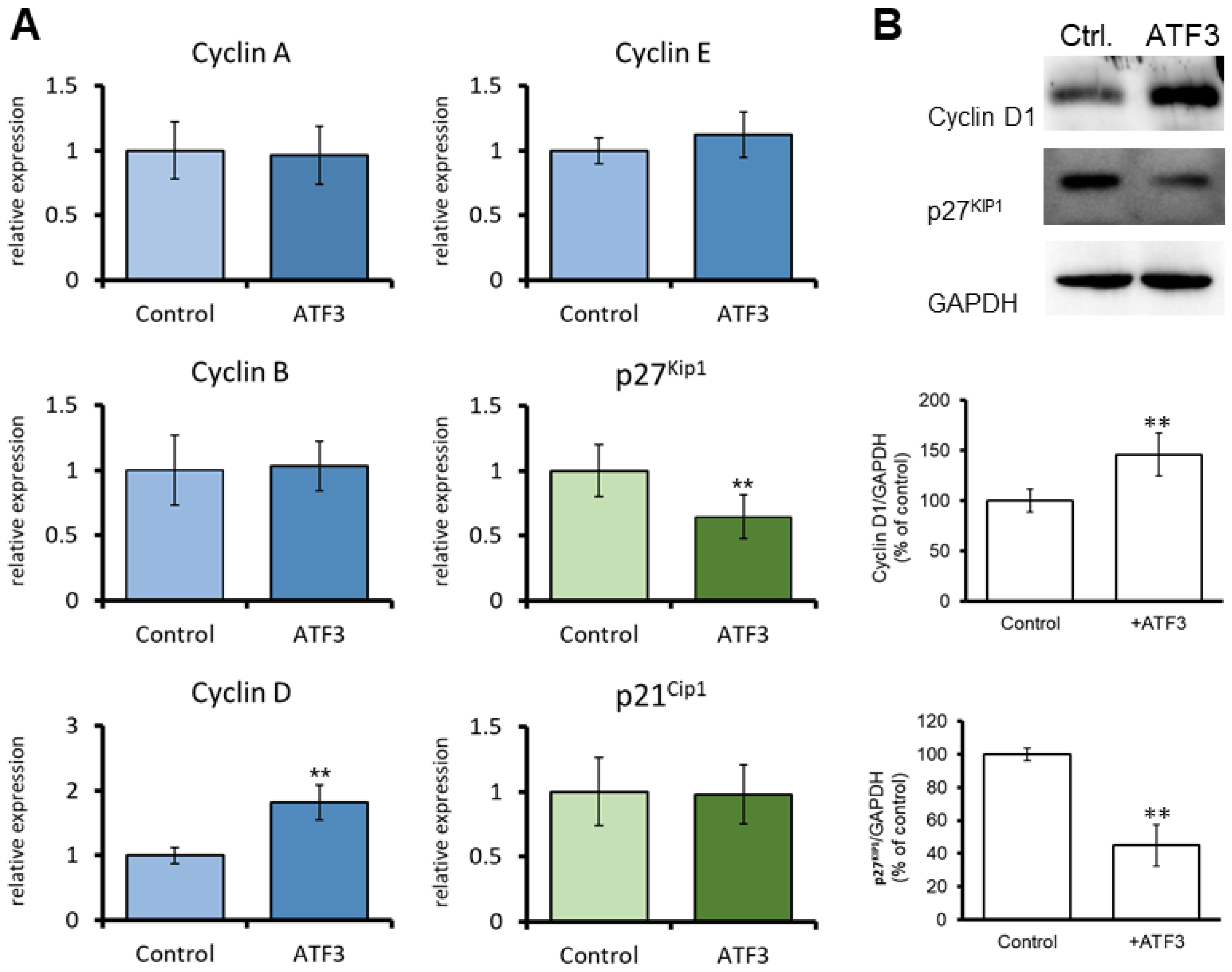
3.3. ATF3 Induces the Expression of Cyclin D in CECs
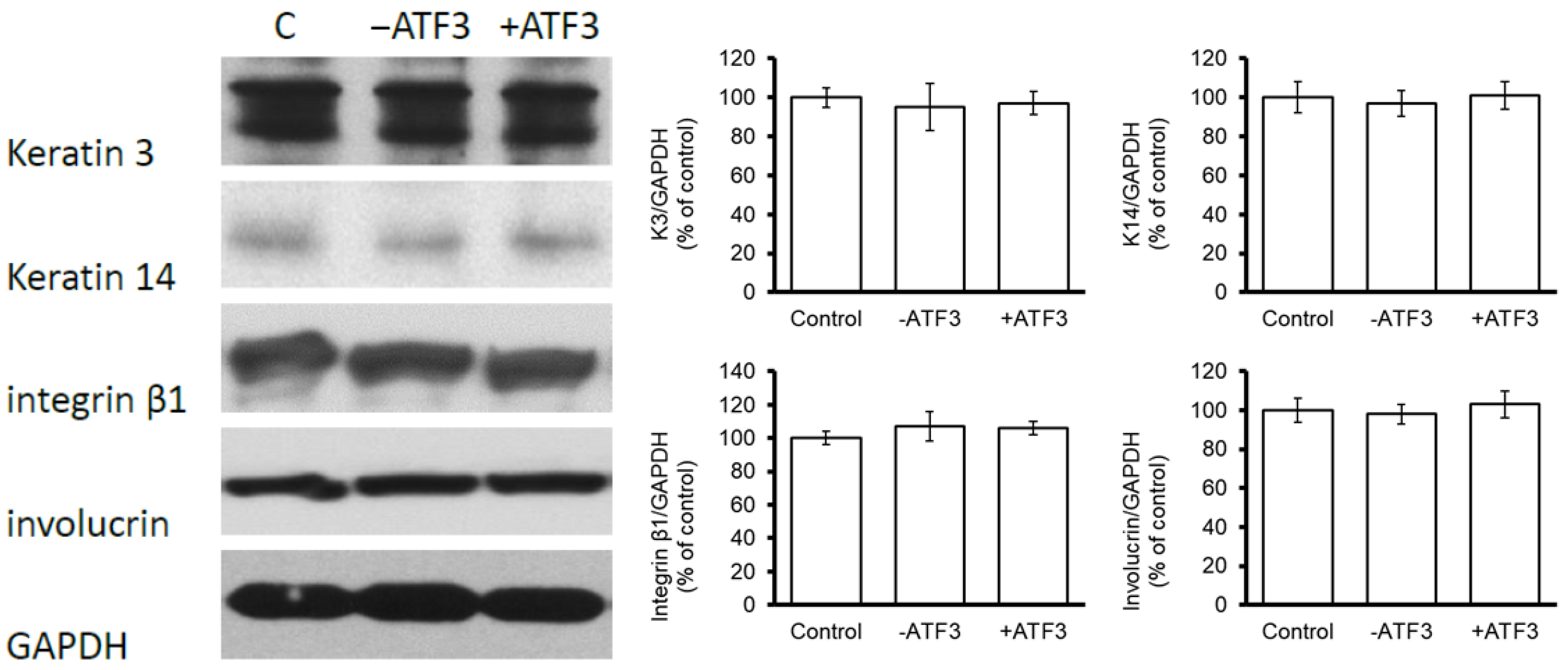
3.4. ATF3 Did Not Affect the Expression of Keratinocyte Differentiation-Related Proteins in CECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schlötzer-Schrehardt, U.; Kruse, F.E. Identification and characterization of limbal stem cells. Exp. Eye Res. 2005, 81, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Dua, H.S.; Azuara-Blanco, A. Limbal Stem Cells of the Corneal Epithelium. Surv. Ophthalmol. 2000, 44, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.; Zhou, H. Master regulatory role of p63 in epidermal development and disease. Cell. Mol. Life Sci. 2017, 75, 1179–1190. [Google Scholar] [CrossRef]
- Pellegrini, G.; Dellambra, E.; Golisano, O.; Martinelli, E.; Fantozzi, I.; Bondanza, S.; Ponzin, D.; McKeon, F.; De Luca, M. p63 identifies keratinocyte stem cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3156–3161. [Google Scholar] [CrossRef]
- Candi, E.; Dinsdale, D.; Rufini, A.; Salomoni, P.; Knight, R.A.; Mueller, M.; Krammer, P.H.; Melino, G. TAp63 and DeltaNp63 in cancer and epidermal development. Cell Cycle 2007, 6, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, Y.J.; Chen, H.C.; Chu, W.K.; Cheng, C.C.; Kuo, P.C.; Lin, L.Y.; Ma, H.K.; Chen, J.K. STAT3 regulates the proliferation and differentiation of rabbit limbal epithelial cells via a DeltaNp63-dependent mechanism. Investig. Ophthalmol. Vis. Sci. 2011, 52, 4685–4693. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, Y.J.; Kuo, P.C.; Chen, J.K. Transcriptional regulators of the DeltaNp63: Their role in limbal epithelial cell proliferation. J. Cell. Physiol. 2013, 228, 536–546. [Google Scholar] [CrossRef]
- Hai, T.; Hartman, M.G. The molecular biology and nomenclature of the activating transcription factor/cAMP responsive element binding family of transcription factors: Activating transcription factor proteins and homeostasis. Gene 2001, 273, 1–11. [Google Scholar] [CrossRef]
- Persengiev, S.P.; Green, M.R. The role of ATF/CREB family members in cell growth, survival and apoptosis. Apoptosis Int. J. Program. Cell Death 2003, 8, 225–228. [Google Scholar] [CrossRef]
- Eaton, E.M.; Hanlon, M.; Bundy, L.; Sealy, L. Characterization of C/EBPbeta isoforms in normal versus neoplastic mammary epithelial cells. J. Cell. Physiol. 2001, 189, 91–105. [Google Scholar] [CrossRef]
- Tamura, K.; Hua, B.; Adachi, S.; Guney, I.; Kawauchi, J.; Morioka, M.; Tamamori-Adachi, M.; Tanaka, Y.; Nakabeppu, Y.; Sunamori, M.; et al. Stress response gene ATF3 is a target of c-myc in serum-induced cell proliferation. EMBO J. 2005, 24, 2590–2601. [Google Scholar] [CrossRef] [PubMed]
- Herberts, C.A.; Kwa, M.S.; Hermsen, H.P. Risk factors in the development of stem cell therapy. J. Transl. Med. 2011, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Lidzbarsky, G.; Gutman, D.; Shekhidem, H.A.; Sharvit, L.; Atzmon, G. Genomic Instabilities, Cellular Senescence, and Aging: In Vitro, In Vivo and Aging-Like Human Syndromes. Front. Med. 2018, 5, 104. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Kaghad, M.; Wang, Y.; Gillett, E.; Fleming, M.D.; Dötsch, V.; Andrews, N.C.; Caput, D.; McKeon, F. p63, a p53 Homolog at 3q27–29, Encodes Multiple Products with Transactivating, Death-Inducing, and Dominant-Negative Activities. Mol. Cell 1998, 2, 305–316. [Google Scholar] [CrossRef]
- Kouwenhoven, E.N.; van Heeringen, S.J.; Tena, J.J.; Oti, M.; Dutilh, B.E.; Alonso, M.E.; de la Calle-Mustienes, E.; Smeenk, L.; Rinne, T.; Parsaulian, L.; et al. Genome-Wide Profiling of p63 DNA–Binding Sites Identifies an Element that Regulates Gene Expression during Limb Development in the 7q21 SHFM1 Locus. PLoS Genet. 2010, 6, e1001065. [Google Scholar] [CrossRef] [PubMed]
- Kouwenhoven, E.N.; Oti, M.; Niehues, H.; van Heeringen, S.; Schalkwijk, J.; Stunnenberg, H.G.; van Bokhoven, H.; Zhou, H. Transcription factor p63 bookmarks and regulates dynamic enhancers during epidermal differentiation. EMBO Rep. 2015, 16, 863–878. [Google Scholar] [CrossRef]
- Cheng, C.C.; Wang, D.Y.; Kao, M.H.; Chen, J.K. The growth-promoting effect of KGF on limbal epithelial cells is mediated by upregulation of DeltaNp63alpha through the p38 pathway. J. Cell Sci. 2009, 122, 4473–4480. [Google Scholar] [CrossRef]
- Di Iorio, E.; Barbaro, V.; Ruzza, A.; Ponzin, D.; Pellegrini, G.; De Luca, M. Isoforms of DeltaNp63 and the migration of ocular limbal cells in human corneal regeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 9523–9528. [Google Scholar] [CrossRef]
- Kawasaki, S.; Tanioka, H.; Yamasaki, K.; Connon, C.J.; Kinoshita, S. Expression and tissue distribution of p63 isoforms in human ocular surface epithelia. Exp. Eye Res. 2006, 82, 293–299. [Google Scholar] [CrossRef]
- Perez, C.A.; Ott, J.; Mays, D.J.; A Pietenpol, J. p63 consensus DNA-binding site: Identification, analysis and application into a p63MH algorithm. Oncogene 2007, 26, 7363–7370. [Google Scholar] [CrossRef]
- Chiang, C.T.; Chu, W.K.; Chow, S.E.; Chen, J.K. Overexpression of delta Np63 in a human nasopharyngeal carcinoma cell line downregulates CKIs and enhances cell proliferation. J. Cell. Physiol. 2009, 219, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Allan, A.L.; Albanese, C.; Pestell, R.G.; LaMarre, J. Activating Transcription Factor 3 Induces DNA Synthesis and Expression of Cyclin D1 in Hepatocytes. J. Biol. Chem. 2001, 276, 27272–27280. [Google Scholar] [CrossRef] [PubMed]
- Perez, S.; Vial, E.; van Dam, H.; Castellazzi, M. Transcription factor ATF3 partially transforms chick embryo fibroblasts by promoting growth factor-independent proliferation. Oncogene 2001, 20, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Allen-Jennings, A.E.; Hartman, M.G.; Kociba, G.J.; Hai, T. The roles of ATF3 in glucose homeostasis. A transgenic mouse model with liver dysfunction and defects in endocrine pancreas. J. Biol. Chem. 2001, 276, 29507–29514. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Wolfgang, C.D.; Hai, T. Activating transcription factor 3, a stress-inducible gene, suppresses Ras-stimulated tumorigenesis. J. Biol. Chem. 2006, 281, 10473–10481. [Google Scholar] [CrossRef] [PubMed]
- Nobori, K.; Ito, H.; Tamamori-Adachi, M.; Adachi, S.; Ono, Y.; Kawauchi, J.; Kitajima, S.; Marumo, F.; Isobe, M. ATF3 inhibits doxorubicin-induced apoptosis in cardiac myocytes: A novel cardioprotective role of ATF3. J. Mol. Cell. Cardiol. 2002, 34, 1387–1397. [Google Scholar] [CrossRef]
- James, C.G.; Woods, A.; Underhill, T.M.; Beier, F. The transcription factor ATF3 is upregulated during chondrocyte differentiation and represses cyclin D1 and A gene transcription. BMC Mol. Biol. 2006, 7, 30. [Google Scholar] [CrossRef]
- Wang, D.-Y.; Hsueh, Y.-J.; Yang, V.C.; Chen, J.-K. Propagation and phenotypic preservation of rabbit limbal epithelial cells on amniotic membrane. Investig. Opthalmol. Vis. Sci. 2003, 44, 4698–4704. [Google Scholar] [CrossRef]
- Tsai, R.J.-F.; Li, L.-M.; Chen, J.-K. Reconstruction of Damaged Corneas by Transplantation of Autologous Limbal Epithelial Cells. N. Engl. J. Med. 2000, 343, 86–93. [Google Scholar] [CrossRef]
- Tseng, S.C.; Chen, S.-Y.; Mead, O.G.; Tighe, S. Niche regulation of limbal epithelial stem cells: HC-HA/PTX3 as surrogate matrix niche. Exp. Eye Res. 2020, 199, 108181. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Sun, C.C.; Yu, W.H.; Hsieh, H.L.; Ma, D.H.; Pang, J.H.; Yang, C.M. Novel laminin 5 gamma 2-chain fragments potentiating the limbal epithelial cell outgrowth on amniotic membrane. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4631–4639. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.B.; Larochelle, M.B.; Pedler, M.G.; Petrash, J.M.; Enzenauer, R.W. The effect of amniotic membrane grafting on healing and wound strength after strabismus surgery in a rabbit model. J. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2018, 22, 22–26.e1. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.P.; Tassone, D.; Ding, N.S.; Nossent, J. Antibody response to the COVID-19 ChAdOx1nCov-19 and BNT162b vaccines after temporary suspension of DMARD therapy in immune-mediated inflammatory disease: An extension study (RESCUE 2). RMD Open 2023, 9, e002871. [Google Scholar] [CrossRef]
- Ramos da Silva, J.; Bitencourt Rodrigues, K.; Formoso Pelegrin, G.; Silva Sales, N.; Muramatsu, H.; de Oliveira Silva, M.; Porchia, B.; Moreno, A.C.R.; Aps, L.; Venceslau-Carvalho, A.A.; et al. Single immunizations of self-amplifying or non-replicating mRNA-LNP vaccines control HPV-associated tumors in mice. Sci. Transl. Med. 2023, 15, eabn3464. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Entrez Gene No. | Gene Description | Array (log2) |
|---|---|---|---|
| ATF3 | 467 | Activating transcription factor 3 | 3.764 |
| JUN | 3725 | Jun oncogene | 1.621 |
| EGR1 | 1958 | Early growth response 1 | 1.391 |
| PAX6 | 5080 | Paired box protein 6 | 0.893 |
| CEBPB | 1051 | CCAAT/enhancer-binding protein beta | 0.820 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsueh, Y.-J.; Meir, Y.-J.J.; Hsiao, H.-Y.; Cheng, C.-M.; Ma, H.-K.D.; Wu, W.-C.; Chen, H.-C. Transcription Factor ATF3 Participates in DeltaNp63-Mediated Proliferation of Corneal Epithelial Cells. J. Pers. Med. 2023, 13, 700. https://doi.org/10.3390/jpm13040700
Hsueh Y-J, Meir Y-JJ, Hsiao H-Y, Cheng C-M, Ma H-KD, Wu W-C, Chen H-C. Transcription Factor ATF3 Participates in DeltaNp63-Mediated Proliferation of Corneal Epithelial Cells. Journal of Personalized Medicine. 2023; 13(4):700. https://doi.org/10.3390/jpm13040700
Chicago/Turabian StyleHsueh, Yi-Jen, Yaa-Jyuhn James Meir, Hui-Yi Hsiao, Chao-Min Cheng, Hui-Kang David Ma, Wei-Chi Wu, and Hung-Chi Chen. 2023. "Transcription Factor ATF3 Participates in DeltaNp63-Mediated Proliferation of Corneal Epithelial Cells" Journal of Personalized Medicine 13, no. 4: 700. https://doi.org/10.3390/jpm13040700