
The Composition of Small Extracellular Vesicles (sEVs) in the Blood Plasma of Colorectal Cancer Patients Reflects the Presence of Metabolic Syndrome and Correlates with Angiogenesis and the Effectiveness of Thermoradiation Therapy
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Treatment
2.2. Measurement of Anthropometric Parameters and Blood Plasma Metabolic Markers
2.3. Immunohistochemical Staining and Assessment
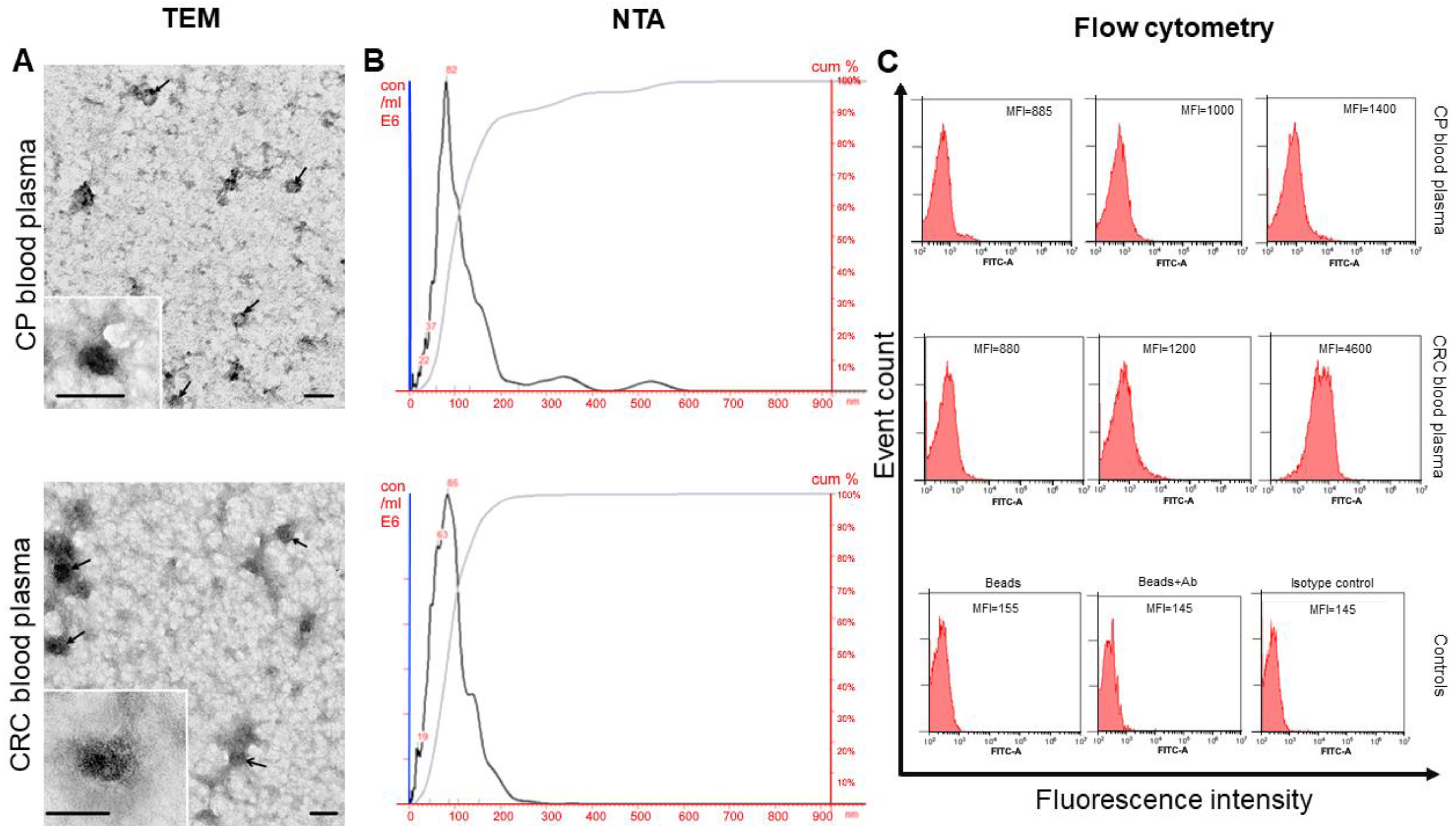
2.4. Small EVs Isolation, Electron Microscopy, and Nanoparticle Tracking Analysis
2.5. Flow Cytometry
2.5.1. Analysis of CD9/CD63/CD81/CD24 Subpopulations in Plasma sEVs
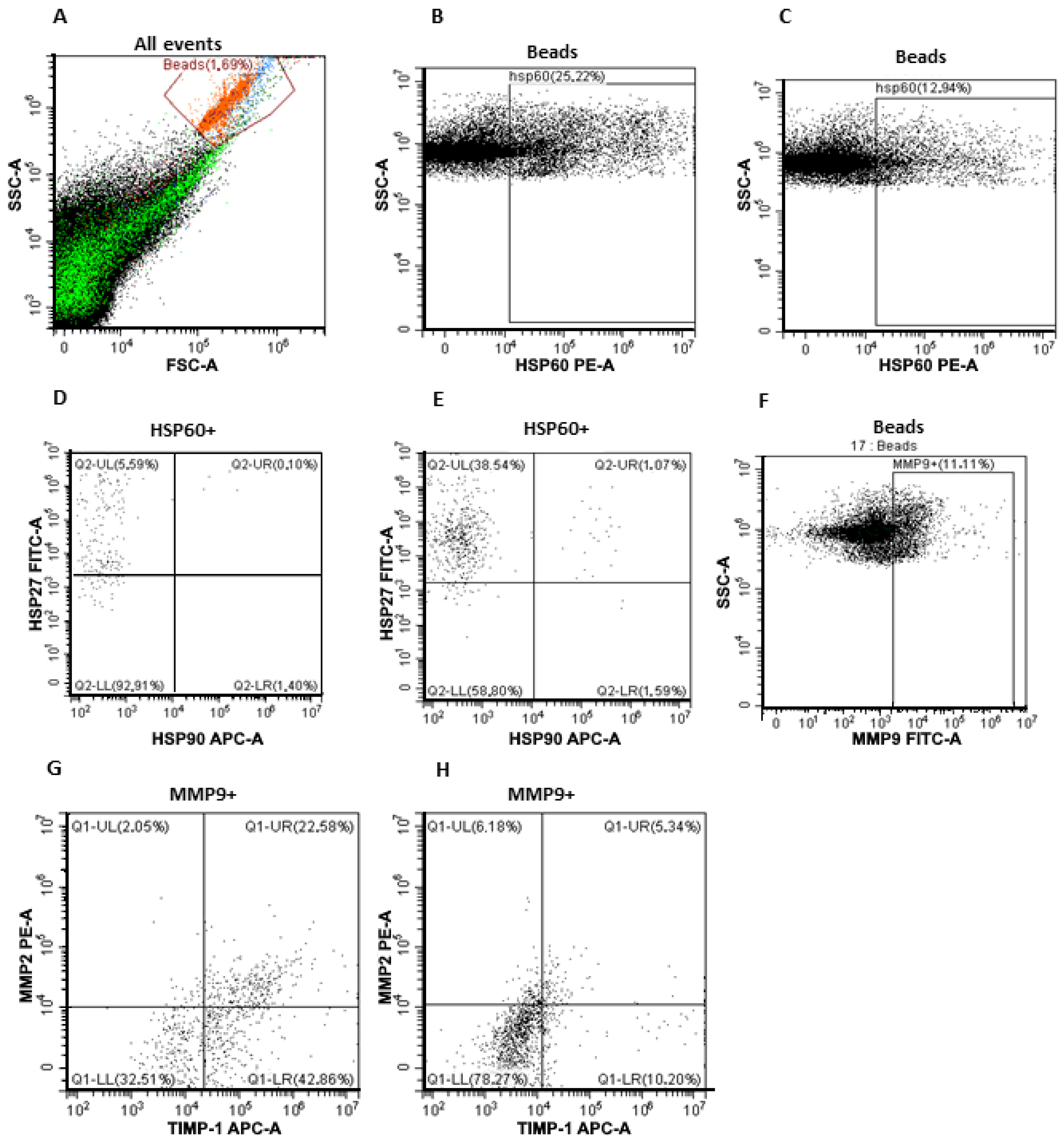
2.5.2. Analysis of HSP60/HSP27/HSP90 Subpopulations on the Surface of CD9-Positive sEVs
2.5.3. Analysis of MMP9/MMP2/TIMP1 Subpopulations on the Surface of CD9-Positive sEVs
2.5.4. Analysis of HSP60/HSP27/HSP90 and MMP9/MMP2/TIMP1 Subpopulations on the Surface of FABP4-Positive sEVs
2.5.5. Analysis of Tspan8/CD151 Subpopulations in Plasma EVs. Evaluation of the Tspan8/CD151 Tetraspanin Subpopulations in Plasma sEVs
2.6. Statistical Analysis
3. Results
3.1. Characteristics of Isolated sEVs
3.2. Subpopulations of MMPs and HSPs on the Surface of CD9-Positive and FABP4-Positive Plasma sEVs from CPPs and CRCPs
3.3. The Expression of MMPs, TIMP1, and HSPs on the Surface of CD9-Positive and FABP4-Positive sEVs Related to Body Mass Index, Plasma Triglycerides, and Plasma HDL Cholesterol Levels in CRCPs
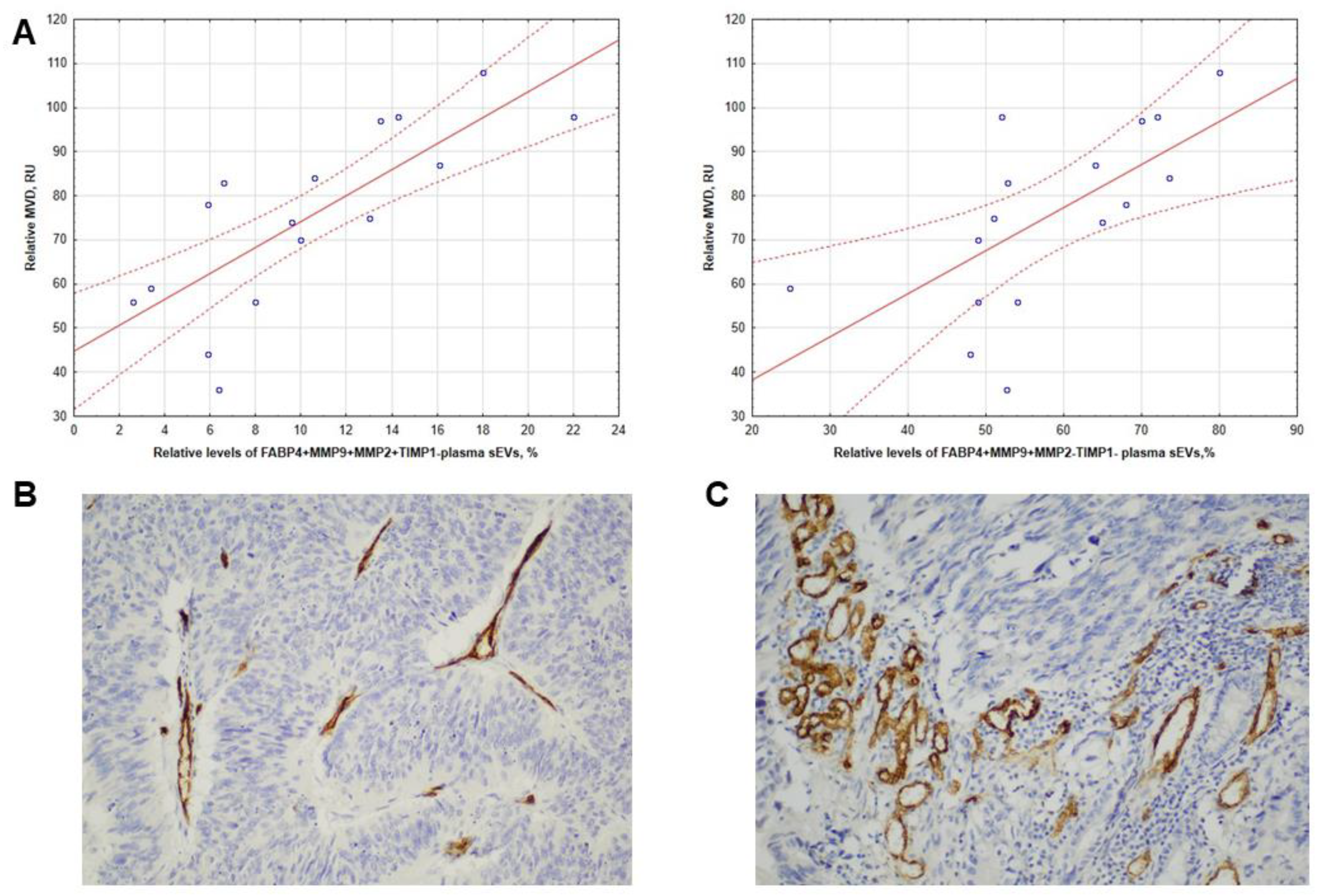
3.4. Intratumoral CD31 Expression in CRCPs Related to the Level of FABP4+MMP9+MMP2+TIMP1- and FABP4+MMP9+MMP2-TIMP1- Blood Plasma sEVs
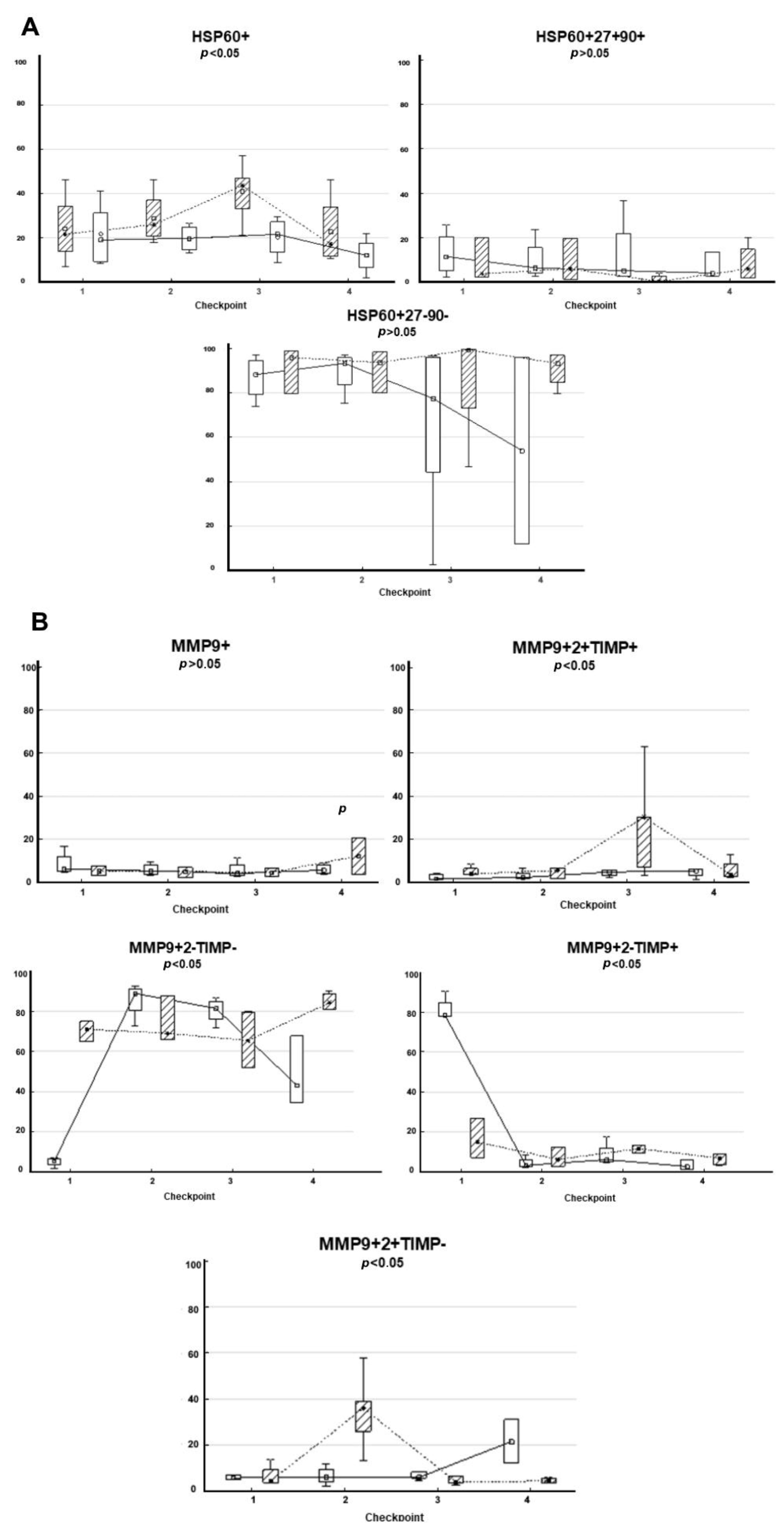
3.5. Composition of HSPs and MMPs on the Surface of CD9-Positive sEVs Derived from RCPs during Combined Treatment with Thermoradiotherapy: Relationship with Treatment Efficacy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yunusova, N.V.; Kondakova, I.V.; Kolomiets, L.A.; Afanas’ev, S.G.; Kishkina, A.Y.; Spirina, L.V. The role of metabolic syndrome variant in the malignant tumors progression. Diabetes Metab. Syndr. 2018, 12, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Qian, J.M.; Li, J.N. The metabolic syndrome and its components as prognostic factors in colorectal cancer: A meta-analysis and systematic review. J. Gastroenterol. Hepatol. 2023, 38, 187–196. [Google Scholar] [CrossRef]
- Wang, J.; Wu, Y.; Guo, J.; Fei, X.; Yu, L.; Ma, S. Adipocyte-derived exosomes promote lung cancer metastasis by increasing MMP9 activity via transferring MMP3 to lung cancer cells. Oncotarget 2017, 8, 81880–81891. [Google Scholar] [CrossRef] [PubMed]
- Yunusova, N.V.; Tugutova, E.A.; Tamkovich, S.N.; Kondakova, I.V. The role of exosomal tetraspanins and proteases in tumor progression. Biochem. Mosc. Suppl. Ser. B 2018, 12, 191–202. [Google Scholar] [CrossRef]
- Sandvig, K.; Llorente, A. Proteomic analysis of microvesicles released by the human prostate cancer cell line PC-3. Mol. Cell. Proteom. 2012, 11, M111-012914. [Google Scholar] [CrossRef] [PubMed]
- Hemler, M.E. Tetraspanin proteins promote multiple cancer stages. Nat. Rev. Cancer 2014, 14, 49–60. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Gao, X.; Salomon, C.; Freeman, D.J. Extracellular vesicles from adipose tissue-a potential role in obesity and type 2 diabetes? Front. Endocrinol. 2017, 8, 202. [Google Scholar] [CrossRef]
- Furuhashi, M. Fatty acid-binding protein 4 in cardiovascular and metabolic diseases. J. Atheroscler. Thromb. 2019, 26, 216–232. [Google Scholar] [CrossRef]
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating adipocyte-derived extracellular vesicles are novel markers of metabolic stress. J. Mol. Med. 2016, 94, 1241–1253. [Google Scholar] [CrossRef]
- DeClercq, V.; d’Eon, B.; McLeod, R.S. Fatty acids increase adiponectin secretion through both classical and exosome pathways. Biochim. Biophys. Acta 2015, 1851, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Durcin, M.; Fleury, A.; Taillebois, E.; Hilairet, G.; Krupova, Z.; Henry, C.; Truchet, S.; Trötzmüller, M.; Köfeler, H.; Mabilleau, G.; et al. Characterisation of adipocyte-derived extracellular vesicle subtypes identifies distinct protein and lipid signatures for large and small extracellular vesicles. J. Extracell. Vesicles 2017, 6, 1305677. [Google Scholar] [CrossRef] [PubMed]
- Buzas, E.I.; Toth, E.A.; Sodar, B.W.; Szabo-Taylor, K.E. Molecular interactions at the surface of extracellular vesicles. Semin. Immunopathol. 2018, 40, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Khokha, R. Metalloproteinases in extracellular vesicles. Biochim. Biophys. Acta Mol. Cell. Res. 2017, 1864 Pt A, 1989–2000. [Google Scholar] [CrossRef]
- Kaur, P.; Hurwitz, M.D.; Krishnan, S.; Asea, A. Combined hyperthermia and radiotherapy for the treatment of cancer. Cancers 2011, 3, 3799–3823. [Google Scholar] [CrossRef]
- Taha, E.A.; Ono, K.; Eguchi, T. Roles of extracellular HSPs as biomarkers in immune surveillance and immune evasion. Int. J. Mol. Sci. 2019, 20, 4588. [Google Scholar] [CrossRef]
- Lv, L.H.; Wan, Y.L.; Lin, Y.; Zhang, W.; Yang, M.; Li, G.L.; Lin, H.M.; Shang, C.Z.; Chen, Y.J.; Min, J. Anticancer drugs cause release of exosomes with heat shock proteins from human hepatocellular carcinoma cells that elicit effective natural killer cell antitumor responses in vitro. J. Biol. Chem. 2012, 287, 15874–15885. [Google Scholar] [CrossRef]
- Barrea, L.; Muscogiuri, G.; Pugliese, G.; de Alteriis, G.; Colao, A.; Savastano, S. Metabolically Healthy Obesity (MHO) vs. Metabolically Unhealthy Obesity (MUO) Phenotypes in PCOS: Association with endocrine-metabolic profile, adherence to the mediterranean diet, and body composition. Nutrients 2021, 13, 3925. [Google Scholar] [CrossRef]
- Lambregts, D.M.J.; Boellaard, T.N.; Beets-Tan, R.G.H. Response evaluation after neoadjuvant treatment for rectal cancer using modern MR imaging: A pictorial review. Insight Imaging 2019, 10, 15. [Google Scholar] [CrossRef]
- Vermeulen, P.B.; Gasparini, G.; Fox, S.B.; Colpaert, C.; Marson, L.P.; Gion, M.; Beliën, J.A.; de Waal, R.M.; Van Marck, E.; Magnani, E.; et al. Second international consensus on the methodology and criteria of evaluation of angiogenesis quantification in solid human tumours. Eur. J. Cancer 2002, 38, 1564–1579. [Google Scholar] [CrossRef]
- Yunusova, N.V.; Zambalova, E.A.; Patysheva, M.R.; Kolegova, E.S.; Afanas’ev, S.G.; Cheremisina, O.V.; Grigor’eva, A.E.; Tamkovich, S.N.; Kondakova, I.V. Exosomal protease cargo as prognostic biomarker in colorectal cancer. Asian Pac. J. Cancer Prev. 2021, 22, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Yunusova, N.; Dzhugashvili, E.; Yalovaya, A.; Kolomiets, L.; Shefer, A.; Grigor’eva, A.; Tupikin, A.; Kondakova, I.; Tamkovich, S. Comparative analysis of tumor-associated microRNAs and tetraspanines from exosomes of plasma and ascitic fluids of ovarian cancer patients. Int. J. Mol. Sci. 2022, 24, 464. [Google Scholar] [CrossRef] [PubMed]
- Tamkovich, S.N.; Yunusova, N.V.; Somov, A.K.; Afanas’ev, S.G.; Kakurina, G.V.; Kolegova, E.S.; Tugutova, E.A.; Laktionov, P.P.; Kondakova, I.V. Comparative subpopulation analysis of plasma exosomes from cancer patients. Biochem. Mosc. Suppl. Ser. B 2018, 12, 151–155. [Google Scholar] [CrossRef]
- Kralisch, S.; Ebert, T.; Lossner, U.; Jessnitzer, B.; Stumvoll, M.; Fasshauer, M. Adipocyte fatty acid-binding protein is released from adipocytes by a non-conventional mechanism. Int. J. Obes. 2014, 38, 1251–1254. [Google Scholar] [CrossRef] [PubMed]
- Hubal, M.J.; Nadler, E.P.; Ferrante, S.C.; Barberio, M.D.; Suh, J.H.; Wang, J.; Dohm, G.L.; Pories, W.J.; Mietus-Snyder, M.; Freishtat, R.J. Circulating adipocyte-derived exosomal microRNAs associated with decreased insulin resistance after gastric bypass. Obesity 2017, 25, 102–110. [Google Scholar] [CrossRef]
- Gustafson, C.M.; Shepherd, A.J.; Miller, V.M.; Jayachandran, M. Age- and sex-specific differences in blood-borne microvesicles from apparently healthy humans. Biol. Sex Differ. 2015, 6, 10. [Google Scholar] [CrossRef]
- Bavisotto, C.C.; Cipolla, C.; Graceffa, G.; Barone, R.; Bucchieri, F.; Bulone, D.; Cabibi, D.; Campanella, C.; Gammazza, A.M.; Pitruzzella, A.; et al. Immumirphological pattern of molecular chaperones in normal and pathological thyroid tissues and circulating exosomes: Potential use in clinics. Int. J. Mol. Sci. 2019, 20, 4496. [Google Scholar] [CrossRef]
- Freeman, D.W.; Noren Hooten, N.; Eitan, E.; Green, J.; Mode, N.A.; Bodogai, M.; Zhang, Y.; Lehrmann, E.; Zonderman, A.B.; Biragyn, A.; et al. Altered extracellular vesicle concentration, cargo, and function in diabetes. Diabetes 2018, 67, 2377–2388. [Google Scholar] [CrossRef]
- Todorova, D.; Simoncini, S.; Lacroix, R.; Sabatier, F.; Dignat-George, F. Extracellular vesicles in angiogenesis. Circ. Res. 2017, 120, 1658–1673. [Google Scholar] [CrossRef]
- Aslan, C.; Maralbashi, S.; Salari, F.; Kahroba, H.; Sigaroodi, F.; Kazemi, T.; Kharaziha, P. Tumor-derived exosomes: Implication in angiogenesis and antiangiogenesis cancer therapy. J. Cell. Physiol. 2019, 234, 16885–16903. [Google Scholar] [CrossRef]
- Huang, M.; Lei, Y.; Zhong, Y.; Chung, C.; Wang, M.; Hu, M.; Deng, L. New insights into the regulatory roles of extracellular vesicles in tumor angiogenesis and their clinical implications. Front. Cell. Dev. Biol. 2021, 9, 791882. [Google Scholar] [CrossRef] [PubMed]
- Konoshenko, M.; Sagaradze, G.; Orlova, E.; Shtam, T.; Proskura, K.; Kamyshinsky, R.; Yunusova, N.; Alexandrova, A.; Efimenko, A.; Tamkovich, S. Total blood exosomes in breast cancer: Potential role in crucial steps of tumorigenesis. Int. J. Mol. Sci. 2020, 21, 7341. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotypes of EVs, % | Groups | p-Level | |
|---|---|---|---|
| CPPs (n = 10) | CRCPs (n = 22) | ||
| HSP60+ | 19.4 ± 1.86 | 11.10 ± 1.21 | <0.05 |
| HSP60+HSP27+HSP90- | 6.30 ± 3.26 | 26.5 ± 3.01 | <0.05 |
| HSP60+HSP27-HSP90- | 91.7 ± 3.52 | 73.0 ± 2.91 | <0.05 |
| HSP60+HSP27-HSP90+ | 1.35 ± 0.55 | 1.49 ± 0.25 | |
| MMP9+ | 6.45 ± 1.82 | 11.0 ± 1.02 | <0.05 |
| MMP9+MMP2+TIMP1+ | 8.70 ± 1.02 | 5.80 ± 0.93 | <0.05 |
| MMP9+MMP2+TIMP1- | 11.0 ± 0.92 | 9.70 ± 1.37 | |
| MMP9+MMP2-TIMP1- | 71.7 ± 10.2 | 75.3 ± 4.50 | |
| MM9+MMP2-TIMP1+ | 11.4 ± 3.71 | 10.4 ± 2.85 | |
| Phenotypes of EVs, % | Groups | p-Level | |
|---|---|---|---|
| CPPs (n = 10) | CRCPs (n = 22) | ||
| HSP60+ | 18.9 ± 3.10 | 19.5 ± 2.21 | |
| HSP60+HSP27+HSP90- | 24.1 ± 3.90 | 23.7 ± 3.22 | |
| HSP60+HSP27-HSP90- | 74.7 ± 5.55 | 75.0 ± 5.35 | |
| HSP60+HSP27-HSP90+ | 0.95 ± 0.35 | 1.05 ± 0.24 | |
| MMP9+ | 8.33 ± 1.15 | 11.2 ± 2.00 | |
| MMP9+MMP2+TIMP1+ | 3.33 ± 1.71 | 10.9 ± 2.40 | <0.05 |
| MMP9+MMP2+TIMP1- | 10.6 ± 2.53 | 10.3 ± 1.77 | |
| MMP9+MMP2-TIMP1- | 82.3 ± 5.83 | 61.3 ± 6.38 | <0.05 |
| MMP9+MMP2-TIMP1+ | 3.73 ± 0.65 | 17.4 ± 3.10 | <0.05 |
| Phenotypes of EVs, % | Age | BMI | Waist Circumference | TGs | Total Cholesterol | HDL Cholesterol |
|---|---|---|---|---|---|---|
| HSP60+ | ||||||
| HSP60+HSP27+HSP90- | 0.61 | |||||
| HSP60+HSP27-HSP90- | −0.61 | |||||
| HSP60+HSP27-HSP90+ | −0.61 | −0.68 | ||||
| MMP9+ | 0.53 | 0.60 | ||||
| MMP9+MMP2+TIMP1+ | 0.57 | |||||
| MMP9+MMP2+TIMP1- | ||||||
| MMP9+MMP2-TIMP1- | ||||||
| MMP9+MMP2-TIMP1+ | 0.56 | 0.62 |
| Phenotypes of EVs, % | Age | BMI | Waist Circumference | TGs | Total Cholesterol | HDL Cholesterol |
|---|---|---|---|---|---|---|
| HSP60+ | −0.59 | |||||
| HSP60+HSP27+HSP90- | ||||||
| HSP60+HSP27-HSP90- | 0.53 | 0.54 | ||||
| HSP60+HSP27-HSP90+ | −0.68 | −0.79 | ||||
| MMP9+ | −0.74 | |||||
| MMP9+MMP2+TIMP1+ | 0.67 | |||||
| MMP9+MMP2+TIMP1- | ||||||
| MMP9+MMP2-TIMP1- | ||||||
| MMP9+MMP2-TIMP1+ | 0.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yunusova, N.V.; Svarovsky, D.A.; Konovalov, A.I.; Kostromitsky, D.N.; Startseva, Z.A.; Cheremisina, O.V.; Afanas’ev, S.G.; Kondakova, I.V.; Grigor’eva, A.E.; Vtorushin, S.V.; et al. The Composition of Small Extracellular Vesicles (sEVs) in the Blood Plasma of Colorectal Cancer Patients Reflects the Presence of Metabolic Syndrome and Correlates with Angiogenesis and the Effectiveness of Thermoradiation Therapy. J. Pers. Med. 2023, 13, 684. https://doi.org/10.3390/jpm13040684
Yunusova NV, Svarovsky DA, Konovalov AI, Kostromitsky DN, Startseva ZA, Cheremisina OV, Afanas’ev SG, Kondakova IV, Grigor’eva AE, Vtorushin SV, et al. The Composition of Small Extracellular Vesicles (sEVs) in the Blood Plasma of Colorectal Cancer Patients Reflects the Presence of Metabolic Syndrome and Correlates with Angiogenesis and the Effectiveness of Thermoradiation Therapy. Journal of Personalized Medicine. 2023; 13(4):684. https://doi.org/10.3390/jpm13040684
Chicago/Turabian StyleYunusova, Natalia V., Dmitry A. Svarovsky, Artem I. Konovalov, Dmitry N. Kostromitsky, Zhanna A. Startseva, Olga V. Cheremisina, Sergey G. Afanas’ev, Irina V. Kondakova, Alina E. Grigor’eva, Sergey V. Vtorushin, and et al. 2023. "The Composition of Small Extracellular Vesicles (sEVs) in the Blood Plasma of Colorectal Cancer Patients Reflects the Presence of Metabolic Syndrome and Correlates with Angiogenesis and the Effectiveness of Thermoradiation Therapy" Journal of Personalized Medicine 13, no. 4: 684. https://doi.org/10.3390/jpm13040684