
Susceptibility to Colorectal Cancer Based on HSD17B4 rs721673 and rs721675 Polymorphisms and Alcohol Intake among Taiwan Biobank Participants: A Retrospective Case Control Study Using the Nationwide Claims Data
 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Data Sources
2.2. Ethical Considerations
2.3. Study Population
2.4. Covariate Assessment
2.5. Genotyping Data and Imputation
2.6. Statistical Analyses
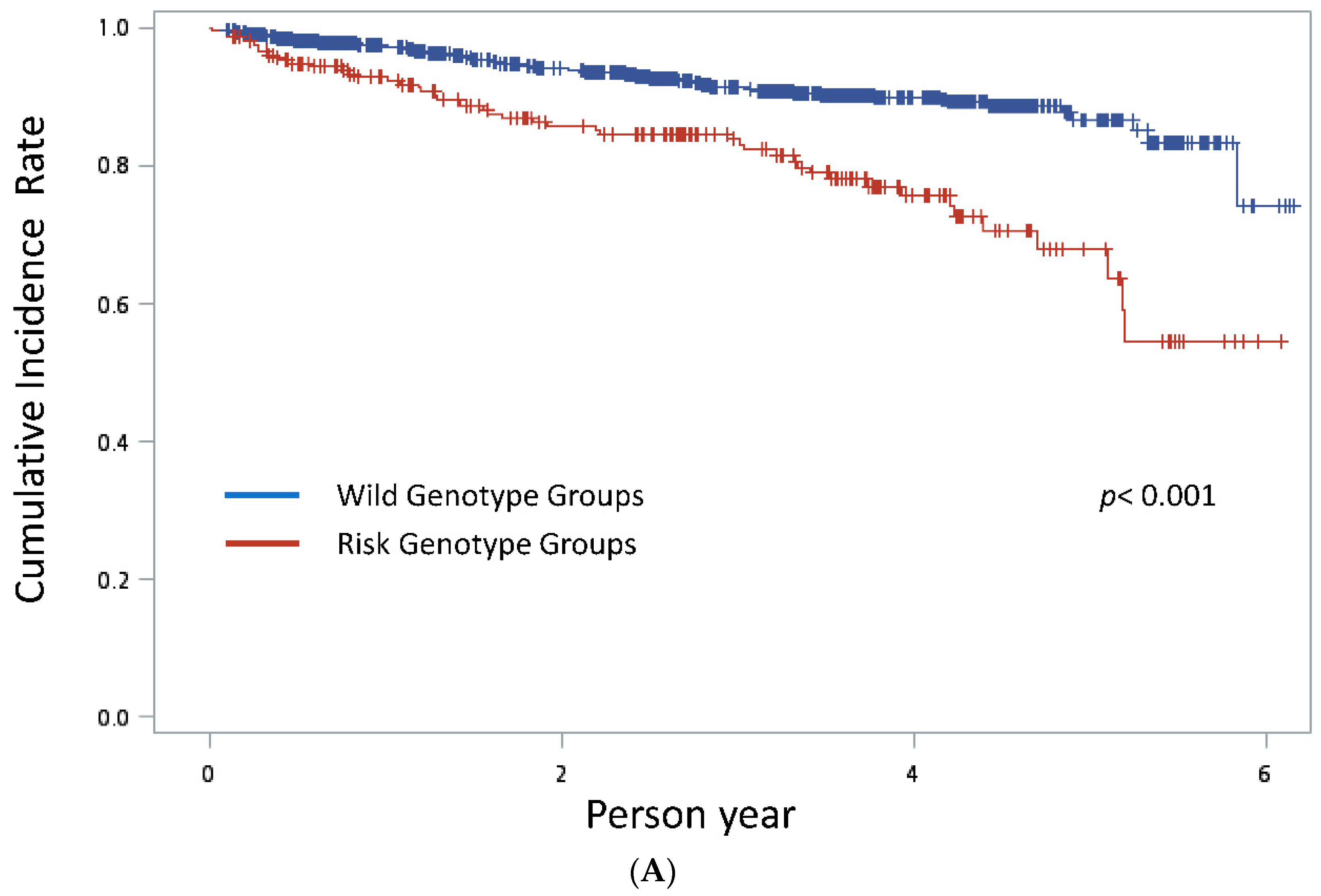
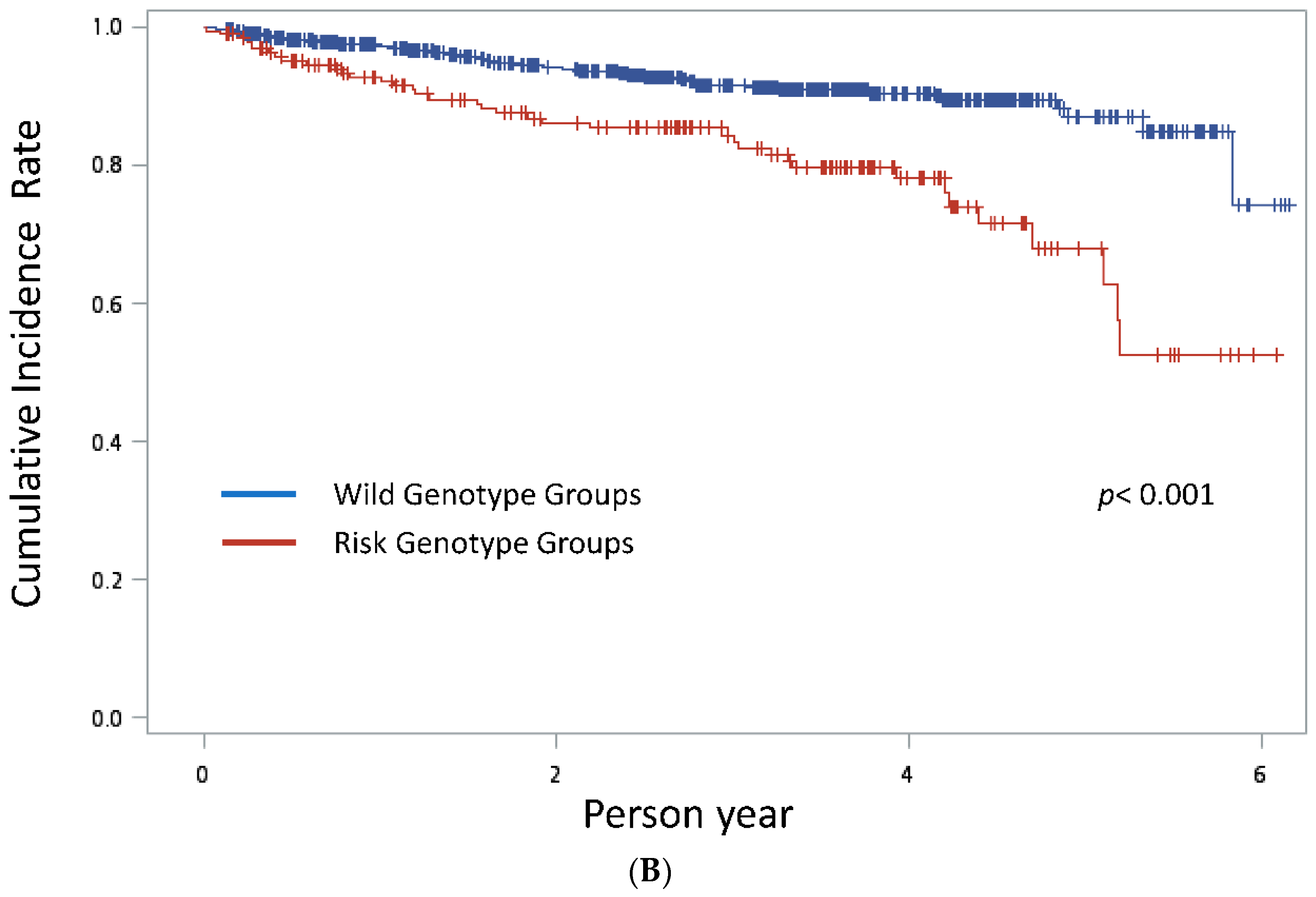
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- 2017 Cancer Registry Annual Report; Ministry of Health and Welfare: Taipei City, Taiwan, 2017.
- 2018 Cause of Death Statistics; Ministry of Health and Welfare: Taipei City, Taiwan, 2018.
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Przeglad Gastroenterol. 2019, 14, 89. [Google Scholar] [CrossRef]
- Graff, R.E.; Möller, S.; Passarelli, M.N.; Witte, J.S.; Skytthe, A.; Christensen, K.; Tan, Q.; Adami, H.-O.; Czene, K.; Harris, J.R. Familial risk and heritability of colorectal cancer in the nordic twin study of cancer. Clin. Gastroenterol. Hepatol. 2017, 15, 1256–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Jia, W.H.; Matsuo, K.; Shin, A.; Xiang, Y.B.; Matsuda, K.; Jee, S.H.; Kim, D.H.; Cheah, P.Y.; Ren, Z. Genome-wide association study identifies a new SMAD7 risk variant associated with colorectal cancer risk in East Asians. Int. J. Cancer 2014, 135, 948–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, R.; Nejati, F.; Nasiri, N.; Khodabakhsh, P.; Khodabakhsh, P. Association of APC and MUTYH Mutations with Colorectal Cancer; Research Square: Durham, NC, USA, 2021. [Google Scholar]
- Valle, L.; Hernandez-Illan, E.; Bellido, F.; Aiza, G.; Castillejo, A.; Castillejo, M.-I.; Navarro, M.; Segui, N.; Vargas, G.; Guarinos, C. New insights into POLE and POLD1 germline mutations in familial colorectal cancer and polyposis. Hum. Mol. Genet. 2014, 23, 3506–3512. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.; Hearle, N.; Shah, B.; Murday, V.; Hodgson, S.; Lucassen, A.; Eccles, D.; Talbot, I.; Neale, K.; Lim, A. Further observations on LKB1/STK11 status and cancer risk in Peutz–Jeghers syndrome. Br. J. Cancer 2003, 89, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Sayed, M.; Ahmed, A.; Ringold, J.; Anderson, M.; Bair, J.; Mitros, F.; Lynch, H.; Tinley, S.; Petersen, G.; Giardiello, F. Germlinesmad4 orbmpria mutations and phenotype of juvenile polyposis. Annal. Surg. Oncol. 2002, 9, 901–906. [Google Scholar] [CrossRef]
- Lammi, L.; Arte, S.; Somer, M.; Järvinen, H.; Lahermo, P.; Thesleff, I.; Pirinen, S.; Nieminen, P. Mutations in AXIN2 cause familial tooth agenesis and predispose to colorectal cancer. Am. J. Hum. Genet. 2004, 74, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.-L.; Kawabata, M.; Imamura, T.; Akiyama, Y.; Nomizu, T.; Miyazono, K.; Yuasa, Y. HNPCC associated with germline mutation in the TGF-β type II receptor gene. Nat. Genet. 1998, 19, 17–18. [Google Scholar] [CrossRef]
- De la Chapelle, A. Genetic predisposition to colorectal cancer. Nat. Rev. Cancer 2004, 4, 769–780. [Google Scholar] [CrossRef]
- Jasperson, K.W.; Tuohy, T.M.; Neklason, D.W.; Burt, R.W. Hereditary and familial colon cancer. Gastroenterology 2010, 138, 2044–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, E.E.; Kluska, A.; Piątkowska, M.; Kulecka, M.; Bałabas, A.; Zeber-Lubecka, N.; Goryca, K.; Ambrożkiewicz, F.; Karczmarski, J.; Olesiński, T. GWAS Links New Variant in Long Non-Coding RNA LINC02006 with Colorectal Cancer Susceptibility. Biology 2021, 10, 465. [Google Scholar] [CrossRef] [PubMed]
- Jostins, L.; Barrett, J.C. Genetic risk prediction in complex disease. Hum. Mol. Genet. 2011, 20, R182–R188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, P.S.; Chen, T.Y.; Giovannucci, E. Cigarette smoking and colorectal cancer incidence and mortality: Systematic review and meta-analysis. Int. J. Cancer 2009, 124, 2406–2415. [Google Scholar] [CrossRef]
- Moghaddam, A.A.; Woodward, M.; Huxley, R. Obesity and risk of colorectal cancer: A meta-analysis of 31 studies with 70,000 events. Cancer Epidemiol. Biomark. Prev. 2007, 16, 2533–2547. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.; Jahanzaib Anwar, M.; Usman, A.; Keshavarzian, A.; Bishehsari, F. Colorectal cancer and alcohol consumption—Populations to molecules. Cancers 2018, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- Botteri, E.; Borroni, E.; Sloan, E.K.; Bagnardi, V.; Bosetti, C.; Peveri, G.; Santucci, C.; Specchia, C.; van den Brandt, P.; Gallus, S. Smoking and colorectal cancer risk, overall and by molecular subtypes: A meta-analysis. Off. J. Am. Coll. Gastroenterol. ACG 2020, 115, 1940–1949. [Google Scholar] [CrossRef]
- Shaukat, A.; Dostal, A.; Menk, J.; Church, T.R. BMI is a risk factor for colorectal cancer mortality. Dig. Dis. Sci. 2017, 62, 2511–2517. [Google Scholar] [CrossRef]
- Jemal, A.; Center, M.M.; DeSantis, C.; Ward, E.M. Global Patterns of Cancer Incidence and Mortality Rates and TrendsGlobal Patterns of Cancer. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1893–1907. [Google Scholar] [CrossRef] [Green Version]
- Bishehsari, F.; Magno, E.; Swanson, G.; Desai, V.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Alcohol and gut-derived inflammation. Alcohol Res. Curr. Rev. 2017, 38, 163. [Google Scholar]
- Varela-Rey, M.; Woodhoo, A.; Martinez-Chantar, M.-L.; Mato, J.M.; Lu, S.C. Alcohol, DNA methylation, and cancer. Alcohol Res. 2013, 35, 25–35. [Google Scholar] [PubMed]
- Cui, R.; Okada, Y.; Jang, S.; Ku, J.; Park, J.; Kamatani, Y.; Hosono, N.; Tsunoda, T.; Kumar, V.; Tanikawa, C. Common variant in 6q26-q27 is associated with distal colon cancer in an Asian population. Gut 2011, 60, 799–805. [Google Scholar] [CrossRef] [Green Version]
- Hang, D.; Joshi, A.D.; He, X.; Chan, A.T.; Jovani, M.; Gala, M.K.; Ogino, S.; Kraft, P.; Turman, C.; Peters, U. Colorectal cancer susceptibility variants and risk of conventional adenomas and serrated polyps: Results from three cohort studies. Int. J. Epidemiol. 2020, 49, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Carr, P.R.; Weigl, K.; Edelmann, D.; Jansen, L.; Chang-Claude, J.; Brenner, H.; Hoffmeister, M. Estimation of absolute risk of colorectal cancer based on healthy lifestyle, genetic risk, and colonoscopy status in a population-based study. Gastroenterology 2020, 159, 129–138.e9. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Jia, G.; Wen, W.; Shu, X.-O.; Zheng, W. Healthy lifestyles, genetic modifiers, and colorectal cancer risk: A prospective cohort study in the UK Biobank. Am. J. Clin. Nutr. 2021, 113, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.-T.; Hung, T.-H.; Yeh, C.-K. Taiwan regulation of biobanks. SAGE J. 2015, 43, 4. [Google Scholar] [CrossRef]
- Hsieh, C.-Y.; Su, C.-C.; Shao, S.-C.; Sung, S.-F.; Lin, S.-J.; Yang, Y.-H.K.; Lai, E.C.-C. Taiwan’s national health insurance research database: Past and future. Clin. Epidemiol. 2019, 11, 349. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.S.-H.; Hsieh, C.-H.; Pei, D.; Hung, Y.-J.; Kuo, S.-W.; Lin, E. Association and interaction analyses of genetic variants in ADIPOQ, ENPP1, GHSR, PPARγ and TCF7L2 genes for diabetic nephropathy in a Taiwanese population with type 2 diabetes. Nephrol. Dialysis Transplant. 2009, 24, 3360–3366. [Google Scholar] [CrossRef] [Green Version]
- Chou, W.C.; Hsiung, C.N.; Chen, W.T.; Tseng, L.M.; Wang, H.C.; Chu, H.W.; Hou, M.F.; Yu, J.C.; Shen, C.Y. A functional variant near XCL1 gene improves breast cancer survival via promoting cancer immunity. Int. J. Cancer 2020, 146, 2182–2193. [Google Scholar] [CrossRef]
- Chen, C.-H.; Yang, J.-H.; Chiang, C.W.; Hsiung, C.-N.; Wu, P.-E.; Chang, L.-C.; Chu, H.-W.; Chang, J.; Song, I.-W.; Yang, S.-L. Population structure of Han Chinese in the modern Taiwanese population based on 10,000 participants in the Taiwan Biobank project. Hum. Mol. Genet. 2016, 25, 5321–5331. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.-J.; Tsai, S.-J.; Kuo, P.-H.; Liu, Y.-L.; Yang, A.C.; Lin, E.; Lan, T.-H. An association study in the Taiwan Biobank reveals RORA as a novel locus for sleep duration in the Taiwanese population. Sleep Med. 2020, 73, 70–75. [Google Scholar] [CrossRef]
- Consortium, G.; Ardlie, K.G.; Deluca, D.S.; Segrè, A.V.; Sullivan, T.J.; Young, T.R.; Gelfand, E.T.; Trowbridge, C.A.; Maller, J.B.; Tukiainen, T. The Genotype-Tissue Expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef] [Green Version]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucl. Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [Green Version]
- Möller, G.; Leenders, F.; van Grunsven, E.G.; Dolez, V.; Qualmann, B.; Kessels, M.M.; Markus, M.; Krazeisen, A.; Husen, B.; Wanders, R.J. Characterization of the HSD17B4 gene: D-specific multifunctional protein 2/17β-hydroxysteroid dehydrogenase IV. J. Steroid Biochem. Mol. Biol. 1999, 69, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Breitling, R.; Marijanović, Z.; Perović, D.; Adamski, J. Evolution of 17β-HSD type 4, a multifunctional protein of β-oxidation. Mol. Cell Endocrinol. 2001, 171, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Peltoketo, H.; Simard, J.; Adamski, J. 17beta-hydroxysteroid dehydrogenase (HSD)/17-ketosteroid reductase (KSR) family; nomenclature and main characteristics of the 17HSD/KSR enzymes. J. Mol. Endocrinol. 1999, 23, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cavalieri, E.; Frenkel, K.; Liehr, J.G.; Rogan, E.; Roy, D. Chapter 4: Estrogens as endogenous genotoxic agents—DNA adducts and mutations. JNCI Monogr. 2000, 2000, 75–94. [Google Scholar] [CrossRef] [Green Version]
- Cavalieri, E.; Chakravarti, D.; Guttenplan, J.; Hart, E.; Ingle, J.; Jankowiak, R.; Muti, P.; Rogan, E.; Russo, J.; Santen, R. Catechol estrogen quinones as initiators of breast and other human cancers: Implications for biomarkers of susceptibility and cancer prevention. Biochim. Biophys. Acta BBA Rev. Cancer 2006, 1766, 63–78. [Google Scholar] [CrossRef]
- Cavalieri, E.; Rogan, E. The 3, 4-Quinones of Estrone and Estradiol Are the Initiators of Cancer whereas Resveratrol and N-acetylcysteine Are the Preventers. Int. J. Mol. Sci. 2021, 22, 8238. [Google Scholar] [CrossRef] [PubMed]
- Miao, S.; Yang, F.; Wang, Y.; Shao, C.; Zava, D.T.; Ding, Q.; Shi, Y.E. 4-Hydroxy estrogen metabolite, causing genomic instability by attenuating the function of spindle-assembly checkpoint, can serve as a biomarker for breast cancer. Am. J. Transl. Res. 2019, 11, 4992. [Google Scholar]
- Rawłuszko-Wieczorek, A.A.; Marczak, Ł.; Horst, N.; Horbacka, K.; Krokowicz, P.; Jagodziński, P.P. Significance of intratissue estrogen concentration coupled with estrogen receptors levels in colorectal cancer prognosis. Oncotarget 2017, 8, 115546. [Google Scholar] [CrossRef] [Green Version]
- Honma, N.; Yamamoto, K.; Ohnaka, K.; Morita, M.; Toyomura, K.; Kono, S.; Muramatsu, M.; Arai, T.; Ueki, T.; Tanaka, M. Estrogen receptor-β gene polymorphism and colorectal cancer risk: Effect modified by body mass index and isoflavone intake. Int. J. Cancer 2013, 132, 951–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Albeniz, X.; Rudolph, A.; Hutter, C.; White, E.; Lin, Y.; Rosse, S.A.; Figueiredo, J.C.; Harrison, T.A.; Jiao, S.; Brenner, H. CYP24A1 variant modifies the association between use of oestrogen plus progestogen therapy and colorectal cancer risk. Br. J. Cancer 2016, 114, 221–229. [Google Scholar] [CrossRef]
- Huang, H.; Liu, R.; Huang, Y.; Feng, Y.; Fu, Y.; Chen, L.; Chen, Z.; Cai, Y.; Zhang, Y.; Chen, Y. Acetylation-mediated degradation of HSD17B4 regulates the progression of prostate cancer. Aging 2020, 12, 14699. [Google Scholar] [CrossRef]
- Lu, X.; Ma, P.; Shi, Y.; Yao, M.; Hou, L.; Zhang, P.; Jiang, L. NF-κB increased expression of 17β-hydroxysteroid dehydrogenase 4 promotes HepG2 proliferation via inactivating estradiol. Mol. Cell Endocrinol. 2015, 401, 1–11. [Google Scholar] [CrossRef]
- Yamashita, S.; Hattori, N.; Fujii, S.; Yamaguchi, T.; Takahashi, M.; Hozumi, Y.; Kogawa, T.; El-Omar, O.; Liu, Y.-Y.; Arai, N. Multi-omics analyses identify HSD17B4 methylation-silencing as a predictive and response marker of HER2-positive breast cancer to HER2-directed therapy. Sci. Rep. 2020, 10, 15530. [Google Scholar] [CrossRef]
- Meier, M.; Möller, G.; Adamski, J. Perspectives in Understanding the Role of Human 17β-Hydroxysteroid Dehydrogenases in Health and Disease. Ann. N. Y. Acad. Sci. 2009, 1155, 15–24. [Google Scholar] [CrossRef]
- Wong, M.C.; Ding, H.; Wang, J.; Chan, P.S.; Huang, J. Prevalence and risk factors of colorectal cancer in Asia. Intest. Res. 2019, 17, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A review of colorectal cancer in terms of epidemiology, risk factors, development, symptoms and diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef] [PubMed]
- Wimberly, A.L.; Forsyth, C.B.; Khan, M.W.; Pemberton, A.; Khazaie, K.; Keshavarzian, A. Ethanol-Induced Mast Cell-Mediated Inflammation Leads to Increased Susceptibility of Intestinal Tumorigenesis in the APCΔ468 Min Mouse Model of Colon Cancer. Alcohol. Clin. Exp. Res. 2013, 37, E199–E208. [Google Scholar] [CrossRef] [Green Version]
- Slattery, M.L.; Potter, J.; Caan, B.; Edwards, S.; Coates, A.; Ma, K.-N.; Berry, T.D. Energy balance and colon cancer—Beyond physical activity. Cancer Res. 1997, 57, 75–80. [Google Scholar]
- Cai, S.; Li, Y.; Ding, Y.; Chen, K.; Jin, M. Alcohol drinking and the risk of colorectal cancer death. Eur. J. Cancer Prev. 2014, 23, 532–539. [Google Scholar] [CrossRef]
- Tsuruya, A.; Kuwahara, A.; Saito, Y.; Yamaguchi, H.; Tenma, N.; Inai, M.; Takahashi, S.; Tsutsumi, E.; Suwa, Y.; Totsuka, Y. Major anaerobic bacteria responsible for the production of carcinogenic acetaldehyde from ethanol in the colon and rectum. Alcohol Alcohol. 2016, 51, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Jing, L.; Jin, C.-m.; Li, S.-s.; Zhang, F.-m.; Yuan, L.; Li, W.-m.; Sang, Y.; Li, S.; Zhou, L.-j. Chronic alcohol intake-induced oxidative stress and apoptosis: Role of CYP2E1 and calpain-1 in alcoholic cardiomyopathy. Mol. Cell Biochem. 2012, 359, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.-W.; Son, M.-J.; Abdelmegeed, M.A.; Banerjee, A.; Morgan, T.R.; Yoo, S.-H.; Song, B.-J. Binge alcohol promotes hypoxic liver injury through a CYP2E1–HIF-1α-dependent apoptosis pathway in mice and humans. Free Rad. Biol. Med. 2014, 77, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, S.D.; Lim, R.W. Epigenetic effects of ethanol on the liver and gastrointestinal system. Alcohol Res. Curr. Rev. 2013, 35, 47. [Google Scholar]
- Krattinger, R.; Boström, A.; Lee, S.M.; Thasler, W.E.; Schiöth, H.B.; Kullak-Ublick, G.A.; Mwinyi, J. Chenodeoxycholic acid significantly impacts the expression of miRNAs and genes involved in lipid, bile acid and drug metabolism in human hepatocytes. Life Sci. 2016, 156, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.-S.; Lee, H.-W.; Weng, M.-w.; Wang, H.-T.; Hu, Y.; Chen, L.-C.; Park, S.-H.; Chan, H.-w.; Xu, J.; Wu, X.-R. DNA damage, DNA repair and carcinogenicity: Tobacco smoke versus electronic cigarette aerosol. Mutat. Res. Rev. Mutat. Res. 2022, 789, 108409. [Google Scholar] [CrossRef]
- Cirillo, F.; Catellani, C.; Sartori, C.; Lazzeroni, P.; Amarri, S.; Street, M.E. Obesity, insulin resistance, and colorectal cancer: Could miRNA dysregulation play a role? Int. J. Mol. Sci. 2019, 20, 2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Boakye, D.; Chen, X.; Hoffmeister, M.; Brenner, H. Association of body mass index with risk of early-onset colorectal cancer: Systematic review and meta-analysis. Am. J. Gastroenterol. 2021, 116, 2173. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Fang, W.-H.; Wang, C.-C.; Kao, T.-W.; Chang, Y.-W.; Yang, H.-F.; Wu, C.-J.; Sun, Y.-S.; Chen, W.-L. Detrimental association between betel nut chewing and colorectal polyps in adult populations. PLoS ONE 2018, 13, e0206383. [Google Scholar] [CrossRef]
- Konstandi, M.; Cheng, J.; Gonzalez, F.J. Sex steroid hormones regulate constitutive expression of Cyp2e1 in female mouse liver. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1118–E1128. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Nguyen, N.; Colditz, G.A. Links between alcohol consumption and breast cancer: A look at the evidence. Women Health 2015, 11, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Oyesanmi, O.; Snyder, D.; Sullivan, N.; Reston, J.; Treadwell, J.; Schoelles, K.M. Alcohol Consumption and Cancer Risk: Understanding Possible Causal Mechanisms for Breast and Colorectal Cancers; HRB National Drugs Library: Dublin, Ireland, 2010. [Google Scholar]



{kind=link}
{kind=link}
| CRC (n = 145) | Non-CRC (n = 1316) | p-Value | |
|---|---|---|---|
| Age, years [mean ± SE] | 58.41 ± 7.64 | 58.27 ± 7.7 | 0.835 |
| Male sex [No. (%)] | 66 (45.52) | 606 (46.05) | 0.973 |
| BMI, kg/m2 [mean ± SE] | 24.91 ± 3.10 | 25.02 ± 3.56 | 0.685 |
| Smoking [No. (%)] | 41 (28.28) | 393 (29.86) | 0.763 |
| Alcohol [No. (%)] | 15 (10.34) | 139 (10.56) | 1.000 |
| betel nut-chewing [No. (%)] | 18 (12.41) | 182 (13.83) | 0.731 |
| CCI [mean ± SE] | 1.37 ± 2.13 | 0.56 ± 0.96 | <0.001 * |
| Covariates | rs721673 (A > G) | rs721675 (A > T) | ||
|---|---|---|---|---|
| * aOR(95%C.I.) | * aOR(95%C.I.) | |||
| Age | ||||
| <50 | 2.72 | (1.89–3.91) | 1.73 | (0.51–5.85) |
| ≥50 | 1.73 | (0.51–5.85) | 2.75 | (1.81–4.15) |
| Sex | ||||
| Male | 2.64 | (1.67–4.17) | 2.27 | (1.34–3.85) |
| Female | 2.58 | (1.54–4.31) | 2.99 | (1.69–5.26) |
| BMI | ||||
| BMI < 24 | 2.59 | (1.48–4.52) | 2.37 | (1.29–4.37) |
| 24 ≤ BMI < 27 | 2.89 | (1.63–5.1) | 3.24 | (1.67–6.29) |
| BMI ≥ 27 | 2.58 | (1.3–5.1) | 2.61 | (1.18–0.01) |
| Alcohol consumption | ||||
| NO | 2.3 | (1.6–3.31) | 2.26 | (1.5–3.39) |
| Yes | 15.15 | (3.91–58.82) | 16.39 | (3.33–83.33) |
| Smoking | ||||
| NO | 2.45 | (1.64–3.66) | 2.42 | (1.54–3.83) |
| Yes | 3.3 | (1.7–6.41) | 3.32 | (1.59–6.94) |
| Betel nut-chewing habits | ||||
| NO | 2.95 | (1.74–3.61) | 2.51 | (1.66–3.8) |
| Yes | 3.53 | (1.28–9.71) | 2.95 | (0.92–9.43) |
| CCI score | ||||
| CCI = 0 | 2.15 | (1.26–3.67) | 2.06 | (1.16–3.67) |
| CCI ≥ 1 | 1.76 | (1.82–4.67) | 3.07 | (1.78–5.30) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, T.-C.; Chuang, M.-H.; Hsiung, C.-N.; Chang, P.-K.; Sun, C.-A.; Yang, T.; Chou, Y.-C.; Hu, J.-M.; Hsu, C.-H. Susceptibility to Colorectal Cancer Based on HSD17B4 rs721673 and rs721675 Polymorphisms and Alcohol Intake among Taiwan Biobank Participants: A Retrospective Case Control Study Using the Nationwide Claims Data. J. Pers. Med. 2023, 13, 576. https://doi.org/10.3390/jpm13040576
Lin T-C, Chuang M-H, Hsiung C-N, Chang P-K, Sun C-A, Yang T, Chou Y-C, Hu J-M, Hsu C-H. Susceptibility to Colorectal Cancer Based on HSD17B4 rs721673 and rs721675 Polymorphisms and Alcohol Intake among Taiwan Biobank Participants: A Retrospective Case Control Study Using the Nationwide Claims Data. Journal of Personalized Medicine. 2023; 13(4):576. https://doi.org/10.3390/jpm13040576
Chicago/Turabian StyleLin, Tzu-Chiao, Min-Hua Chuang, Chia-Ni Hsiung, Pi-Kai Chang, Chien-An Sun, Tsan Yang, Yu-Ching Chou, Je-Ming Hu, and Chih-Hsiung Hsu. 2023. "Susceptibility to Colorectal Cancer Based on HSD17B4 rs721673 and rs721675 Polymorphisms and Alcohol Intake among Taiwan Biobank Participants: A Retrospective Case Control Study Using the Nationwide Claims Data" Journal of Personalized Medicine 13, no. 4: 576. https://doi.org/10.3390/jpm13040576