
The Progress and Pitfalls of Pharmacogenetics-Based Precision Medicine in Schizophrenia Spectrum Disorders: A Systematic Review and Meta-Analysis
 ,
,  and
and 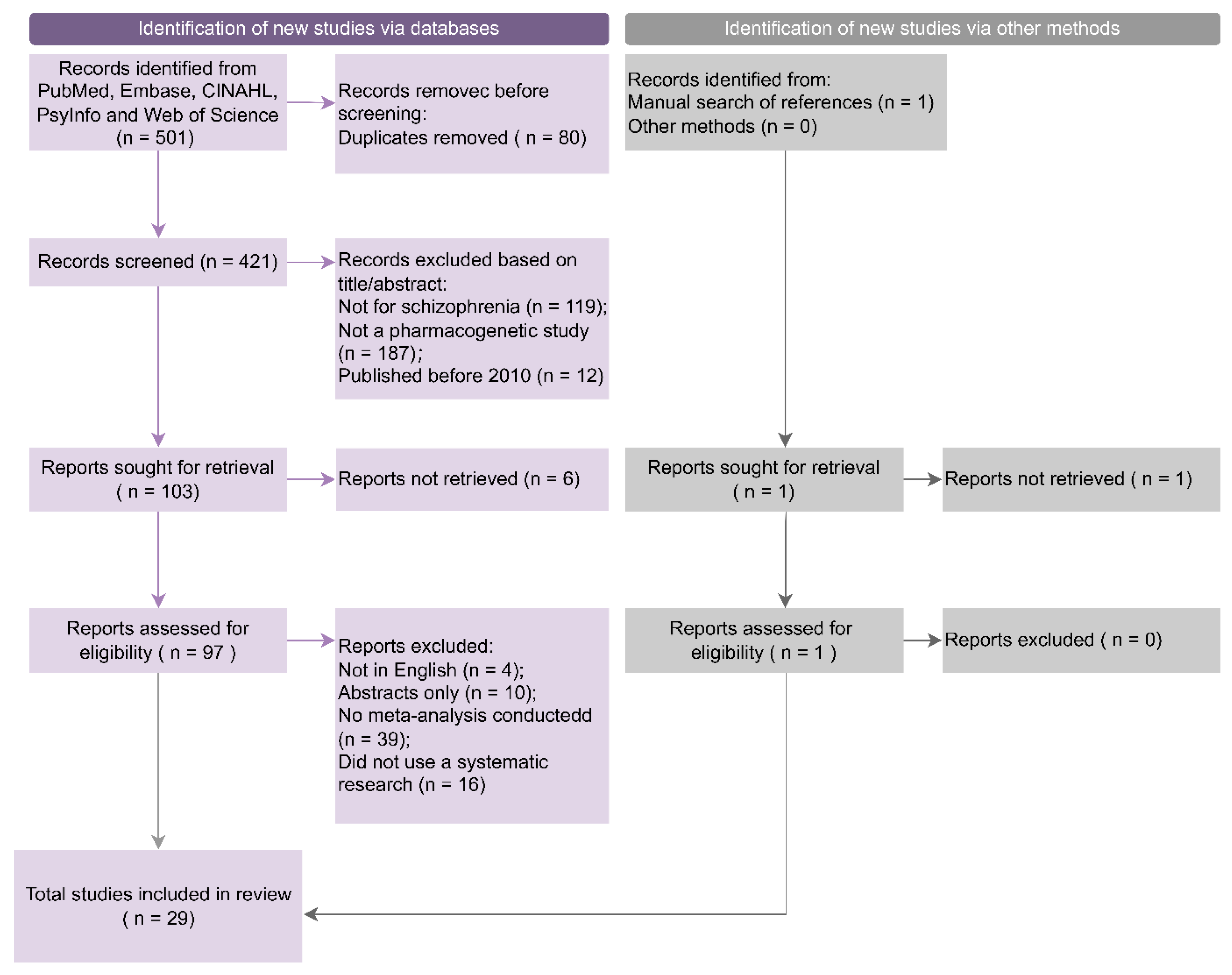{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Pharmacogenetics
1.2. Role of Pharmacogenetics in Precision Psychiatry
1.3. PGx Studies Shortcomings
2. Materials and Methods
2.1. Search Strategy and Study Selection
2.2. Outcomes
2.3. Data Extraction and Synthesis
2.4. Data Synthesis and Analysis
2.5. Additive Effects
3. Results
3.1. Study Inclusion
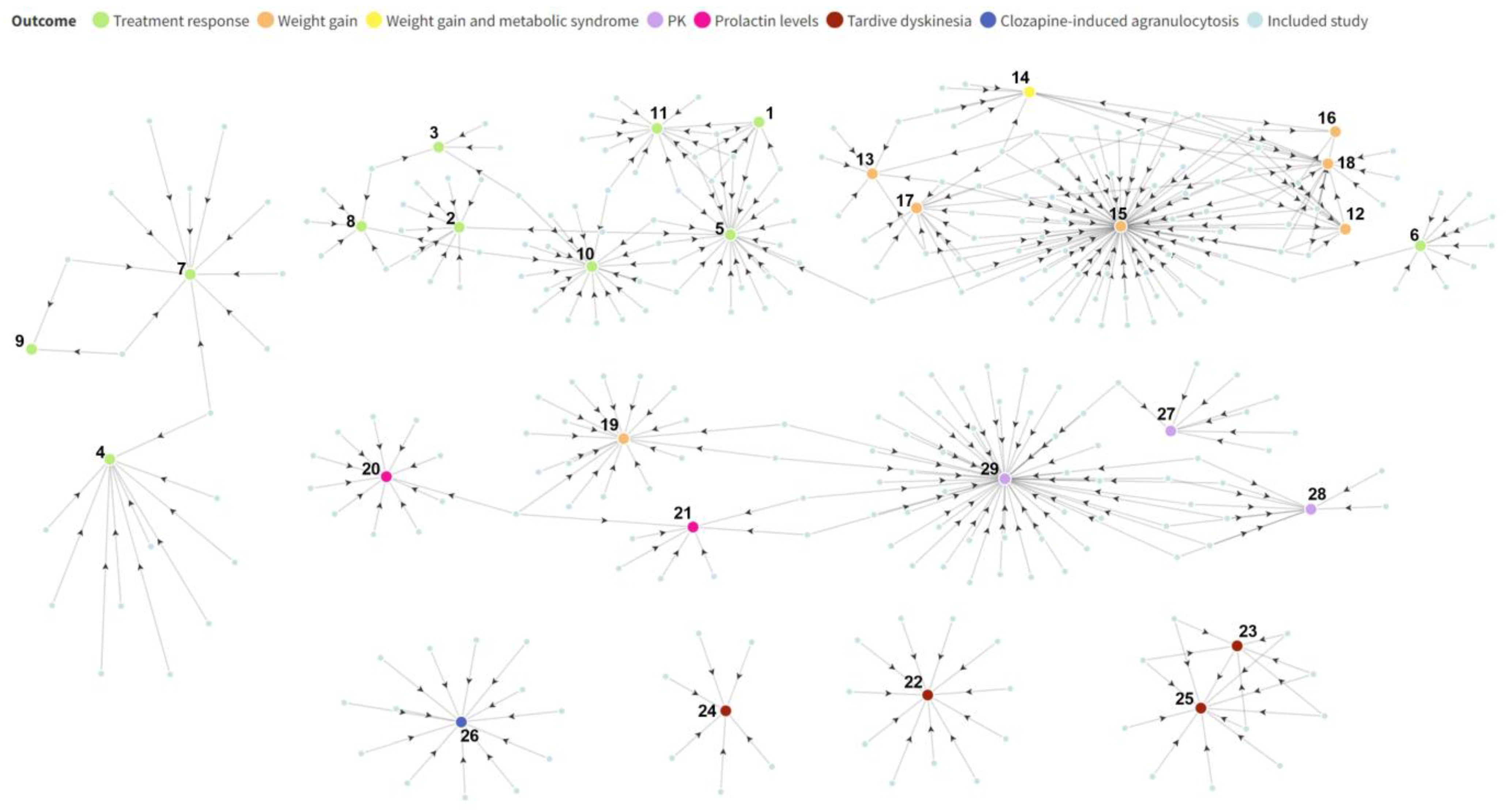
3.2. Original Research Studies Included in Meta-Analysis Studies
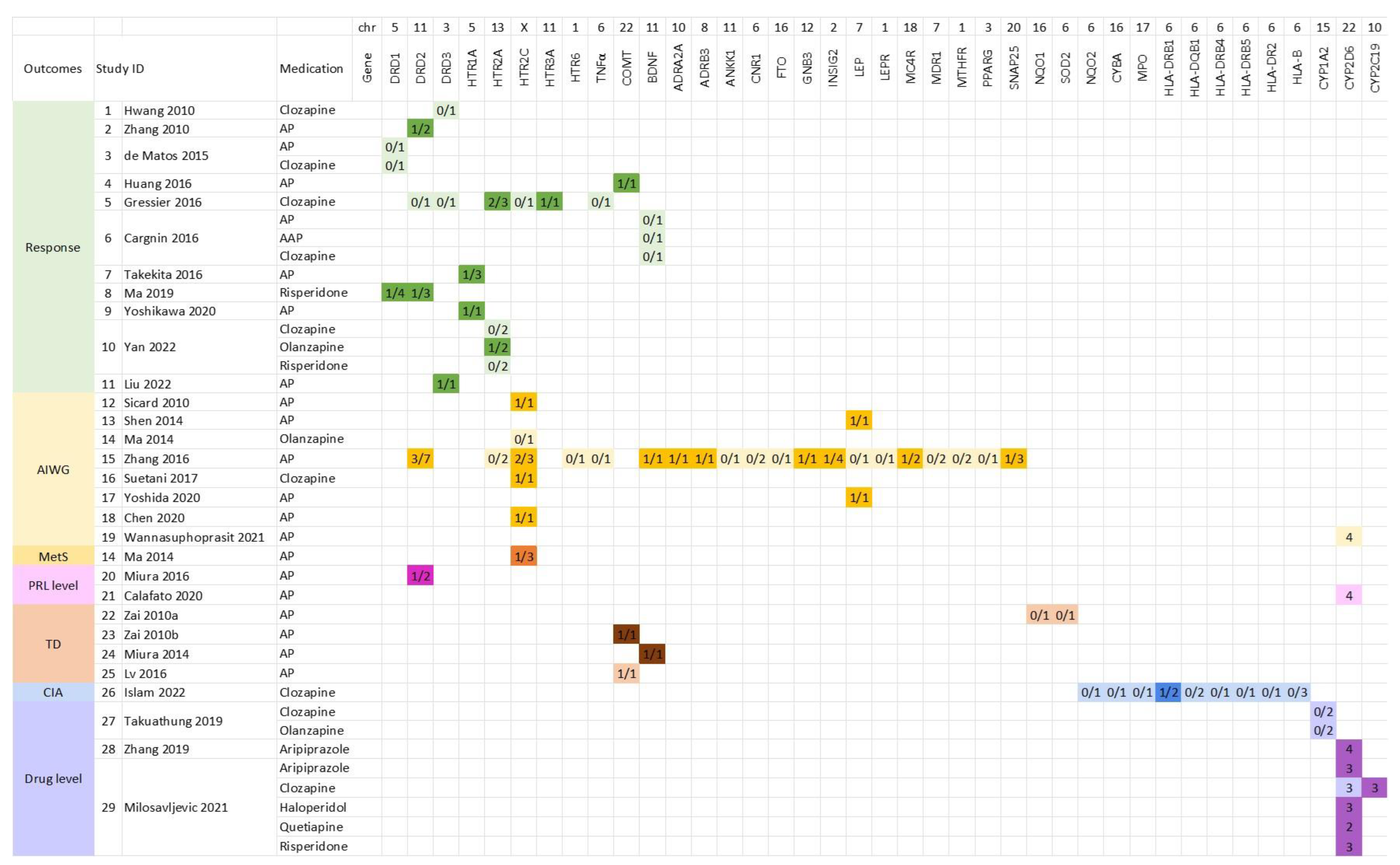
3.3. Genes Investigated in Included Meta-Analysis Studies
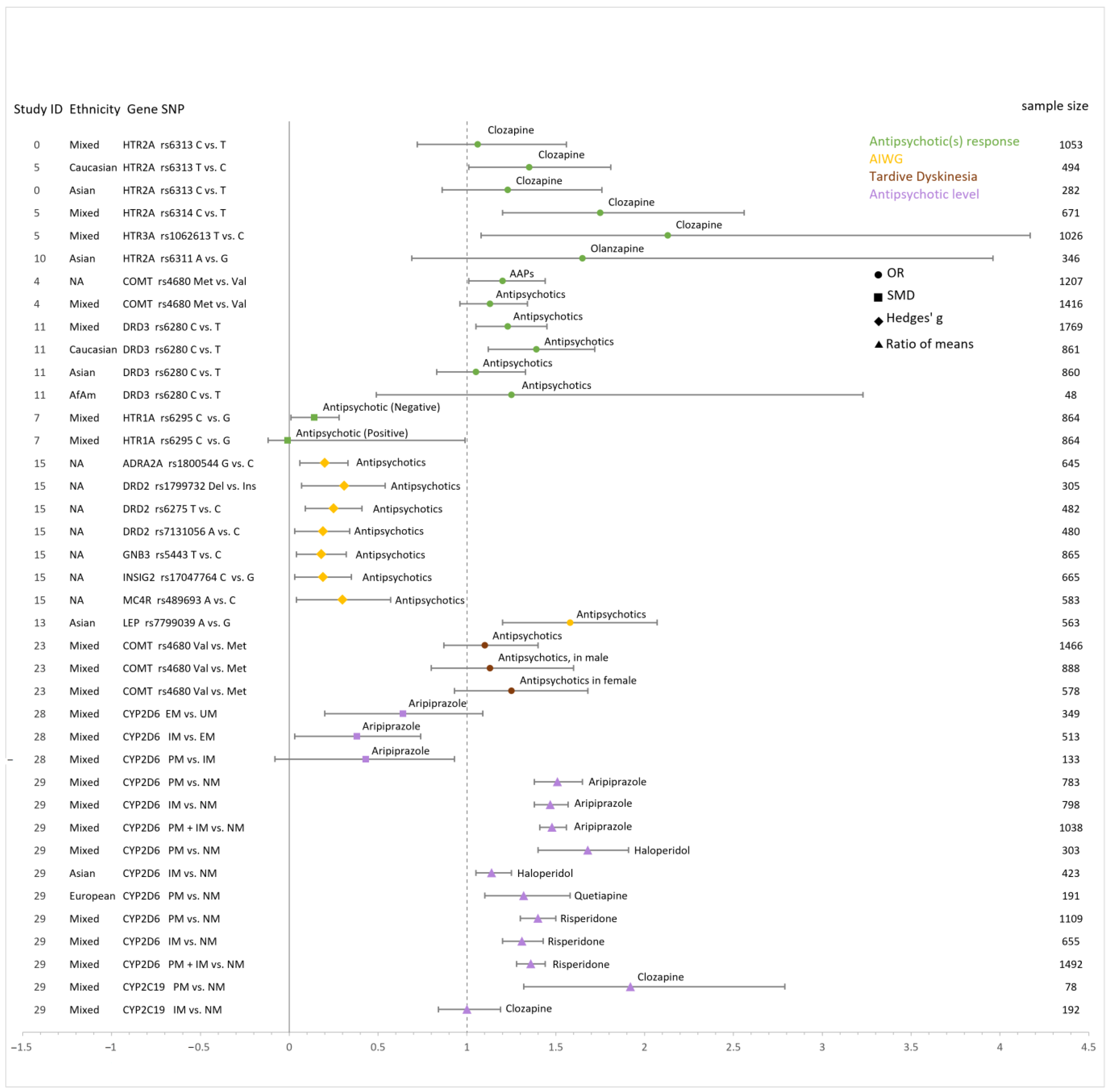
3.4. Additive Effect Sizes
3.5. Antipsychotics Response
3.5.1. HTR2A
3.5.2. DRD1, DRD2 and DRD3
3.5.3. COMT
3.5.4. HTR1A
3.6. Antipsychotic-Induced Weight Gain
3.6.1. DRD2
3.6.2. HTR2C
3.6.3. LEP
3.7. Antipsychotic-Induced Metabolic Syndrome
3.8. Antipsychotic-Related Prolactin Level
3.9. Tardive Dyskinesia
3.10. Clozapine-Induced Agranulocytosis
3.11. Pharmacokinetics of Antipsychotics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tandon, R.; Gaebel, W.; Barch, D.M.; Bustillo, J.; Gur, R.E.; Heckers, S.; Malaspina, D.; Owen, M.J.; Schultz, S.; Tsuang, M.; et al. Definition and description of schizophrenia in the DSM-5. Schizophr. Res. 2013, 150, 3–10. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Liu, Q.; Li, N.; Guo, L.; Gao, F.; Bai, L.; Gao, F.; Lyu, J. Trends in the incidence and DALYs of schizophrenia at the global, regional and national levels: Results from the Global Burden of Disease Study 2017. Epidemiol. Psychiatr. Sci. 2020, 29, e91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csernansky, J.G.; Schuchart, E.K. Relapse and rehospitalisation rates in patients with schizophrenia: Effects of second generation antipsychotics. CNS Drugs 2002, 16, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Karlsgodt, K.H.; Sun, D.; Cannon, T.D. Structural and Functional Brain Abnormalities in Schizophrenia. Curr. Dir. Psychol. Sci. 2010, 19, 226–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCutcheon, R.A.; Krystal, J.H.; Howes, O.D. Dopamine and glutamate in schizophrenia: Biology, symptoms and treatment. World Psychiatry 2020, 19, 15–33. [Google Scholar] [CrossRef] [Green Version]
- Karam, C.S.; Ballon, J.S.; Bivens, N.M.; Freyberg, Z.; Girgis, R.R.; Lizardi-Ortiz, J.E.; Markx, S.; Lieberman, J.A.; Javitch, J.A. Signaling pathways in schizophrenia: Emerging targets and therapeutic strategies. Trends Pharmacol. Sci. 2010, 31, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Emsley, R.; Chiliza, B.; Asmal, L.; Harvey, B.H. The nature of relapse in schizophrenia. BMC Psychiatry 2013, 13, 50. [Google Scholar] [CrossRef] [Green Version]
- Elkis, H.; Buckley, P.F. Buckley, Treatment-Resistant Schizophrenia. Psychiatr. Clin. 2016, 39, 239–265. [Google Scholar]
- Zipursky, R.B.; Menezes, N.M.; Streiner, D.L. Risk of symptom recurrence with medication discontinuation in first-episode psychosis: A systematic review. Schizophr. Res. 2014, 152, 408–414. [Google Scholar] [CrossRef]
- I-SHARED (Instrument for SHAREd Decision-Making in Depression). Available online: https://www.i-shared.nl/about/ (accessed on 23 February 2023).
- Simoons, M.; Ruhé, H.G.; van Roon, E.N.; Schoevers, R.A.; Bruggeman, R.; Cath, D.C.; Muis, D.; Arends, J.; Doornbos, B.; Mulder, H. Design and methods of the ‘monitoring outcomes of psychiatric pharmacotherapy’ (MOPHAR) monitoring program: A study protocol. BMC Health Serv. Res. 2019, 19, 125. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, M.A.; Armstrong, S.C.; Markman, J.D. Psychotropic Medication Monitoring: A Review. Prim. Care Companion CNS Disord. 2019, 21, 22912. [Google Scholar] [CrossRef] [PubMed]
- Scala, J.J.; Ganz, A.B.; Snyder, M.P. Precision Medicine Approaches to Mental Health Care. Physiology 2023, 38, 82–98. [Google Scholar] [CrossRef]
- Habtewold, T.D.; Rodijk, L.H.; Liemburg, E.J.; Sidorenkov, G.; Boezen, H.M.; Bruggeman, R.; Alizadeh, B.Z. A systematic review and narrative synthesis of data-driven studies in schizophrenia symptoms and cognitive deficits. Transl. Psychiatry 2020, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, B.S.; Williams, L.M.; Steiner, J.; Leboyer, M.; Carvalho, A.F.; Berk, M. The new field of ‘precision psychiatry’. BMC Med. 2017, 15, 80. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Vicente, A.E.; Lumbreras, E.; Hernández, J.M.; Martín, M.; Calles, A.; Otín, C.L.; Algarra, S.M.; Páez, D.; Taron, M. Pharmacogenetics and pharmacogenomics as tools in cancer therapy. Drug Metab. Pers. Ther. 2016, 31, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Van Schaik, R.H.; Müller, D.J.; Serretti, A.; Ingelman-Sundberg, M. Pharmacogenetics in Psychiatry: An Update on Clinical Usability. Front. Pharmacol. 2020, 11, 575540. [Google Scholar] [CrossRef]
- Rysz, J.; Franczyk, B.; Rysz-Górzyńska, M.; Gluba-Brzózka, A. Pharmacogenomics of Hypertension Treatment. Int. J. Mol. Sci. 2020, 21, 4709. [Google Scholar] [CrossRef] [PubMed]
- Murphy, L.E.; Fonseka, T.M.; Bousman, C.A.; Müller, D.J. Gene-drug pairings for antidepressants and antipsychotics: Level of evidence and clinical application. Mol. Psychiatry 2022, 27, 593–605. [Google Scholar] [CrossRef]
- Fleeman, N.; Dundar, Y.; Dickson, R.; Jorgensen, A.; Pushpakom, S.; McLeod, C.; Pirmohamed, M.; Walley, T. Cytochrome P450 testing for prescribing antipsychotics in adults with schizophrenia: Systematic review and meta-analyses. Pharm. J. 2011, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Caudle, K.E.; Sangkuhl, K.; Whirl-Carrillo, M.; Swen, J.J.; Haidar, C.E.; Klein, T.E.; Gammal, R.S.; Relling, M.V.; Scott, S.A.; Hertz, D.L.; et al. Standardizing CYP2D6 Genotype to Phenotype Translation: Consensus Recommendations from the Clinical Pharmacogenetics Implementation Consortium and Dutch Pharmacogenetics Working Group. Clin. Transl. Sci. 2020, 13, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Annotation of FDA Label for Clozapine and CYP2D6. Available online: https://www.pharmgkb.org/labelAnnotation/PA166104815 (accessed on 3 November 2022).
- Annotation of FDA Label for Aripiprazole and CYP2D6. Available online: https://www.pharmgkb.org/labelAnnotation/PA166104839 (accessed on 3 November 2022).
- Van Westrhenen, R.; Aitchison, K.J.; Ingelman-Sundberg, M.; Jukić, M.M. Pharmacogenomics of Antidepressant and Antipsychotic Treatment: How Far Have We Got and Where Are We Going? Front. Psychiatry 2020, 11, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloret-Linares, C.; Rollason, V.; Lorenzini, K.I.; Samer, C.; Daali, Y.; Gex-Fabry, M.; Aubry, J.-M.; Desmeules, J.; Besson, M. Screening for genotypic and phenotypic variations in CYP450 activity in patients with therapeutic problems in a psychiatric setting, a retrospective study. Pharmacol. Res. 2017, 118, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Gaedigk, A.; Sangkuhl, K.; Whirl-Carrillo, M.; Klein, T.; Leeder, J.S. Prediction of CYP2D6 phenotype from genotype across world populations. Genet. Med. 2017, 19, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Ellingrod, V.L.; Perry, P.J.; Ringold, J.C.; Lund, B.C.; Bever-Stille, K.; Fleming, F.; Holman, T.L.; Miller, D. Weight gain associated with the −759C/T polymorphism of the 5HT2C receptor and olanzapine. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2005, 134B, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Kuzman, M.R.; Medved, V.; Bozina, N.; Hotujac, L.; Sain, I.; Bilusic, H. The influence of 5-HT2C and MDR1 genetic polymorphisms on antipsychotic-induced weight gain in female schizophrenic patients. Psychiatry Res. 2008, 160, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Zhao, A.-L.; Zhao, J.-P.; Zhang, Y.-H.; Xue, Z.-M.; Chen, J.-D.; Chen, X.-G. Dopamine d4 receptor gene exon iii polymorphism and interindividual variation in response to clozapine. Int. J. Neurosci. 2005, 115, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Hwang, R.; Tiwari, A.K.; Zai, C.C.; Felsky, D.; Remington, E.; Wallace, T.; Tong, R.P.; Souza, R.P.; Oh, G.; Potkin, S.G.; et al. Dopamine D4 and D5 receptor gene variant effects on clozapine response in schizophrenia: Replication and exploration. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 37, 62–75. [Google Scholar] [CrossRef]
- Gressier, F.; Porcelli, S.; Calati, R.; Serretti, A. Pharmacogenetics of clozapine response and induced weight gain: A comprehensive review and meta-analysis. Eur. Neuropsychopharmacol. 2016, 26, 163–185. [Google Scholar] [CrossRef]
- Zhang, J.-P.; Lencz, T.; Zhang, R.X.; Nitta, M.; Maayan, L.; John, M.; Robinson, D.G.; Fleischhacker, W.W.; Kahn, R.S.; Ophoff, R.A.; et al. Pharmacogenetic Associations of Antipsychotic Drug-Related Weight Gain: A Systematic Review and Meta-analysis. Schizophr. Bull. 2016, 42, 1418–1437. [Google Scholar] [CrossRef] [Green Version]
- Jarvik, G.P.; Evans, J.P. Mastering genomic terminology. Genet. Med. 2017, 19, 491–492. [Google Scholar] [CrossRef] [Green Version]
- Hwang, R.; Zai, C.; Tiwari, A.; Müller, D.J.; Arranz, M.J.; Morris, A.G.; McKenna, P.; Munro, J.; Potkin, S.G.; Lieberman, J.A.; et al. Effect of dopamine D3 receptor gene polymorphisms and clozapine treatment response: Exploratory analysis of nine polymorphisms and meta-analysis of the Ser9Gly variant. Pharm. J. 2010, 10, 200–218. [Google Scholar] [CrossRef] [Green Version]
- Sicard, M.N.; Zai, C.C.; Tiwari, A.K.; Souza, R.P.; Meltzer, H.Y.; Lieberman, J.A.; Kennedy, J.L.; Müller, D.J. Polymorphisms of the HTR2C gene and antipsychotic-induced weight gain: An update and meta-analysis. Pharmacogenomics 2010, 11, 1561–1571. [Google Scholar] [CrossRef] [PubMed]
- Lisoway, A.J.; Chen, C.C.; Zai, C.C.; Tiwari, A.K.; Kennedy, J.L. Toward personalized medicine in schizophrenia: Genetics and epigenetics of antipsychotic treatment. Schizophr. Res. 2021, 232, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Brandl, E.J.; Kennedy, J.L.; Müller, D.J. Pharmacogenetics of antipsychotics. Can. J. Psychiatry 2014, 59, 76–88. [Google Scholar] [CrossRef] [Green Version]
- Yepes-Nuñez, J.J.; Urrutia, G.; Romero-Garcia, M.; Alonso-Fernandez, S. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Rev. Esp. Cardiol. 2021, 74, 790–799. [Google Scholar]
- Martorell-Marugan, J.; Toro-Dominguez, D.; Alarcon-Riquelme, M.E.; Carmona-Saez, P. MetaGenyo: A web tool for meta-analysis of genetic association studies. BMC Bioinform. 2017, 18, 1–6. [Google Scholar] [CrossRef]
- Cargnin, S.; Massarotti, A.; Terrazzino, S. BDNF Val66Met and clinical response to antipsychotic drugs: A systematic review and meta-analysis. Eur. Psychiatry 2016, 33, 45–53. [Google Scholar] [CrossRef]
- Yan, P.; Gao, B.; Wang, S.; Wang, S.; Li, J.; Song, M. Association of 5-HTR2A T102C and A-1438G polymorphisms with clinical response to atypical antipsychotic treatment in schizophrenia: A meta-analysis. Neurosci. Lett. 2022, 770, 136395. [Google Scholar] [CrossRef]
- Liu, C.; Xu, X.; Liu, X.; Zhang, T.; Li, Y.; Yan, P. DRD3 Ser9Gly polymorphism and treatment response to antipsychotics in schizophrenia: A meta-analysis. Neurosci. Lett. 2022, 786. [Google Scholar] [CrossRef]
- Huang, E.; Zai, C.C.; Lisoway, A.; Maciukiewicz, M.; Felsky, D.; Tiwari, A.K.; Bishop, J.R.; Ikeda, M.; Molero, P.; Ortuño, F.; et al. Catechol-O-Methyltransferase Val158Met Polymorphism and Clinical Response to Antipsychotic Treatment in Schizophrenia and Schizo-Affective Disorder Patients: A Meta-Analysis. Int. J. Neuropsychopharmacol. 2016, 19, pyv132. [Google Scholar] [CrossRef] [Green Version]
- Takekita, Y.; Fabbri, C.; Kato, M.; Koshikawa, Y.; Tajika, A.; Kinoshita, T.; Serretti, A. HTR1A Polymorphisms and Clinical Efficacy of Antipsychotic Drug Treatment in Schizophrenia: A Meta-Analysis. Int. J. Neuropsychopharmacol. 2016, 19. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, A.; Li, J.; Meltzer, H.Y. A functional HTR1A polymorphism, rs6295, predicts short-term response to lurasidone: Confirmation with meta-analysis of other antipsychotic drugs. Pharm. J. 2020, 20, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-P.; Lencz, T.; Malhotra, A.K.; Robinson, D.; Yu, J.; Gallego, J.; Fleischhacker, W.W.; Kahn, R.S.; Crespo-Facorro, B.; Vazquez-Bourgon, J.; et al. D2Receptor Genetic Variation and Clinical Response to Antipsychotic Drug Treatment: A Meta-Analysis. Am. J. Psychiatry 2010, 167, 763–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Matos, L.P.; Santana, C.V.; Souza, R.P. Meta-analysis of dopamine receptor D1 rs4532 polymorphism and susceptibility to antipsychotic treatment response. Psychiatry Res. 2015, 229, 586–588. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, X.; Xiang, Q.; Zhou, S.; Zhao, N.; Xie, Q.; Zhao, X.; Zhou, Y.; Cui, Y. Association between dopamine receptor gene polymorphisms and effects of risperidone treatment: A systematic review and meta-analysis. Basic Clin. Pharmacol. Toxicol. 2019, 124, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Maimaitirexiati, T.; Zhang, R.; Gui, X.; Zhang, W.; Xu, G.; Hu, G. HTR2Cpolymorphisms, olanzapine-induced weight gain and antipsychotic-induced metabolic syndrome in schizophrenia patients: A meta-analysis. Int. J. Psychiatry Clin. Pr. 2014, 18, 229–242. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Fang, X.; Zhang, Y.; Song, L.; Zhang, C. Association of the HTR2C-759C/T polymorphism and antipsychotic-induced weight gain: A meta-analysis. Gen. Psychiatry 2020, 33, e100192. [Google Scholar] [CrossRef]
- Shen, J.; Ge, W.; Zhang, J.; Zhu, H.J.; Fang, Y. Leptin -2548g/a gene polymorphism in association with antipsychotic-induced weight gain: A meta-analysis study. Psychiatr. Danub. 2014, 26. [Google Scholar]
- Yoshida, K.; Maciukiewicz, M.; Zai, C.C.; Gonçalves, V.F.; Brandl, E.J.; Lieberman, J.A.; Meltzer, H.Y.; Tiwari, A.K.; Kennedy, J.L.; Müller, D.J. Association between the -2548G/A polymorphism of the leptin gene and antipsychotic-induced weight gain: Analysis of the CATIE sample and meta-analysis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 102, 109952. [Google Scholar] [CrossRef]
- Suetani, R.J.; Siskind, D.; Reichhold, H.; Kisely, S. Genetic variants impacting metabolic outcomes among people on clozapine: A systematic review and meta-analysis. Psychopharmacology 2017, 234, 2989–3008. [Google Scholar] [CrossRef] [PubMed]
- Wannasuphoprasit, Y.; Andersen, S.E.; Arranz, M.J.; Catalan, R.; Jurgens, G.; Kloosterboer, S.M.; Rasmussen, H.B.; Bhat, A.; Irizar, H.; Koller, D.; et al. CYP2D6 Genetic Variation and Antipsychotic-Induced Weight Gain: A Systematic Review and Meta-Analysis. Front. Psychol. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Miura, I.; Zhang, J.-P.; Hagi, K.; Lencz, T.; Kane, J.M.; Yabe, H.; Malhotra, A.K.; Correll, C.U. Variants in the DRD2 locus and antipsychotic-related prolactin levels: A meta-analysis. Psychoneuroendocrinology 2016, 72, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Calafato, M.S.; Austin-Zimmerman, I.; Thygesen, J.H.; Sairam, M.; Metastasio, A.; Marston, L.; Abad-Santos, F.; Bhat, A.; Harju-Seppänen, J.; Irizar, H.; et al. The effect of CYP2D6 variation on antipsychotic-induced hyperprolactinaemia: A systematic review and meta-analysis. Pharm. J. 2020, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Miura, I.; Zhang, J.-P.; Nitta, M.; Lencz, T.; Kane, J.M.; Malhotra, A.K.; Yabe, H.; Correll, C.U. BDNF Val66Met polymorphism and antipsychotic-induced tardive dyskinesia occurrence and severity: A meta-analysis. Schizophr. Res. 2014, 152, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.; Rong, B.; Tong, X.; Li, X.; Chen, X.; Wang, X.; Li, Z. The association between COMT Val158Met gene polymorphism and antipsychotic-induced tardive dyskinesia risk. Int. J. Neurosci. 2016, 126, 1044–1050. [Google Scholar] [CrossRef]
- Zai, C.C.; Tiwari, A.K.; Basile, V.; de Luca, V.; Müller, D.J.; Voineskos, A.N.; Remington, G.; Meltzer, H.Y.; Lieberman, J.A.; Potkin, S.G.; et al. Oxidative stress in tardive dyskinesia: Genetic association study and meta-analysis of NADPH quinine oxidoreductase 1 (NQO1) and Superoxide dismutase 2 (SOD2, MnSOD) genes. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 34, 50–56. [Google Scholar] [CrossRef]
- Zai, C.C.; Tiwari, A.K.; Müller, D.J.; de Luca, V.; Shinkai, T.; Shaikh, S.; Ni, X.; Sibony, D.; Voineskos, A.N.; Meltzer, H.Y.; et al. The catechol-O-methyl-transferase gene in tardive dyskinesia. World J. Biol. Psychiatry 2010, 11, 803–812. [Google Scholar] [CrossRef]
- Islam, F.; Hain, D.; Lewis, D.; Law, R.; Brown, L.C.; Tanner, J.-A.; Müller, D.J. Pharmacogenomics of Clozapine-induced agranulocytosis: A systematic review and meta-analysis. Pharm. J. 2022, 22, 230–240. [Google Scholar] [CrossRef]
- Milosavljević, F.; Bukvić, N.; Pavlović, Z.; Miljević, Č.; Pešić, V.; Molden, E.; Ingelman-Sundberg, M.; Leucht, S.; Jukić, M.M. Association of CYP2C19 and CYP2D6 Poor and Intermediate Metabolizer Status With Antidepressant and Antipsychotic Exposure A Systematic Review and Meta-analysis. JAMA Psychiatry 2021, 78, 270–280. [Google Scholar] [CrossRef]
- Zhang, X.; Xiang, Q.; Zhao, X.; Ma, L.; Cui, Y. Association between aripiprazole pharmacokinetics and CYP2D6 phenotypes: A systematic review and meta-analysis. J. Clin. Pharm. Ther. 2019, 44, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Na Takuathung, M.; Hanprasertpong, N.; Teekachunhatean, S.; Koonrungsesomboon, N. Impact of CYP1A2 genetic polymorphisms on pharmacokinetics of antipsychotic drugs: A systematic review and meta-analysis. Acta Psychiatr. Scand. 2019, 139, 15–25. [Google Scholar] [CrossRef]
- Zai, G.C.; Zai, C.C.; Chowdhury, N.I.; Tiwari, A.K.; Souza, R.P.; Lieberman, J.A.; Meltzer, H.Y.; Potkin, S.G.; Müller, D.J.; Kennedy, J.L. The role of brain-derived neurotrophic factor (BDNF) gene variants in antipsychotic response and antipsychotic-induced weight gain. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Auwerx, C.; Sadler, M.C.; Reymond, A.; Kutalik, Z. From pharmacogenetics to pharmaco-omics: Milestones and future directions. Hum. Genet. Genom. Adv. 2022, 3, 100100. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ingelman-Sundberg, M.; Lauschke, V. Worldwide Distribution of Cytochrome P450 Alleles: A Meta-analysis of Population-scale Sequencing Projects. Clin. Pharmacol. Ther. 2017, 102, 688–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maple, A.M.; Zhao, X.; Elizalde, D.I.; McBride, A.K.; Gallitano, A.L. Htr2a Expression Responds Rapidly to Environmental Stimuli in an Egr3-Dependent Manner. ACS Chem. Neurosci. 2015, 6, 1137–1142. [Google Scholar] [CrossRef] [Green Version]
- Trubetskoy, V.; Pardiñas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.-Y.; Dennison, C.A.; Hall, L.S.; et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, K.M.; Yee, S.W.; Mushiroda, T.; Weinshilboum, R.M.; Ratain, M.J.; Kubo, M. Genome-wide association studies of drug response and toxicity: An opportunity for genome medicine. Nat. Rev. Drug Discov. 2017, 16, 70. [Google Scholar] [CrossRef] [Green Version]
- The International Schizophrenia Consortium. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 2009, 460, 748–752. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; Wilke, M.A.; Lyle, S.M.; Kowalec, K.; Jorgensen, A.; Wright, G.E.; Drögemöller, B.I. A Systematic Review and Analysis of the Use of Polygenic Scores in Pharmacogenomics. Clin. Pharmacol. Ther. 2022, 111, 919–930. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teng, Y.; Sandhu, A.; Liemburg, E.J.; Naderi, E.; Alizadeh, B.Z. The Progress and Pitfalls of Pharmacogenetics-Based Precision Medicine in Schizophrenia Spectrum Disorders: A Systematic Review and Meta-Analysis. J. Pers. Med. 2023, 13, 471. https://doi.org/10.3390/jpm13030471
Teng Y, Sandhu A, Liemburg EJ, Naderi E, Alizadeh BZ. The Progress and Pitfalls of Pharmacogenetics-Based Precision Medicine in Schizophrenia Spectrum Disorders: A Systematic Review and Meta-Analysis. Journal of Personalized Medicine. 2023; 13(3):471. https://doi.org/10.3390/jpm13030471
Chicago/Turabian StyleTeng, Yuxin, Amrit Sandhu, Edith J. Liemburg, Elnaz Naderi, and Behrooz Z. Alizadeh. 2023. "The Progress and Pitfalls of Pharmacogenetics-Based Precision Medicine in Schizophrenia Spectrum Disorders: A Systematic Review and Meta-Analysis" Journal of Personalized Medicine 13, no. 3: 471. https://doi.org/10.3390/jpm13030471