

CD169+ Monocyte and Regulatory T Cell Subsets Are Associated with Disease Activity in Rheumatoid Arthritis
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Blood Processing
2.3. Cell Immunophenotyping
2.4. Cell Expansion and Culture
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Statistical Analysis
3. Results
3.1. Study Participant Demographics
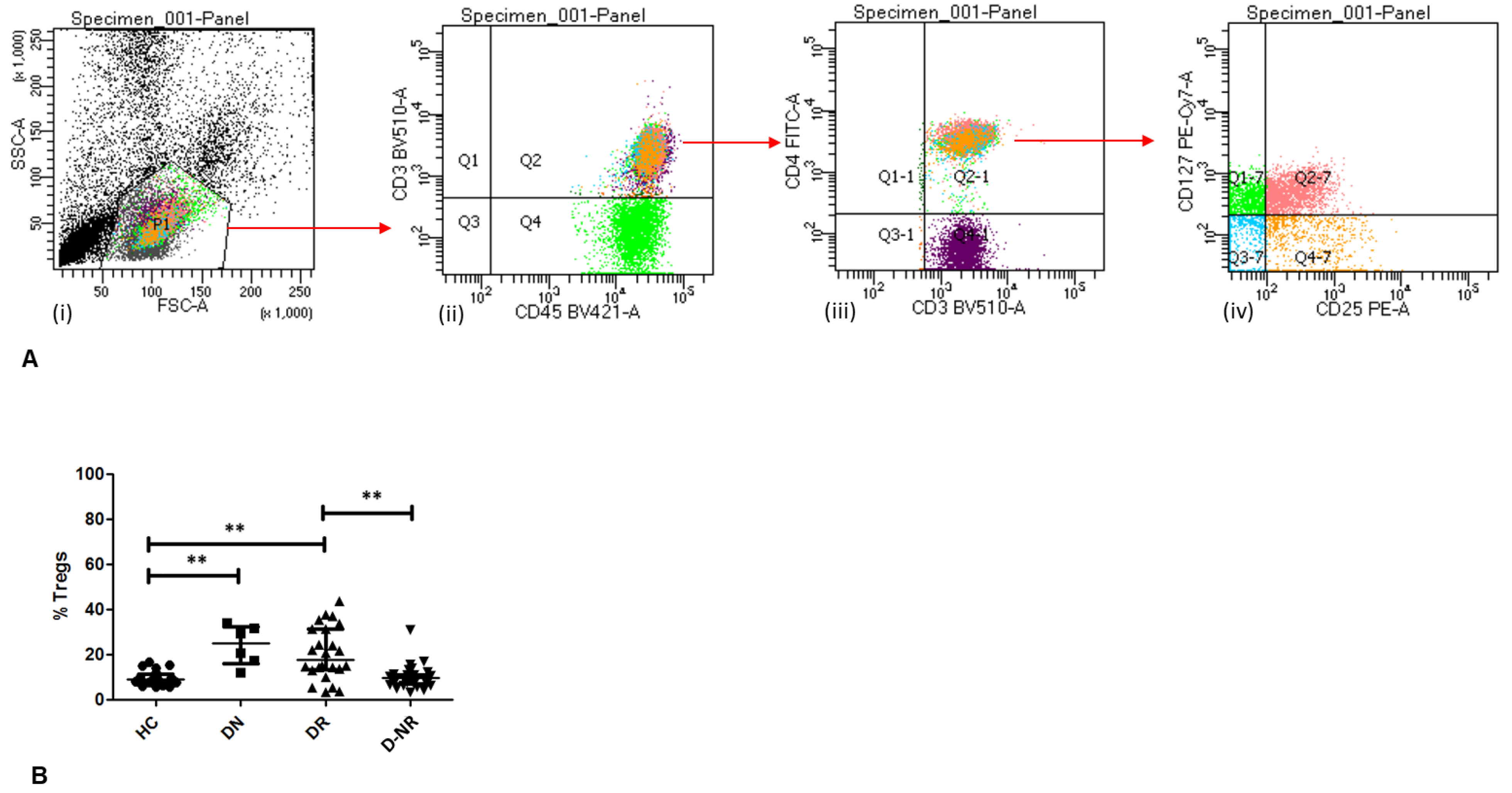
3.2. Frequency of Circulating Tregs in RA
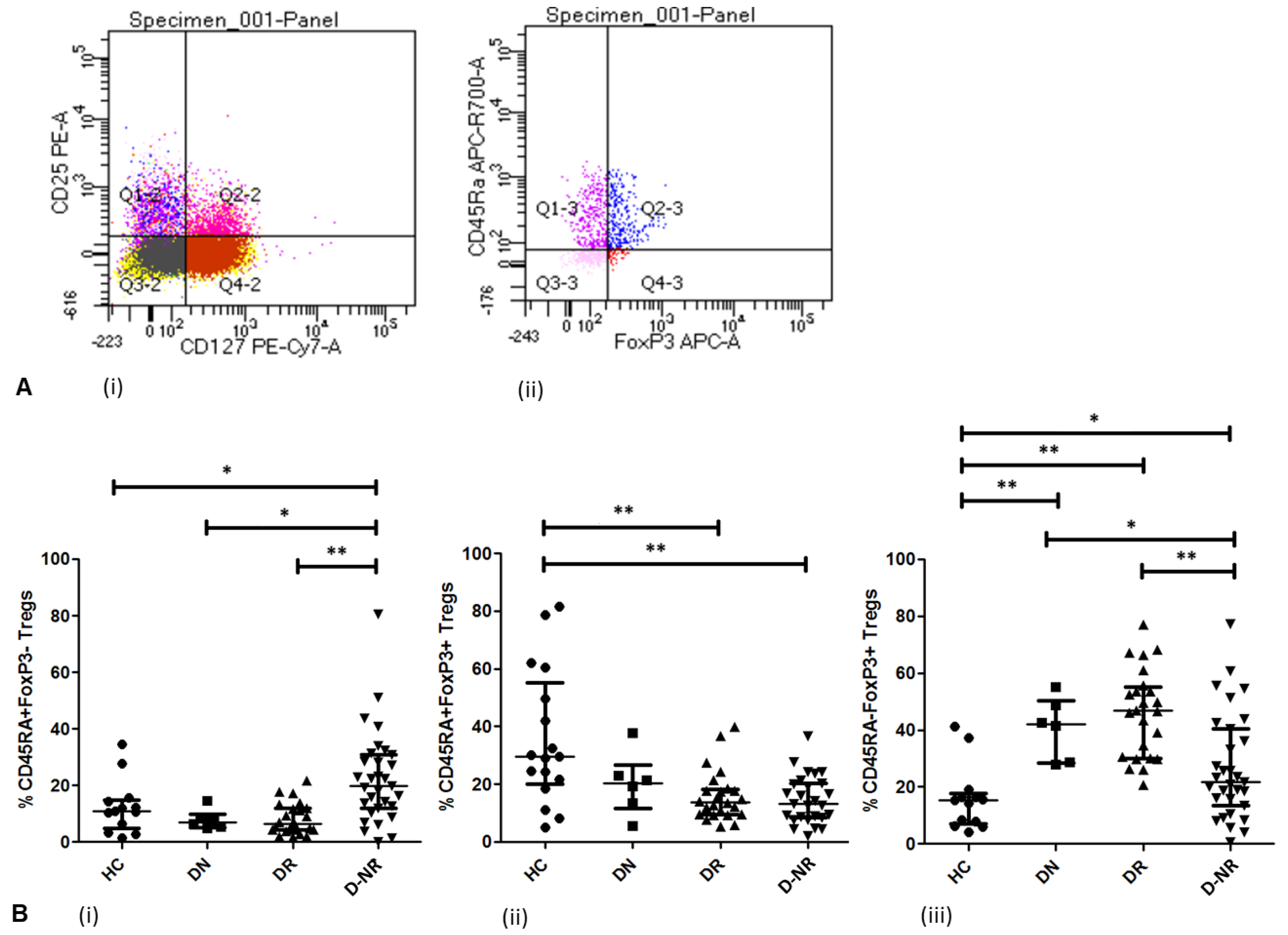
3.3. Treg Activation in RA (FoxP3 and CD45RA)
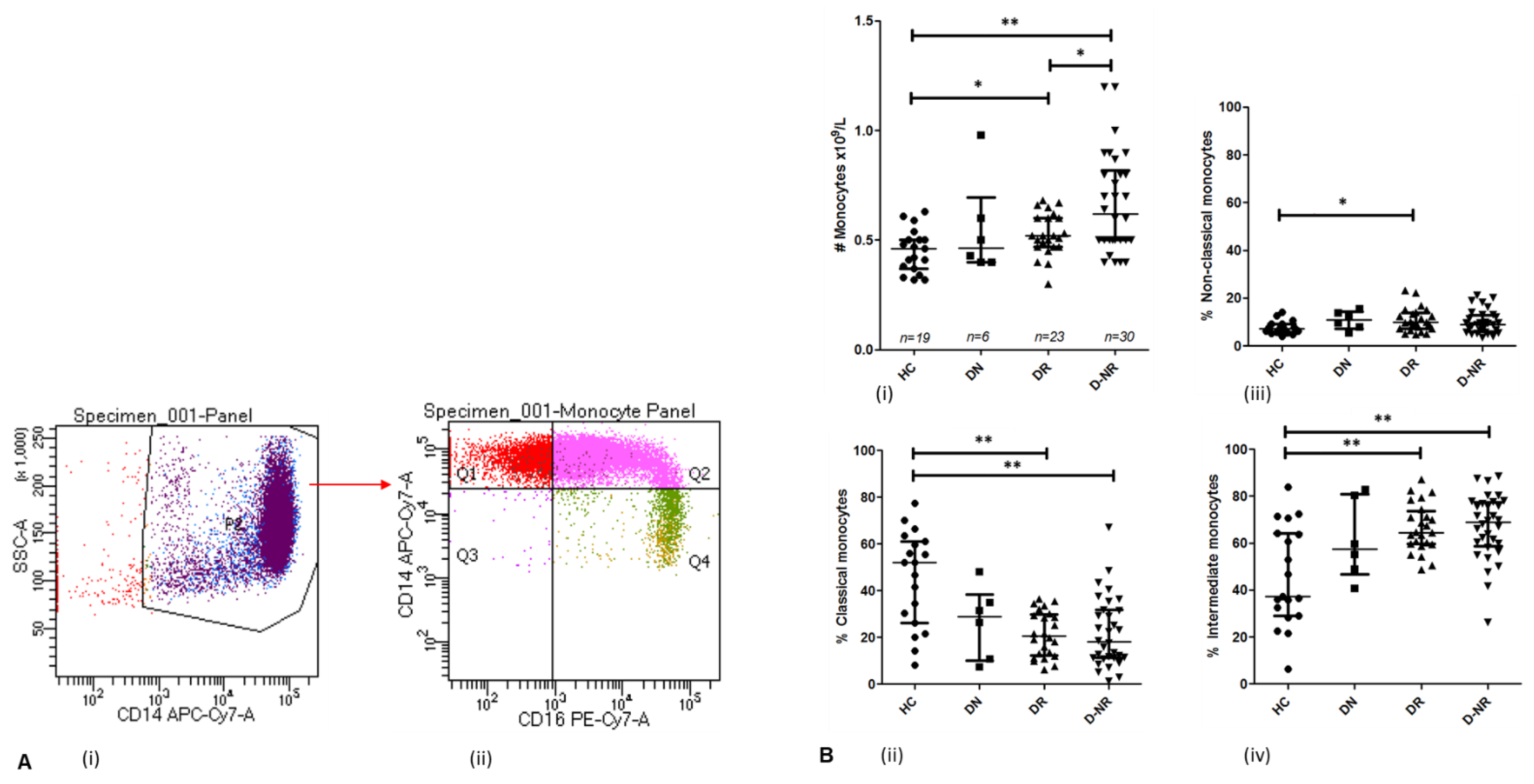
3.4. Monocytes in Health and RA Subgroups
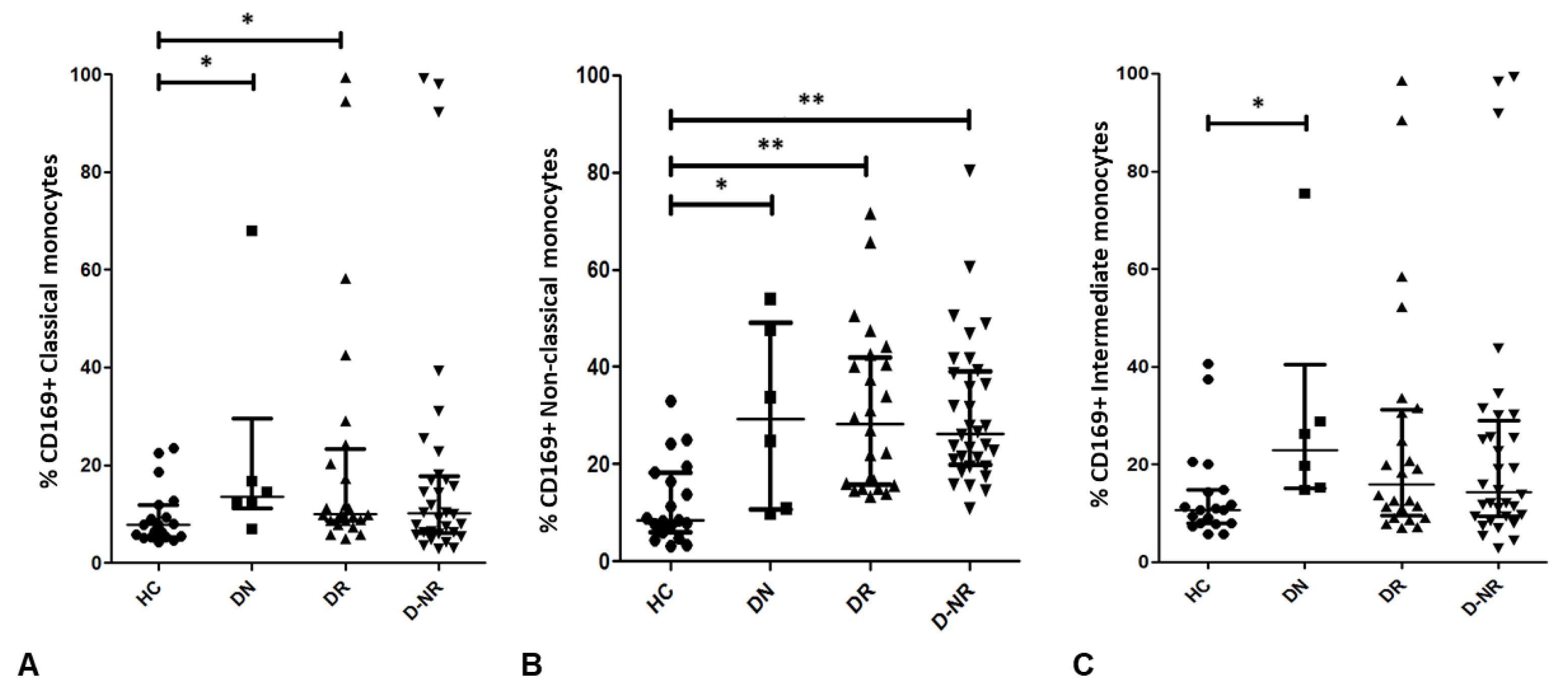
3.5. CD169 Expression on Monocyte Subsets in RA and Health
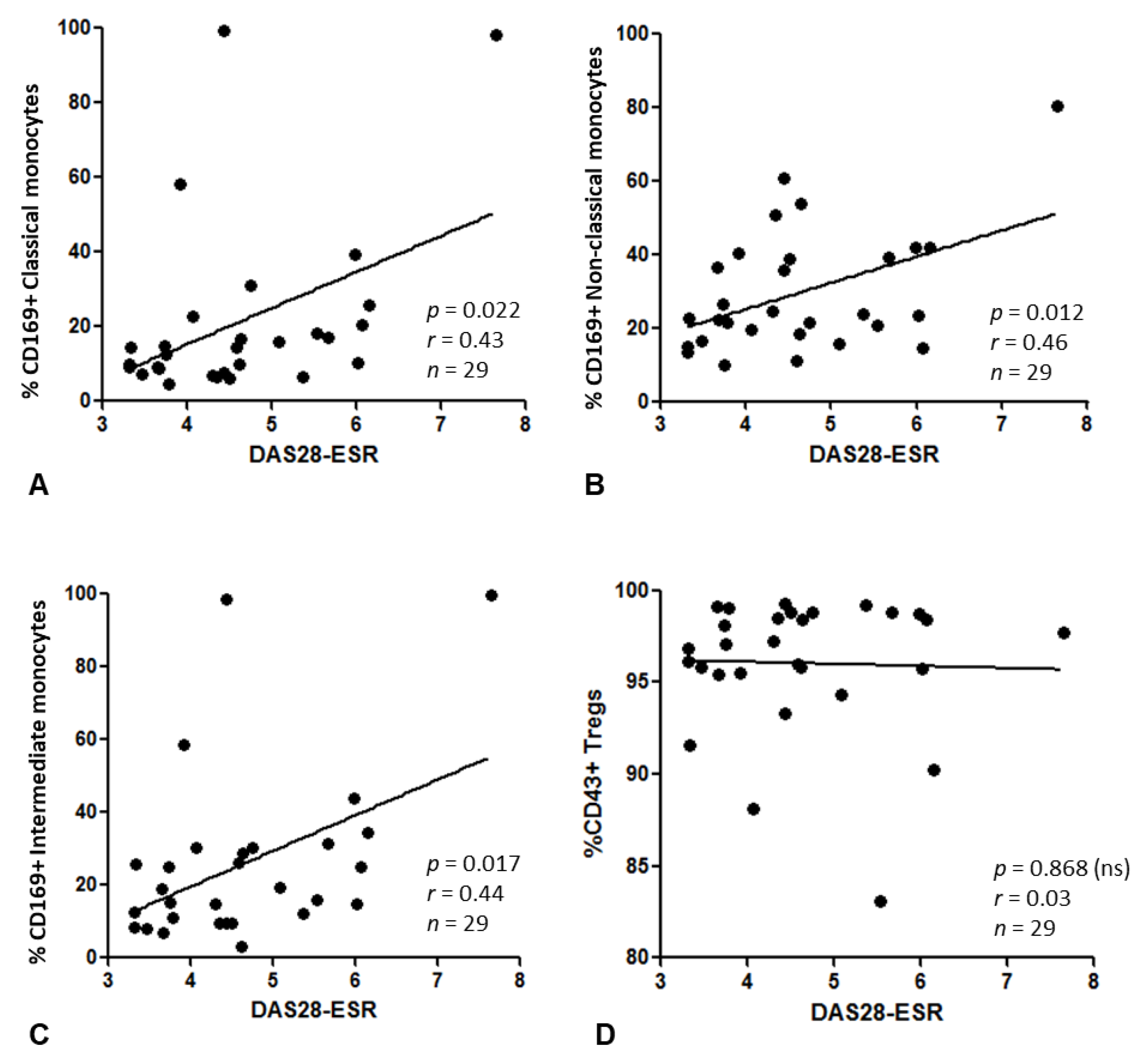
3.6. Relationship between CD169+ Monocytes or CD43+ Tregs and Disease Activity
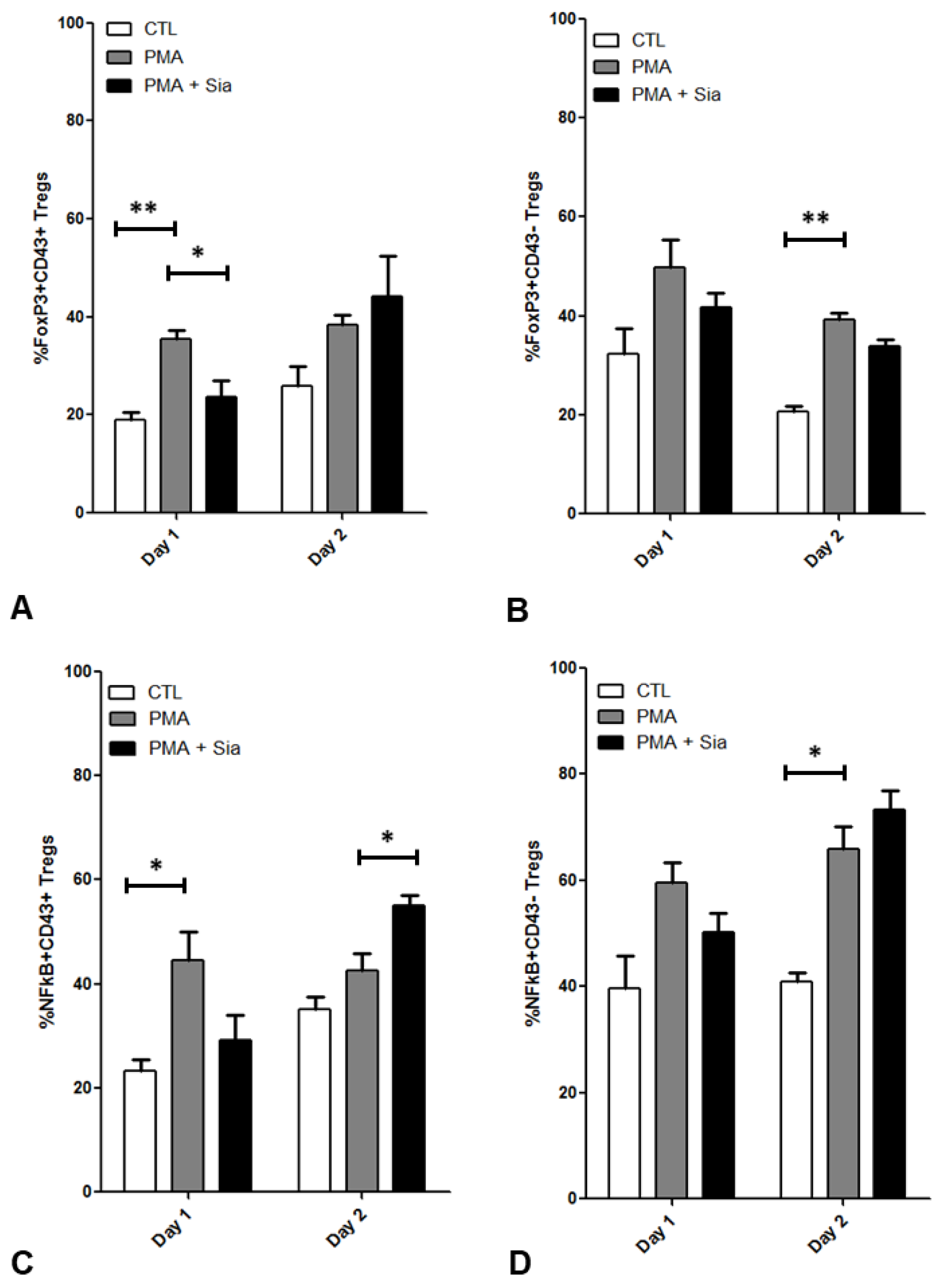
3.7. Effect of Sialic Acid on PMA Stimulated Tregs
4. Discussions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheehan, N.J.; Slavin, B.M.; Donovan, M.P.; Mount, J.N.; Mathews, J.A. Lack of correlation between clinical disease activity and erythrocyte sedimentation rate, acute phase proteins or protease inhibitors in ankylosing spondylitis. Br. J. Rheumatol. 1986, 25, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Ruof, J.; Stucki, G. Validity aspects of erythrocyte sedimentation rate and C-reactive protein in ankylosing spondylitis: A literature review. J. Rheumatol. 1999, 26, 966–970. [Google Scholar] [PubMed]
- Baum, M.K.; Rafie, C.; Sales, S.; Lai, S.; Duan, R.; Jayaweera, D.T.; Page, J.B.; Campa, A. C-reactive protein: A poor marker of cardiovascular disease risk in HIV+ populations with a high prevalence of elevated serum transaminases. Int. J. STD AIDS 2008, 19, 410–413. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S.M.; Kim, K.W.; Yang, C.W.; Park, S.H.; Ju, J.H. Cytokine-mediated bone destruction in rheumatoid arthritis. J. Immunol. Res. 2014, 2014, 263625. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.C.; Manning, H.B.; Jain, A.; Troeberg, L.; Dudhia, J.; Essex, D.; Sandison, A.; Seiki, M.; Nanchahal, J.; Nagase, H.; et al. Membrane type 1 matrix metalloproteinase is a crucial promoter of synovial invasion in human rheumatoid arthritis. Arthritis Rheum. 2009, 60, 686–697. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.; Egan, P.; Guo, G.; Kelly, C.; McClean, P.; McGilligan, V.E.; Mc Allister, R.; Matchett, K.; Martin, C.; Murray, E.K.; et al. Precision medicine to manage chronic immune-related conditions. Transl. Autoimmun. 2022, 2, 295–318. [Google Scholar] [CrossRef]
- van Amelsfort, J.M.; Jacobs, K.M.; Bijlsma, J.W.; Lafeber, F.P.; Taams, L.S. CD4+CD25+ regulatory T cells in rheumatoid arthritis: Differences in the presence, phenotype, and function between peripheral blood and synovial fluid. Arthritis Rheum. 2004, 50, 2775–2785. [Google Scholar] [CrossRef]
- Ehrenstein, M.R.; Evans, J.G.; Singh, A.; Moore, S.; Warnes, G.; Isenberg, D.A.; Mauri, C. Compromised function of regulatory T cells in rheumatoid arthritis and reversal by anti-TNFalpha therapy. J. Exp. Med. 2004, 200, 277–285. [Google Scholar] [CrossRef]
- Mottonen, M.; Heikkinen, J.; Mustonen, L.; Isomaki, P.; Luukkainen, R.; Lassila, O. CD4+ CD25+ T cells with the phenotypic and functional characteristics of regulatory T cells are enriched in the synovial fluid of patients with rheumatoid arthritis. Clin. Exp. Immunol. 2005, 140, 360–367. [Google Scholar] [CrossRef]
- Bonelli, M.; Goschl, L.; Bluml, S.; Karonitsch, T.; Hirahara, K.; Ferner, E.; Steiner, C.W.; Steiner, G.; Smolen, J.S.; Scheinecker, C. Abatacept (CTLA-4Ig) treatment reduces T cell apoptosis and regulatory T cell suppression in patients with rheumatoid arthritis. Rheumatology 2016, 55, 710–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, J.E.; Edwards, J.C.; Blades, S.; Goulding, N.J. Macrophage subpopulations in rheumatoid synovium: Reduced CD163 expression in CD4+ T lymphocyte-rich microenvironments. Arthritis Rheum. 2002, 46, 1210–1216. [Google Scholar] [CrossRef]
- Yoon, B.R.; Yoo, S.J.; Choi, Y.; Chung, Y.H.; Kim, J.; Yoo, I.S.; Kang, S.W.; Lee, W.W. Functional phenotype of synovial monocytes modulating inflammatory T-cell responses in rheumatoid arthritis (RA). PLoS ONE 2014, 9, e109775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, G.J.; Evans, H.G.; Menon, B.; Gullick, N.J.; Kirkham, B.W.; Cope, A.P.; Geissmann, F.; Taams, L.S. Interaction with activated monocytes enhances cytokine expression and suppressive activity of human CD4+CD45ro+CD25+CD127low regulatory T cells. Arthritis Rheum. 2013, 65, 627–638. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Capetillo, L.; Hernandez-Castro, B.; Monsivais-Urenda, A.; Alvarez-Quiroga, C.; Layseca-Espinosa, E.; Abud-Mendoza, C.; Baranda, L.; Urzainqui, A.; Sánchez-Madrid, F.; González-Amaro, R. Induction of Th17 lymphocytes and Treg cells by monocyte-derived dendritic cells in patients with rheumatoid arthritis and systemic lupus erythematosus. Clin. Dev. Immunol. 2013, 2013, 584303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartnell, A.; Steel, J.; Turley, H.; Jones, M.; Jackson, D.G.; Crocker, P.R. Characterization of human sialoadhesin, a sialic acid binding receptor expressed by resident and inflammatory macrophage populations. Blood 2001, 97, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.S.; Cheng, Y.; Lin, Q.S.; Wu, A.L.; Yu, J.; Li, C.; Sun, Y.; Zhong, R.Q.; Wu, L.J. Increased expression of Siglec-1 on peripheral blood monocytes and its role in mononuclear cell reactivity to autoantigen in rheumatoid arthritis. Rheumatology 2014, 53, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Eakin, A.; Bustard, M.; McGeough, C.; Ahmed, T.; Bjourson, A.J.; Gibson, D. Siglec 1 and 2 as potential biomarkers in autoimmune disease. Proteom. Clin. Appl. 2016, 10, 635–644. [Google Scholar] [CrossRef]
- Jiang, H.R.; Hwenda, L.; Makinen, K.; Oetke, C.; Crocker, P.R.; Forrester, J.V. Sialoadhesin promotes the inflammatory response in experimental autoimmune uveoretinitis. J. Immunol. 2006, 177, 2258–2264. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Rauch, U.; Korpos, E.; Song, J.; Loser, K.; Crocker, P.R.; Sorokin, L.M. Sialoadhesin-positive macrophages bind regulatory T cells, negatively controlling their expansion and autoimmune disease progression. J. Immunol. 2009, 182, 6508–6516. [Google Scholar] [CrossRef]
- van den Berg, T.K.; Nath, D.; Ziltener, H.J.; Vestweber, D.; Fukuda, M.; van Die, I.; Crocker, P.R. Cutting edge: CD43 functions as a T cell counterreceptor for the macrophage adhesion receptor sialoadhesin (Siglec-1). J. Immunol. 2001, 166, 3637–3640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashi, N.; Wu, K.; Gronhoj Larsen, C.; Deleuran, M.; Kawana, S.; Yamamoto, K.; Thestrup-Pedersen, K. Expression and function of CD43 and CDw60 on T cells from patients with atopic dermatitis. Acta Derm. Venereol. 2001, 81, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passlick, B.; Flieger, D.; Ziegler-Heitbrock, H.W. Identification and characterization of a novel monocyte subpopulation in human peripheral blood. Blood 1989, 74, 2527–2534. [Google Scholar] [CrossRef] [Green Version]
- Auffray, C.; Sieweke, M.H.; Geissmann, F. Blood monocytes: Development, heterogeneity, and relationship with dendritic cells. Annu. Rev. Immunol. 2009, 27, 669–692. [Google Scholar] [CrossRef] [Green Version]
- Kawanaka, N.; Yamamura, M.; Aita, T.; Morita, Y.; Okamoto, A.; Kawashima, M.; Ueno, A.; Ohmoto, Y.; Makino, H. CD14+,CD16+ blood monocytes and joint inflammation in rheumatoid arthritis. Arthritis Rheum. 2002, 46, 2578–2586. [Google Scholar] [CrossRef]
- Cairns, A.P.; Crockard, A.D.; Bell, A.L. The CD14+ CD16+ monocyte subset in rheumatoid arthritis and systemic lupus erythematosus. Rheumatol. Int. 2002, 21, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Lacerte, P.; Brunet, A.; Egarnes, B.; Duchene, B.; Brown, J.P.; Gosselin, J. Overexpression of TLR2 and TLR9 on monocyte subsets of active rheumatoid arthritis patients contributes to enhance responsiveness to TLR agonists. Arthritis Res. Ther. 2016, 18, 10–11. [Google Scholar] [CrossRef] [Green Version]
- Chara, L.; Sanchez-Atrio, A.; Perez, A.; Cuende, E.; Albarran, F.; Turrion, A.; Chevarria, J.; del Barco, A.A.; Sánchez, M.A.; Monserrat, J.; et al. The number of circulating monocytes as biomarkers of the clinical response to methotrexate in untreated patients with rheumatoid arthritis. J. Transl. Med. 2015, 13, 2. [Google Scholar] [CrossRef] [Green Version]
- Chara, L.; Sanchez-Atrio, A.; Perez, A.; Cuende, E.; Albarran, F.; Turrion, A.; Chevarria, J.; Sánchez, M.A.; Monserrat, J.; de la Hera, A.; et al. Monocyte populations as markers of response to adalimumab plus MTX in rheumatoid arthritis. Arthritis Res. Ther. 2012, 14, R175. [Google Scholar] [CrossRef] [Green Version]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef]
- NICE guideline NG100. Rheumatoid Arthritis in Adults: Diagnosis and Management; National Institute for Health and Care Excellence: London, UK, 2018. [Google Scholar]
- Kikuchi, J.; Hashizume, M.; Kaneko, Y.; Yoshimoto, K.; Nishina, N.; Takeuchi, T. Peripheral blood CD4+ CD25+CD127low regulatory T cells are significantly increased by tocilizumab treatment in patients with rheumatoid arthritis: Increase in regulatory T cells correlates with clinical response. Arthritis Res. Ther. 2015, 17, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata, M.; Ohoka, Y.; Kuwata, T.; Asada, A. Regulation of T cell apoptosis via T cell receptors and steroid receptors. Stem Cells 1996, 14, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, F.; Saegusa, J.; Miyamoto, Y.; Misaki, K.; Kumagai, S.; Morinobu, A. CD45RA-Foxp3high activated/effector regulatory T cells in the CCR7 + CD45RA-CD27 + CD28+central memory subset are decreased in peripheral blood from patients with rheumatoid arthritis. Biochem. Biophys. Res. Commun. 2013, 438, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Walter, G.J.; Fleskens, V.; Frederiksen, K.S.; Rajasekhar, M.; Menon, B.; Gerwien, J.G.; Evans, H.G.; Taams, L.S. Phenotypic, Functional, and Gene Expression Profiling of Peripheral CD45RA+ and CD45RO+ CD4+CD25+CD127low Treg Cells in Patients With Chronic Rheumatoid Arthritis. Arthritis Rheumatol. 2016, 68, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Rossol, M.; Kraus, S.; Pierer, M.; Baerwald, C.; Wagner, U. The CD14bright CD16+ monocyte subset is expanded in rheumatoid arthritis and promotes expansion of the Th17 cell population. Arthritis Rheum. 2012, 64, 671–677. [Google Scholar] [CrossRef]
- Ancuta, P.; Rao, R.; Moses, A.; Mehle, A.; Shaw, S.K.; Luscinskas, F.W.; Gabuzda, D. Fractalkine preferentially mediates arrest and migration of CD16+ monocytes. J. Exp. Med. 2003, 197, 1701–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Greenberg, J.D.; Bhardwaj, N. Dendritic cells as targets for therapy in rheumatoid arthritis. Nat. Rev. Rheumatol. 2009, 5, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Munoz, M.E.; Salas-Vidal, E.; Salaiza-Suazo, N.; Becker, I.; Pedraza-Alva, G.; Rosenstein, Y. The CD43 coreceptor molecule recruits the zeta-chain as part of its signaling pathway. J. Immunol. 2003, 171, 1901–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperling, A.I.; Green, J.M.; Mosley, R.L.; Smith, P.L.; DiPaolo, R.J.; Klein, J.R.; Bluestone, J.A.; Thompson, C.B. CD43 is a murine T cell costimulatory receptor that functions independently of CD28. J. Exp. Med. 1995, 182, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Naito-Matsui, Y.; Takada, S.; Kano, Y.; Iyoda, T.; Sugai, M.; Shimizu, A.; Inaba, K.; Nitschke, L.; Tsubata, T.; Oka, S.; et al. Functional evaluation of activation-dependent alterations in the sialoglycan composition of T cells. J. Biol. Chem. 2014, 289, 1564–1579. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC (n = 21) | DN (n = 6) | DR (n = 24) | DNR (n = 32) | |
|---|---|---|---|---|
| Female, n (%) | 16 (76.2) | 5 (83.3) | 22 (91.7) | 27 (81.8) |
| Age, mean (SD), years | 35.9 (9.2) | 42.5 (16.7) | 59.7 (12.2) | 57.7 (10.1) |
| Disease duration, mean (SD), years | N/A | N/A | 5.0 (3.8) | 10.8 (12.1) |
| Lymphocytes, mean (SD), 109/L | 1.9 (0.4), n = 19 | 1.6 (0.5), n = 6 | 1.7 (0.3), n = 24 | 1.6 (0.5), n = 26 |
| Monocytes, mean (SD), 109/L | 0.5 (0.3), n = 19 | 0.6 (0.2), n = 6 | 0.5 (0.1), n = 24 | 0.6 (0.3), n = 26 |
| ESR, mean (SD), mm/h | N/A | 23.5, (15.2), n = 6 | 12.2 (7.9), n = 24 | 24.1 (20.0), n = 29 |
| CRP, mean (SD), mg/L | N/A | 6.1 (2.9), n = 6 | 5.3 (7.3), n = 22 | 10.6 (9.4), n = 28 |
| DAS28-ESR, mean (SD) | N/A | 4.3 (0.4), n = 4 | 2.6 (1.0), n = 24 | 4.9 (1.1), n = 19 |
| DR | DNR | p Value | |
|---|---|---|---|
| Disease Duration, years | 4.50 ± 3.85 | 9.00 ± 11.43 | <0.01 (**) |
| DAS28-ESR | 2.34 ± 1.05 | 5.51 ± 1.39 | <0.01 (**) |
| ESR, mm/h | 11.00 ± 7.93 | 21.50 ± 16.10 | 0.01 (*) |
| CRP, mg/L | 2.75 ± 7.30 | 8.80 ± 24.80 | 0.08 (ns) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eakin, A.J.; Ahmed, T.; McGeough, C.M.; Drain, S.; Alexander, H.D.; Wright, G.D.; Gardiner, P.V.; Small, D.; Bjourson, A.J.; Gibson, D.S. CD169+ Monocyte and Regulatory T Cell Subsets Are Associated with Disease Activity in Rheumatoid Arthritis. J. Pers. Med. 2022, 12, 1875. https://doi.org/10.3390/jpm12111875
Eakin AJ, Ahmed T, McGeough CM, Drain S, Alexander HD, Wright GD, Gardiner PV, Small D, Bjourson AJ, Gibson DS. CD169+ Monocyte and Regulatory T Cell Subsets Are Associated with Disease Activity in Rheumatoid Arthritis. Journal of Personalized Medicine. 2022; 12(11):1875. https://doi.org/10.3390/jpm12111875
Chicago/Turabian StyleEakin, Amanda J., Tahanver Ahmed, Cathy M. McGeough, Stephen Drain, H. Denis Alexander, Gary D. Wright, Philip V. Gardiner, Dawn Small, Anthony J. Bjourson, and David S. Gibson. 2022. "CD169+ Monocyte and Regulatory T Cell Subsets Are Associated with Disease Activity in Rheumatoid Arthritis" Journal of Personalized Medicine 12, no. 11: 1875. https://doi.org/10.3390/jpm12111875