
The Downregulation of LSAMP Expression Promotes Lung Cancer Progression and Is Associated with Poor Survival Prognosis

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Next-Generation Sequencing
2.3. Bioinformatics
2.4. DNA Methylation and Copy Number
2.5. miRNAs and LSAMP Correlation
2.6. Gene and Protein Interaction of LSAMP
2.7. LSAMP Knockdown
2.8. Cell Proliferation, Transwell Migration, Wound Healing, and Tumor Spheroid Formation
2.9. Immunoblot
2.10. Statistical Analysis
3. Results
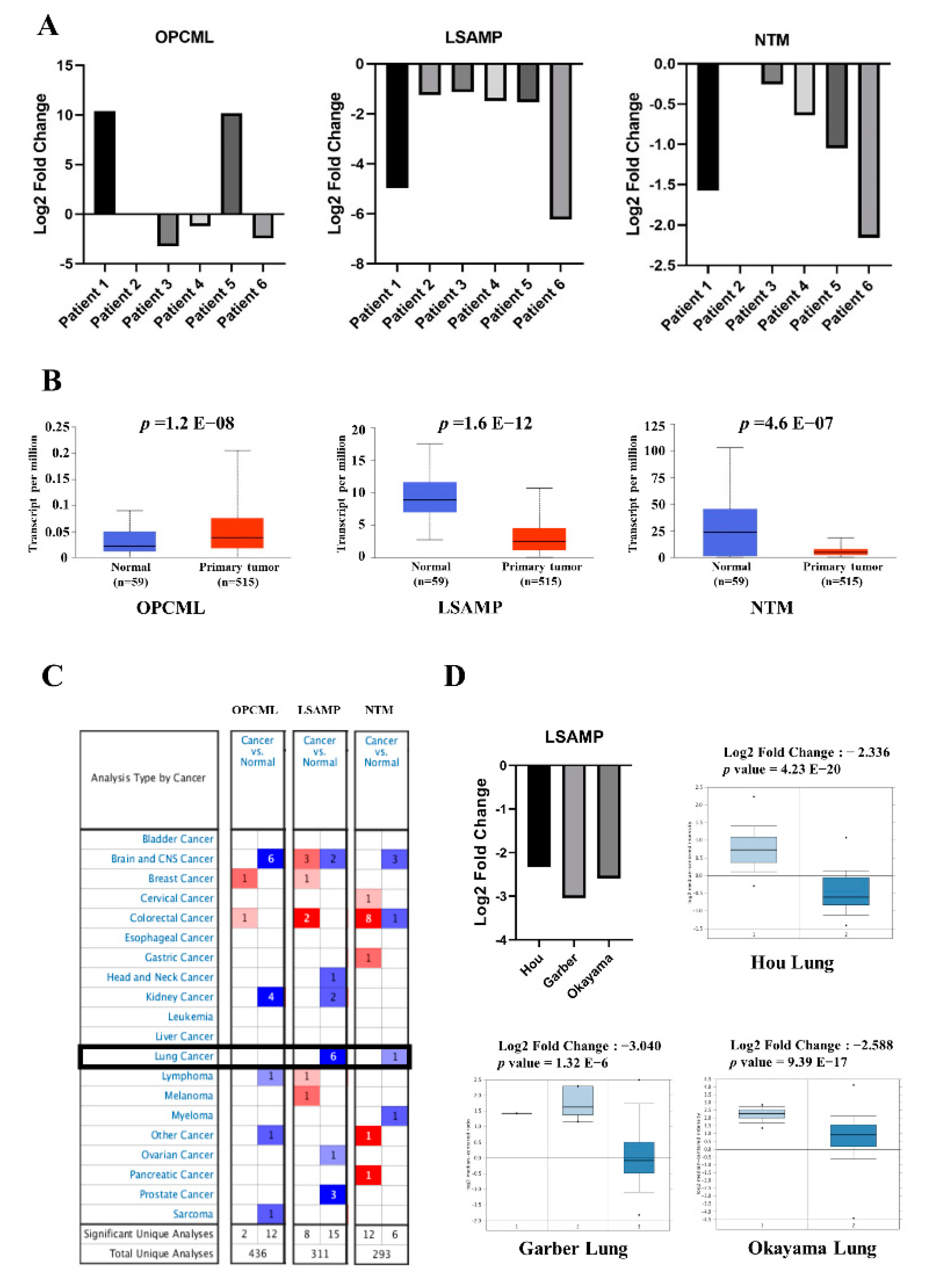
3.1. The Downregulated Expressions of LSAMP and NTM Genes in Lung Adenocarcinoma
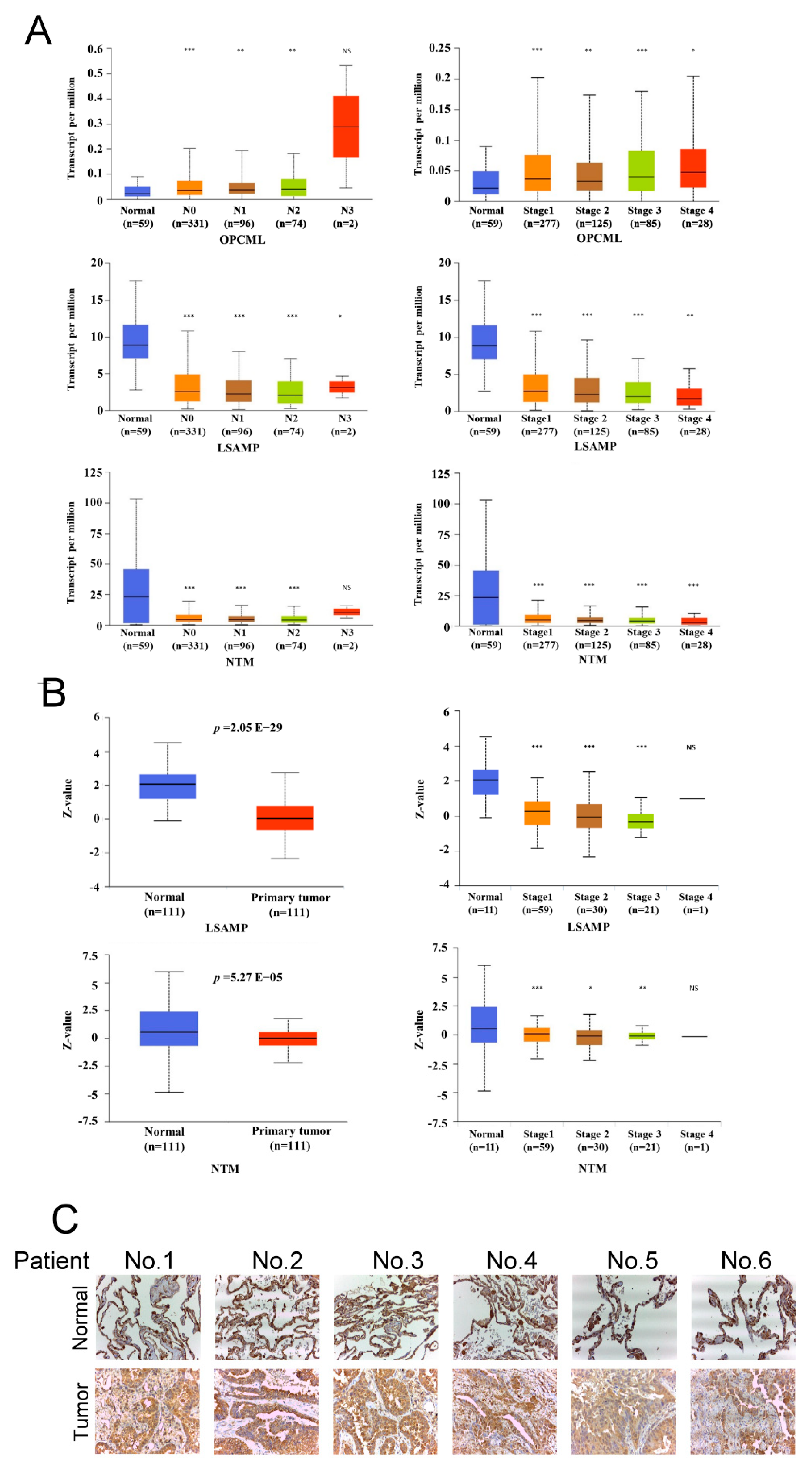
3.2. LSAMP Expression Is Negatively Correlated with Tumor Aggressiveness
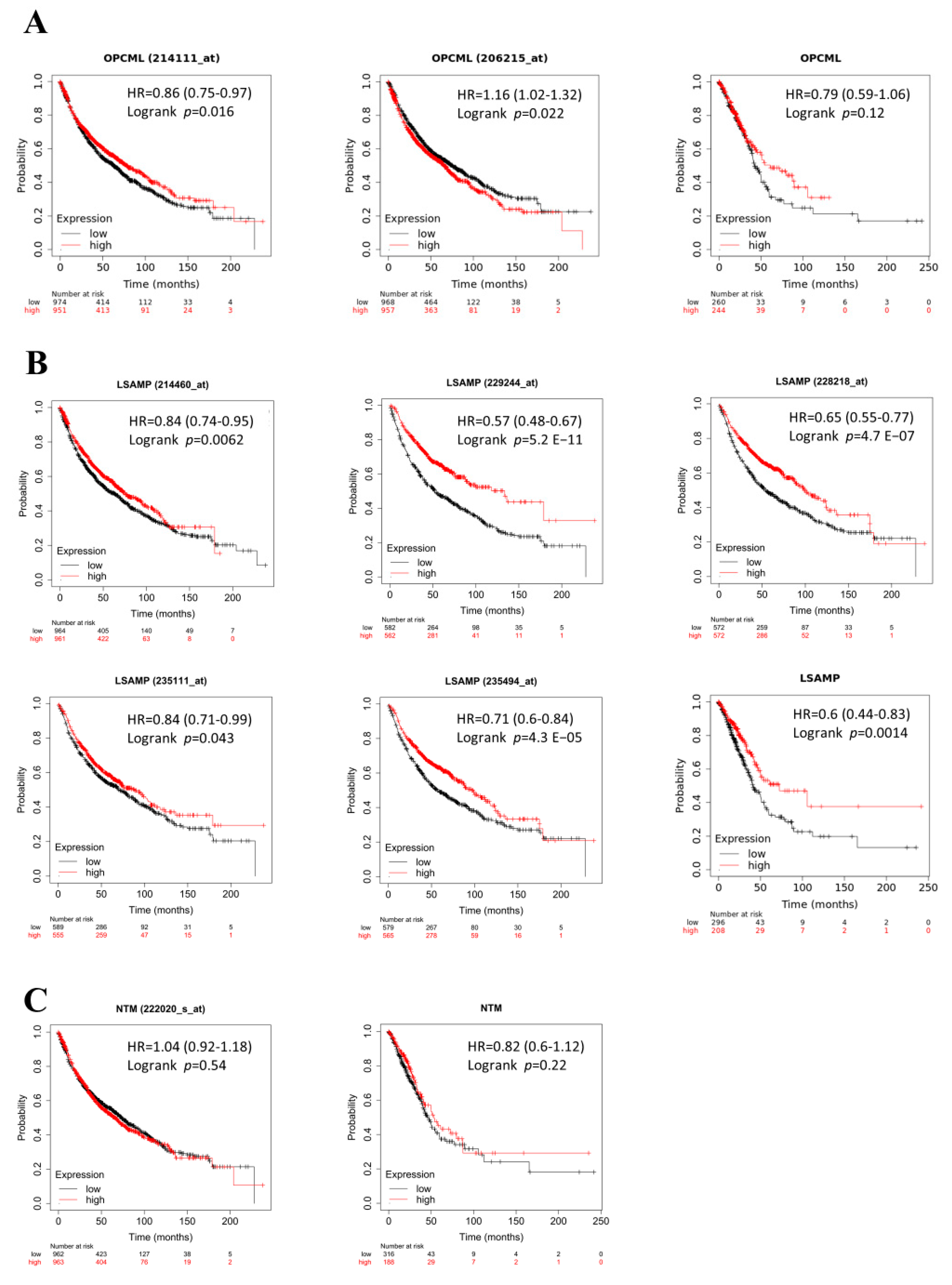
3.3. The Lower Levels of LSAMP Conferred Poor Survival Prognosis Clinically
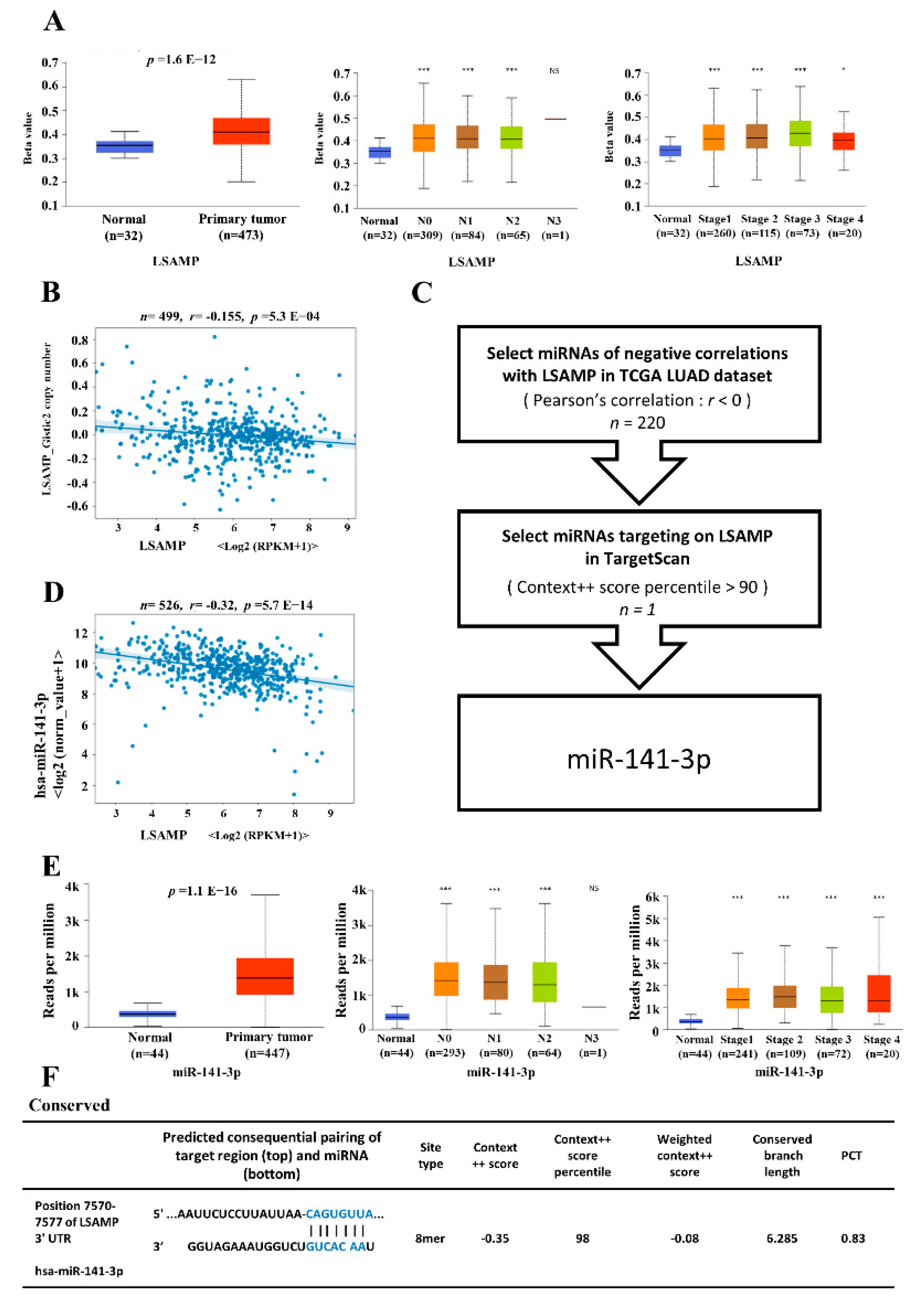
3.4. The Epigenetic Regulatory Mechanisms for LSAMP Expression
3.5. The NEGR1 Interacts with LSAMP Expression
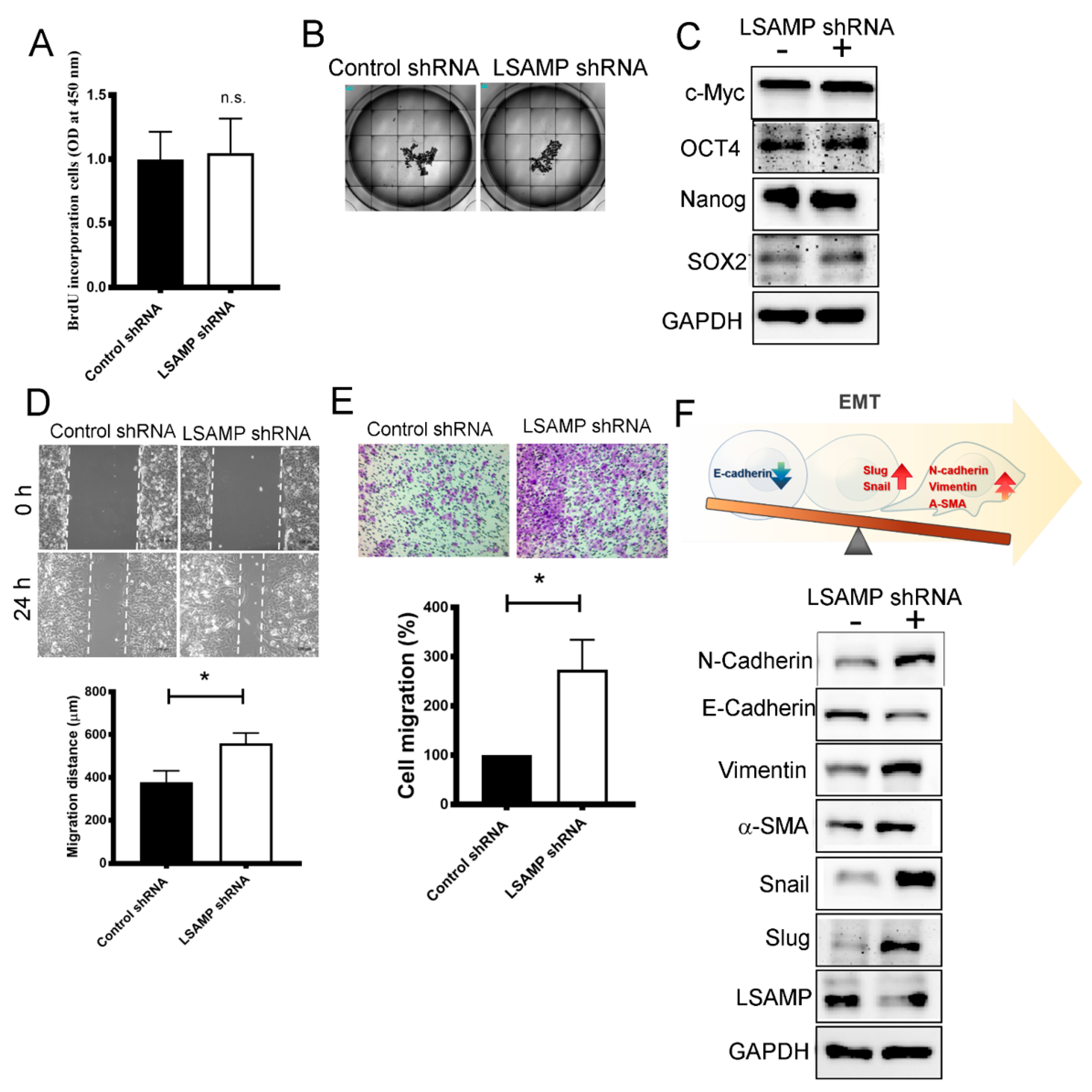
3.6. LSAMP Interferes with Epithelial-Mesenchymal Transition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Hsu, J.C.; Wei, C.-F.; Yang, S.-C.; Lin, P.-C.; Lee, Y.-C.; Lu, C.Y. Lung cancer survival and mortality in Taiwan following the initial launch of targeted therapies: An interrupted time series study. BMJ Open 2020, 10, e033427. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2020, 71, 209–249. [Google Scholar] [CrossRef]
- Gristina, V.; Malapelle, U.; Galvano, A.; Pisapia, P.; Pepe, F.; Rolfo, C.; Tortorici, S.; Bazan, V.; Troncone, G.; Russo, A. The significance of epidermal growth factor receptor uncommon mutations in non-small cell lung cancer: A systematic review and critical appraisal. Cancer Treat. Rev. 2020, 85, 101994. [Google Scholar] [CrossRef] [PubMed]
- Corrales, L.; Scilla, K.; Caglevic, C.; Miller, K.; Oliveira, J.; Rolfo, C. Immunotherapy in Lung Cancer: A New Age in Cancer Treatment. Adv. Exp. Med. Biol. 2018, 995, 65–95. [Google Scholar] [CrossRef]
- Mittal, V. Epithelial Mesenchymal Transition in Aggressive Lung Cancers. Adv. Exp. Med. Biol. 2016, 890, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Wang, S.; Wang, S.; Wu, H.; Yu, J.; Chen, Q.; Cui, W.; Yuan, Y.; Wen, X.; He, J.; et al. SOX5 interacts with YAP1 to drive malignant potential of non-small cell lung cancer cells. Am. J. Cancer Res. 2018, 8, 866–878. [Google Scholar] [PubMed]
- Banyard, J.; Bielenberg, D.R. The role of EMT and MET in cancer dissemination. Connect. Tissue Res. 2015, 56, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Levitt, P. A monoclonal antibody to limbic system neurons. Science 1984, 223, 299–301. [Google Scholar] [CrossRef]
- Struyk, A.; Canoll, P.; Wolfgang, M.; Rosen, C.; D’Eustachio, P.; Salzer, J. Cloning of neurotrimin defines a new subfamily of differentially expressed neural cell adhesion molecules. J. Neurosci. 1995, 15, 2141–2156. [Google Scholar] [CrossRef] [Green Version]
- Funatsu, N.; Miyata, S.; Kumanogoh, H.; Shigeta, M.; Hamada, K.; Endo, Y.; Sokawa, Y.; Maekawa, S. Characterization of a novel rat brain glycosylphosphatidylinositol-anchored protein (Kilon), a member of the IgLON cell adhesion molecule family. J. Biol. Chem. 1999, 274, 8224–8230. [Google Scholar] [CrossRef] [Green Version]
- Nissen, M.S.; Blaabjerg, M. Anti-IgLON5 Disease: A Case With 11-Year Clinical Course and Review of the Literature. Front. Neurol. 2019, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Akeel, M.; McNamee, C.J.; Youssef, S.; Moss, D. DIgLONs inhibit initiation of neurite outgrowth from forebrain neurons via an IgLON-containing receptor complex. Brain Res. 2011, 1374, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Pischedda, F.; Piccoli, G. The IgLON Family Member Negr1 Promotes Neuronal Arborization Acting as Soluble Factor via FGFR2. Front. Mol. Neurosci. 2015, 8, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Maekawa, S.; Miyata, S. IgLON cell adhesion molecules regulate synaptogenesis in hippocampal neurons. Cell Biochem. Funct. 2009, 27, 496–498. [Google Scholar] [CrossRef]
- Karis, K.; Eskla, K.-L.; Kaare, M.; Täht, K.; Tuusov, J.; Visnapuu, T.; Innos, J.; Jayaram, M.; Timmusk, T.; Weickert, C.S.; et al. Altered Expression Profile of IgLON Family of Neural Cell Adhesion Molecules in the Dorsolateral Prefrontal Cortex of Schizophrenic Patients. Front. Mol. Neurosci. 2018, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Jayaram, M.; Kaare, M.; Leidmaa, E.; Jagomäe, T.; Heinla, I.; Hickey, M.A.; Kaasik, A.; Schäfer, M.K.; Innos, J.; et al. Neural cell adhesion molecule Negr1 deficiency in mouse results in structural brain endophenotypes and behavioral deviations related to psychiatric disorders. Sci. Rep. 2019, 9, 5457. [Google Scholar] [CrossRef]
- Vanaveski, T.; Singh, K.; Narvik, J.; Eskla, K.-L.; Visnapuu, T.; Heinla, I.; Jayaram, M.; Innos, J.; Lilleväli, K.; Philips, M.-A.; et al. Promoter-Specific Expression and Genomic Structure of IgLON Family Genes in Mouse. Front. Neurosci. 2017, 11, 38. [Google Scholar] [CrossRef] [Green Version]
- Ntougkos, E.; Rush, R.; Scott, D.; Frankenberg, T.; Gabra, H.; Smyth, J.F.; Sellar, G.C. The IgLON Family in Epithelial Ovarian Cancer: Expression Profiles and Clinicopathologic Correlates. Clin. Cancer Res. 2005, 11, 5764–5768. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Xu, J.; Wang, Y.; Heng, X.; Yang, L.; Xing, X. Loss of opioid binding protein/cell adhesion molecule-like gene expression in gastric cancer. Oncol. Lett. 2018, 15, 9973–9977. [Google Scholar] [CrossRef] [Green Version]
- Barøy, T.; Kresse, S.H.; Skårn, M.; Stabell, M.; Castro, R.; Lauvrak, S.; Llombart-Bosch, A.; Myklebost, O.; Meza-Zepeda, L.A. Reexpression of LSAMP inhibits tumor growth in a preclinical osteosarcoma model. Mol. Cancer 2014, 13, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pros, E.; Saigi, M.; Alameda, D.; Gomez-Mariano, G.; Martinez-Delgado, B.; Alburquerque-Bejar, J.J.; Carretero, J.; Tonda, R.; Esteve-Codina, A.; Catala, I.; et al. Genome-wide profiling of non-smoking-related lung cancer cells reveals common RB1 rearrangements associated with histopathologic transformation in EGFR-mutant tumors. Ann. Oncol. 2020, 31, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Ocak, S.; Sos, M.L.; Thomas, R.K.; Massion, P.P. High-throughput molecular analysis in lung cancer: Insights into biology and potential clinical applications. Eur. Respir. J. 2009, 34, 489–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakai, T.; Prasoon, P.; Hirose, Y.; Shimada, Y.; Ichikawa, H.; Nagahashi, M. Next-generation sequencing-based clinical sequencing: Toward precision medicine in solid tumors. Int. J. Clin. Oncol. 2019, 24, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Nagahashi, M.; Shimada, Y.; Ichikawa, H.; Kameyama, H.; Takabe, K.; Okuda, S.; Wakai, T. Next generation sequencing-based gene panel tests for the management of solid tumors. Cancer Sci. 2019, 110, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, Y.L.; Hung, J.Y.; Lee, Y.L.; Chen, F.W.; Chang, K.F.; Chang, W.A.; Tsai, Y.M.; Chong, I.W.; Kuo, P.L. Identification of novel gene expression signature in lung adenocarcinoma by using next-generation sequencing data and bioinformatics analysis. Oncotarget 2017, 8, 104831–104854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Devreotes, P.; Horwitz, A.R. Signaling Networks that Regulate Cell Migration. Cold Spring Harbor Perspect. Biol. 2015, 7, a005959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Q.S. Targeting non-small cell lung cancer: Driver mutation beyond epidermal growth factor mutation and anaplastic lymphoma kinase fusion. Ther. Adv. Med. Oncol. 2020, 12, 1758835919895756. [Google Scholar] [CrossRef] [Green Version]
- Kresse, S.H.; Ohnstad, H.O.; Paulsen, E.B.; Bjerkehagen, B.; Szuhai, K.; Serra, M.; Schaefer, K.L.; Myklebost, O.; Meza-Zepeda, L.A. LSAMP, a novel candidate tumor suppressor gene in human osteosarcomas, identified by array comparative genomic hybridization. Genes Chromosomes Cancer 2009, 48, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Lilleväli, K.; Gilbert, S.F.; Bregin, A.; Narvik, J.; Jayaram, M.; Rahi, M.; Innos, J.; Kaasik, A.; Vasar, E.; et al. The combined impact of IgLON family proteins Lsamp and Neurotrimin on developing neurons and behavioral profiles in mouse. Brain Res. Bull. 2018, 140, 5–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harjunpää, H.; Llort Asens, M.; Guenther, C.; Fagerholm, S.C. Cell Adhesion Molecules and Their Roles and Regulation in the Immune and Tumor Microenvironment. Front. Immunol. 2019, 10, 1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Gender | Age | Stage | AJCC, 8th Ed. | Comorbidity |
|---|---|---|---|---|---|
| 01 | M | 70 | 2B | T3N0M0 | Old stroke |
| 02 | M | 66 | 4B | T2aN0M1c | Hypertension |
| 03 | F | 51 | 1B | T2aN0M0 | Hepatitis B carrier |
| 04 | M | 53 | 3A | T3N2M0 | Hypertension |
| 05 | F | 60 | 1A | T1bN0M0 | Hepatitis B carrier |
| 06 | M | 67 | 1A | T1aN0M0 | UTUC, 2016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.-Y.; Wu, K.-L.; Chang, Y.-Y.; Liu, Y.-W.; Huang, Y.-C.; Jian, S.-F.; Lin, Y.-S.; Tsai, P.-H.; Hung, J.-Y.; Tsai, Y.-M.; et al. The Downregulation of LSAMP Expression Promotes Lung Cancer Progression and Is Associated with Poor Survival Prognosis. J. Pers. Med. 2021, 11, 578. https://doi.org/10.3390/jpm11060578
Chang C-Y, Wu K-L, Chang Y-Y, Liu Y-W, Huang Y-C, Jian S-F, Lin Y-S, Tsai P-H, Hung J-Y, Tsai Y-M, et al. The Downregulation of LSAMP Expression Promotes Lung Cancer Progression and Is Associated with Poor Survival Prognosis. Journal of Personalized Medicine. 2021; 11(6):578. https://doi.org/10.3390/jpm11060578
Chicago/Turabian StyleChang, Chao-Yuan, Kuan-Li Wu, Yung-Yun Chang, Yu-Wei Liu, Yung-Chi Huang, Shu-Fang Jian, Yi-Shiuan Lin, Pei-Hsun Tsai, Jen-Yu Hung, Ying-Ming Tsai, and et al. 2021. "The Downregulation of LSAMP Expression Promotes Lung Cancer Progression and Is Associated with Poor Survival Prognosis" Journal of Personalized Medicine 11, no. 6: 578. https://doi.org/10.3390/jpm11060578