
Estimation of Lewis Blood Group Status by Fluorescence Melting Curve Analysis in Simultaneous Genotyping of c.385A>T and Fusion Gene in FUT2 and c.59T>G and c.314C>T in FUT3

Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Samples
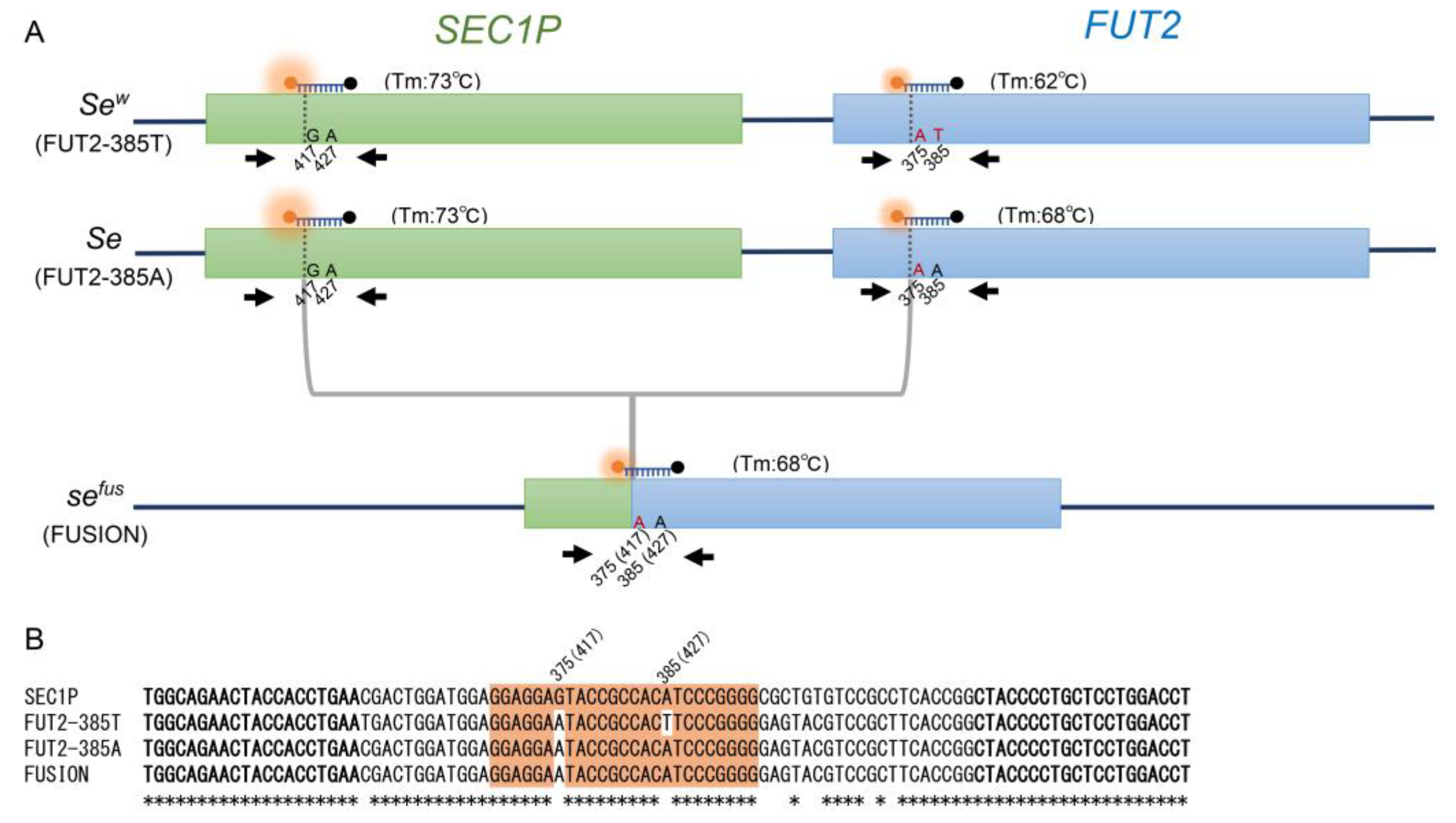
2.2. Asymmetric PCR for c.385A>T and sefus of FUT2
2.3. Triplex PCR for c.385A>T and sefus of FUT2, c.59T>G and c.314C>T of FUT3
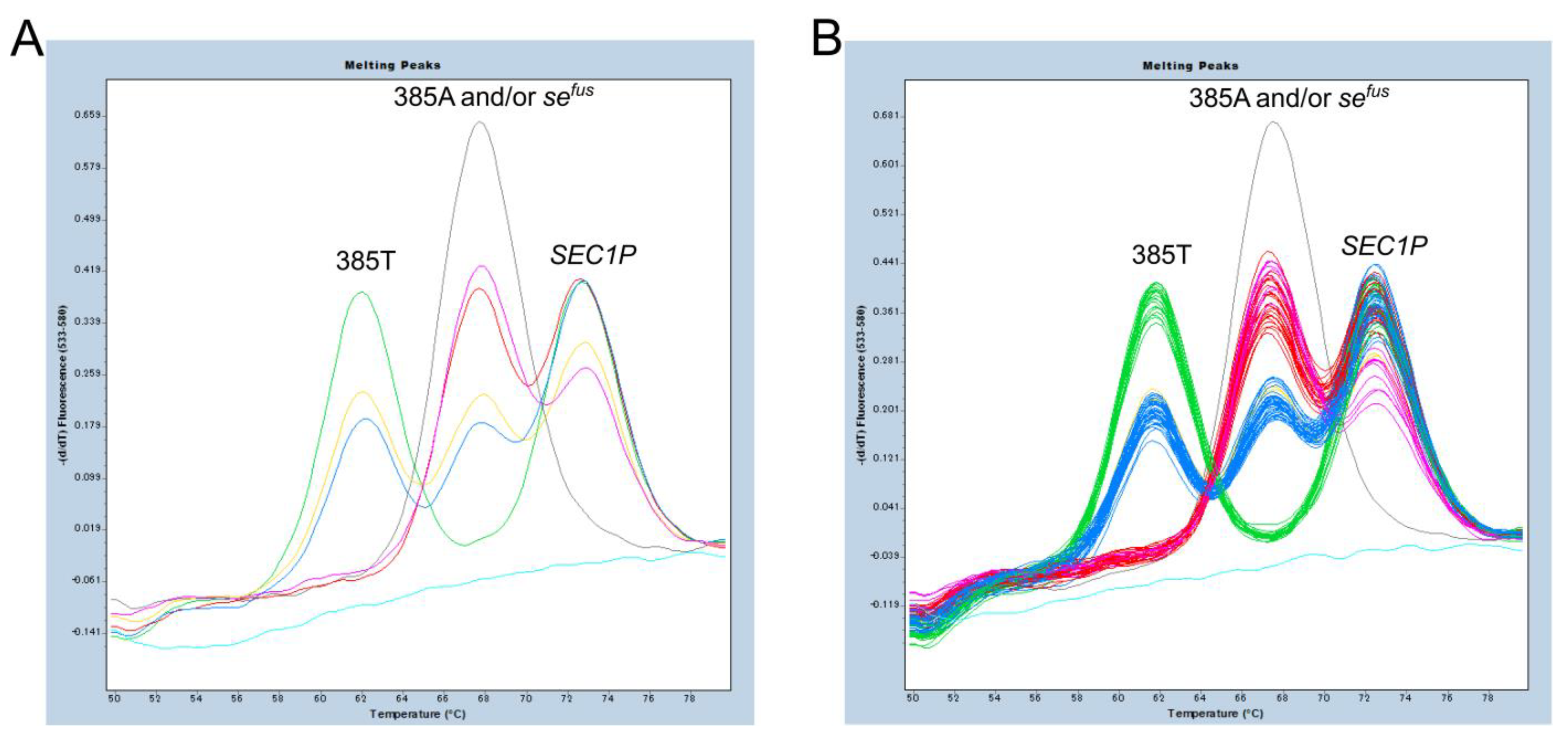
2.4. FMCA for Detection of c.385A>T and sefus of FUT2, c.59T>G and c.314C>T of FUT3
3. Results
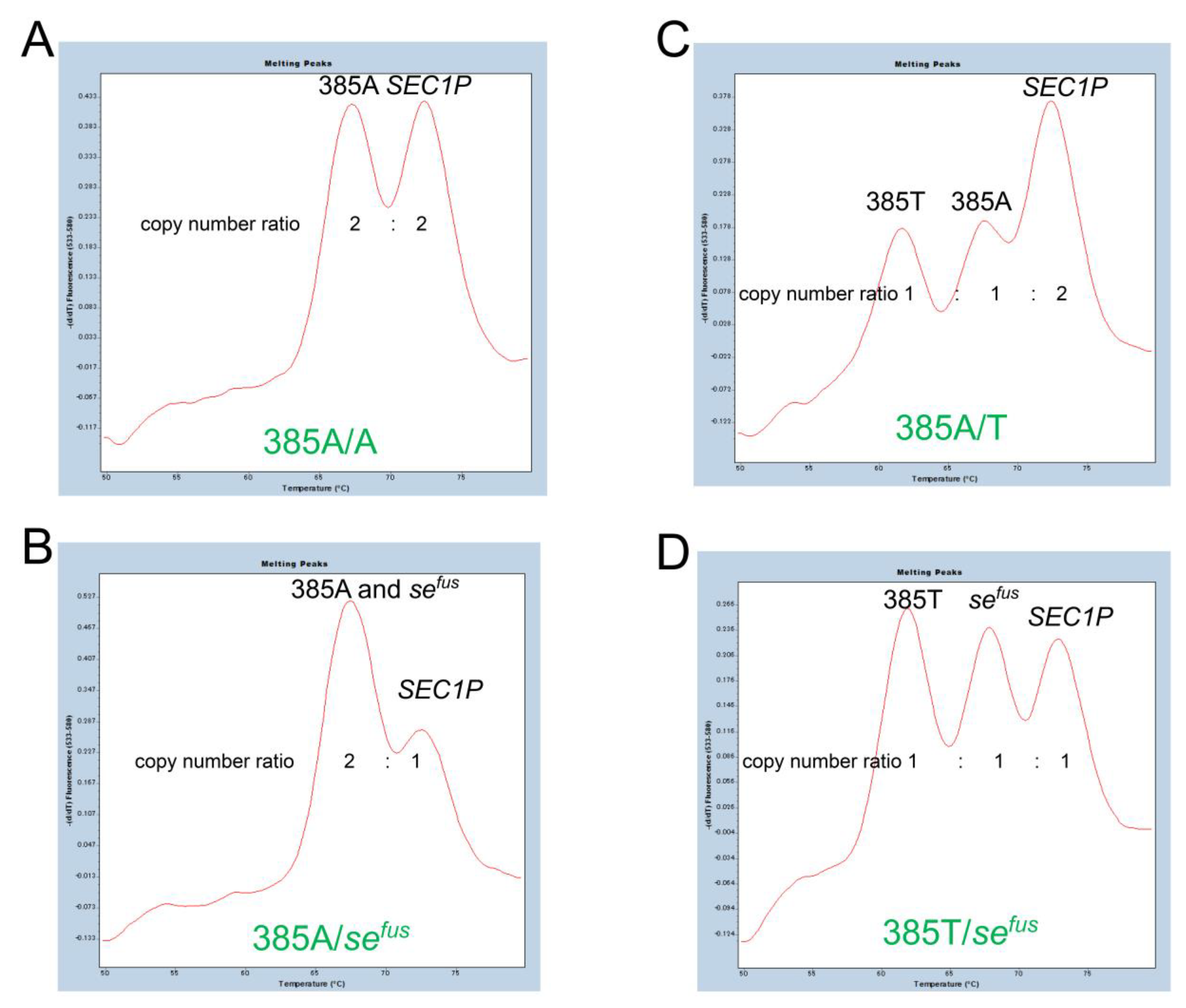
3.1. FMCA for Detection of c.385A>T and sefus of FUT2
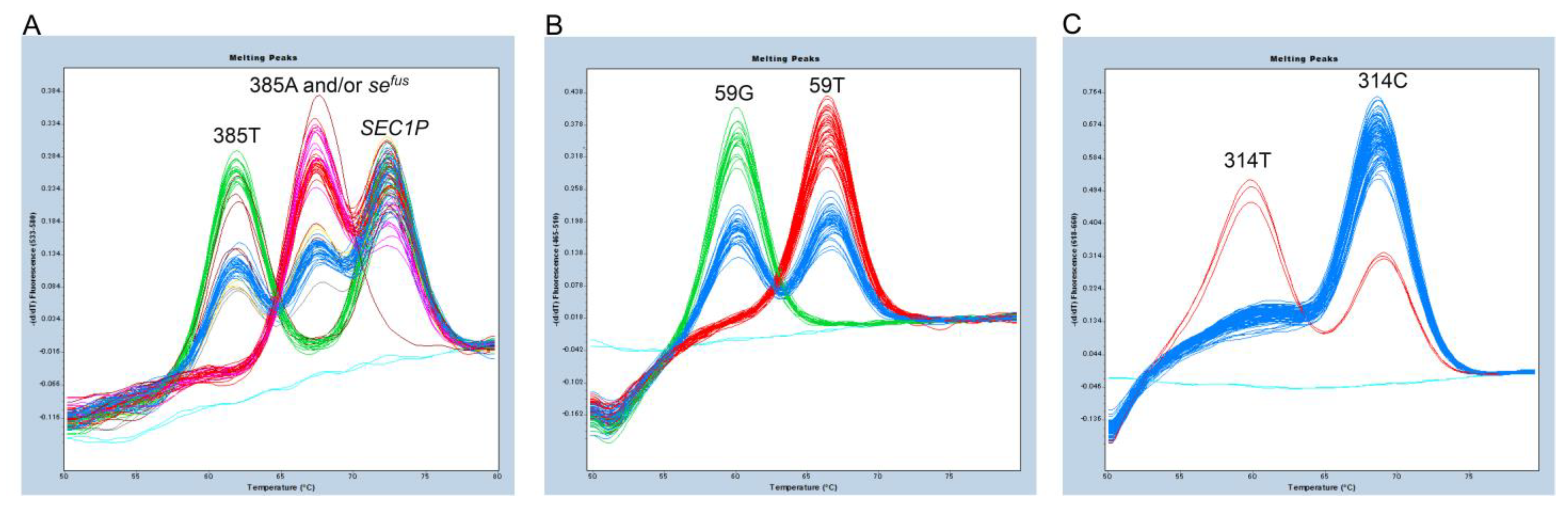
3.2. Triplex FMCA for Detection of c.385A>T and sefus of FUT2, c.59T>G and c.314C>T of FUT3
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oriol, R.; Danilovs, J.; Hawkins, B.R. A new genetic model proposing that the Se gene is a structural gene closely linked to the H gene. Am. J. Hum. Genet. 1981, 33, 421–431. [Google Scholar]
- Clausen, H.; Hakomori, S. ABH and related histo-blood group antigens; immunochemical differences in carrier isotypes and their distribution. Vox Sang. 1989, 56, 1–20. [Google Scholar] [CrossRef]
- Daniels, G. ABO, H, and Lewis Systems. In Human Blood Groups, 3rd ed.; Daniels, G., Ed.; Wiley-Blackwell: West Sussex, UK, 2013; pp. 11–95. [Google Scholar] [CrossRef]
- Henry, S.; Mollicone, R.; Fernandez, P.; Samuelsson, B.; Oriol, R.; Larson, G. Molecular basis for erythrocyte Le(a+ b+) and salivary ABH partial-secretor phenotypes: Expression of a FUT2 secretor allele with an A-->T mutation at nucleotide 385 correlates with reduced alpha(1,2) fucosyltransferase activity. Glycoconj. J. 1996, 13, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Koda, Y.; Soejima, M.; Liu, Y.; Kimura, H. Molecular basis for secretor type alpha(1,2)-fucosyltransferase gene deficiency in a Japanese population: A fusion gene generated by unequal crossover responsible for the enzyme deficiency. Am. J. Hum. Genet. 1996, 59, 343–350. [Google Scholar] [PubMed]
- Boren, T.; Falk, P.; Roth, K.A.; Larson, G.; Normark, S. Attachment of Helicobacter pylori to human gastric epithelium mediated by blood group antigens. Science 1993, 262, 1892–1895. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Niang, M.A.; N’Doye, I.; Critchlow, C.W.; Hawes, S.E.; Hill, A.V.; Kiviat, N.B. Secretor polymorphism and human immunodeficiency virus infection in Senegalese women. J. Infect. Dis. 2000, 181, 737–739. [Google Scholar] [CrossRef]
- Cakir, B.; Heiss, G.; Pankow, J.S.; Salomaa, V.; Sharrett, A.R.; Couper, D.; Weston, B.W. Association of the Lewis genotype with cardiovascular risk factors and subclinical carotid atherosclerosis: The Atherosclerosis Risk in Communities (ARIC) study. J. Intern. Med. 2004, 255, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Hazra, A.; Kraft, P.; Selhub, J.; Giovannucci, E.L.; Thomas, G.; Hoover, R.N.; Chanock, S.J.; Hunter, D.J. Common variants of FUT2 are associated with plasma vitamin B12 levels. Nat. Genet. 2008, 40, 1160–1162. [Google Scholar] [CrossRef] [Green Version]
- McGovern, D.P.; Jones, M.R.; Taylor, K.D.; Marciante, K.; Yan, X.; Dubinsky, M.; Ippoliti, A.; Vasiliauskas, E.; Berel, D.; Derkowski, C.; et al. Fucosyltransferase 2 (FUT2) non-secretor status is associated with Crohn’s disease. Hum. Mol. Genet. 2010, 19, 3468–3476. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Zhang, D.; Zheng, S.; Guo, M.; Lin, X.; Jiang, Y. Association of Ulcerative Colitis with FUT2 and FUT3 Polymorphisms in Patients from Southeast China. PLoS ONE 2016, 11, e0146557. [Google Scholar] [CrossRef] [Green Version]
- Santos-Cortez, R.L.P.; Chiong, C.M.; Frank, D.N.; Ryan, A.F.; Giese, A.P.J.; Bootpetch Roberts, T.; Daly, K.A.; Steritz, M.J.; Szeremeta, W.; Pedro, M.; et al. FUT2 Variants Confer Susceptibility to Familial Otitis Media. Am. J. Hum. Genet. 2018, 103, 679–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Han, R.; Chen, M.; Liu, R.; Wu, M.; Zhang, X.; Ma, Y.; Yuan, Y.; Wang, R.; Shuai, Z.; et al. Associations between fucosyltransferase 3 gene polymorphisms and ankylosing spondylitis: A case-control study of an east Chinese population. PLoS ONE 2020, 15, e0237219. [Google Scholar] [CrossRef] [PubMed]
- Loureiro Tonini, M.A.; Pires Goncalves Barreira, D.M.; Bueno de Freitas Santolin, L.; Bondi Volpini, L.P.; Gagliardi Leite, J.P.; Le Moullac-Vaidye, B.; Le Pendu, J.; Cruz Spano, L. FUT2, Secretor Status and FUT3 Polymorphisms of Children with Acute Diarrhea Infected with Rotavirus and Norovirus in Brazil. Viruses 2020, 12, 1084. [Google Scholar] [CrossRef] [PubMed]
- Matzhold, E.M.; Berghold, A.; Bemelmans, M.K.B.; Banfi, C.; Stelzl, E.; Kessler, H.H.; Steinmetz, I.; Krause, R.; Wurzer, H.; Schlenke, P.; et al. Lewis and ABO histo-blood types and the secretor status of patients hospitalized with COVID-19 implicate a role for ABO antibodies in susceptibility to infection with SARS-CoV-2. Transfusion 2021, 61, 2736–2745. [Google Scholar] [CrossRef]
- Koda, Y.; Tachida, H.; Pang, H.; Liu, Y.; Soejima, M.; Ghaderi, A.A.; Takenaka, O.; Kimura, H. Contrasting patterns of polymorphisms at the ABO-secretor gene (FUT2) and plasma alpha(1,3)fucosyltransferase gene (FUT6) in human populations. Genetics 2001, 158, 747–756. [Google Scholar] [CrossRef]
- Ferrer-Admetlla, A.; Sikora, M.; Laayouni, H.; Esteve, A.; Roubinet, F.; Blancher, A.; Calafell, F.; Bertranpetit, J.; Casals, F. A natural history of FUT2 polymorphism in humans. Mol. Biol. Evol. 2009, 26, 1993–2003. [Google Scholar] [CrossRef] [Green Version]
- Soejima, M.; Fujimoto, R.; Agusa, T.; Iwata, H.; Fujihara, J.; Takeshita, H.; Minh, T.B.; Trang, P.T.; Viet, P.H.; Nakajima, T.; et al. Genetic variation of FUT2 in a Vietnamese population: Identification of two novel Se enzyme-inactivating mutations. Transfusion 2012, 52, 1268–1275. [Google Scholar] [CrossRef]
- Soejima, M.; Koda, Y. High-resolution melting analysis for detection of fusion allele of FUT2. Electrophoresis 2021, 42, 315–318. [Google Scholar] [CrossRef]
- Rouquier, S.; Lowe, J.B.; Kelly, R.J.; Fertitta, A.L.; Lennon, G.G.; Giorgi, D. Molecular cloning of a human genomic region containing the H blood group alpha(1,2)fucosyltransferase gene and two H locus-related DNA restriction fragments. Isolation of a candidate for the human Secretor blood group locus. J. Biol. Chem. 1995, 270, 4632–4639. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.J.; Rouquier, S.; Giorgi, D.; Lennon, G.G.; Lowe, J.B. Sequence and expression of a candidate for the human Secretor blood group alpha(1,2)fucosyltransferase gene (FUT2). Homozygosity for an enzyme-inactivating nonsense mutation commonly correlates with the non-secretor phenotype. J. Biol. Chem. 1995, 270, 4640–4649. [Google Scholar] [CrossRef] [Green Version]
- Henry, S.; Mollicone, R.; Fernandez, P.; Samuelsson, B.; Oriol, R.; Larson, G. Homozygous expression of a missense mutation at nucleotide 385 in the FUT2 gene associates with the Le(a+b+) partial-secretor phenotype in an Indonesian family. Biochem. Biophys. Res. Commun. 1996, 219, 675–678. [Google Scholar] [CrossRef]
- Moller, M.; Joud, M.; Storry, J.R.; Olsson, M.L. Erythrogene: A database for in-depth analysis of the extensive variation in 36 blood group systems in the 1000 Genomes Project. Blood Adv. 2016, 1, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Soejima, M.; Koda, Y. Fluorescence Melting Curve Analysis for Concurrent Genotyping of Three Tag SNPs in FUT3. Diagnostics 2022, 12, 3039. [Google Scholar] [CrossRef] [PubMed]
- El Housni, H.; Heimann, P.; Parma, J.; Vassart, G. Single-nucleotide polymorphism genotyping by melting analysis of dual-labeled probes: Examples using factor V Leiden and prothrombin 20210A mutations. Clin. Chem. 2003, 49, 1669–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Liu, Z.; Liao, Y.; Chen, X.; Zhang, Y.; Li, Q. Multiplex fluorescence melting curve analysis for mutation detection with dual-labeled, self-quenched probes. PLoS ONE 2011, 6, e19206. [Google Scholar] [CrossRef] [PubMed]
- Soejima, M.; Koda, Y. Detection of the weak-secretor rs1047781 (385A>T) single nucleotide polymorphism using an unlabeled probe high-resolution melting-based method. Electrophoresis 2021, 42, 1362–1365. [Google Scholar] [CrossRef]
- Soejima, M.; Koda, Y. TaqMan real-time polymerase chain reaction for detection of SEC1-FUT2 hybrid alleles: Identification of novel hybrid allele. Clin. Chim. Acta 2013, 415, 59–62. [Google Scholar] [CrossRef]
- Good, A.H.; Yau, O.; Lamontagne, L.R.; Oriol, R. Serological and chemical specificities of twelve monoclonal anti-Lea and anti-Leb antibodies. Vox Sang. 1992, 62, 180–189. [Google Scholar] [CrossRef]
- Hutson, A.M.; Airaud, F.; LePendu, J.; Estes, M.K.; Atmar, R.L. Norwalk virus infection associates with secretor status genotyped from sera. J. Med. Virol. 2005, 77, 116–120. [Google Scholar] [CrossRef]
- Koda, Y.; Ishida, T.; Tachida, H.; Wang, B.; Pang, H.; Soejima, M.; Soemantri, A.; Kimura, H. DNA sequence variation of the human ABO-secretor locus (FUT2) in New Guinean populations: Possible early human migration from Africa. Hum. Genet. 2003, 113, 534–541. [Google Scholar] [CrossRef]
- Soejima, M.; Pang, H.; Koda, Y. Genetic variation of FUT2 in a Ghanaian population: Identification of four novel mutations and inference of balancing selection. Ann. Hematol. 2007, 86, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Hippman, C.; Nislow, C. Pharmacogenomic Testing: Clinical Evidence and Implementation Challenges. J. Pers. Med. 2019, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Risch, N.J. Searching for genetic determinants in the new millennium. Nature 2000, 405, 847–856. [Google Scholar] [CrossRef]
- Amundadottir, L.; Kraft, P.; Stolzenberg-Solomon, R.Z.; Fuchs, C.S.; Petersen, G.M.; Arslan, A.A.; Bueno-de-Mesquita, H.B.; Gross, M.; Helzlsouer, K.; Jacobs, E.J.; et al. Genome-wide association study identifies variants in the ABO locus associated with susceptibility to pancreatic cancer. Nat. Genet. 2009, 41, 986–990. [Google Scholar] [CrossRef] [Green Version]
- Vasan, S.K.; Rostgaard, K.; Majeed, A.; Ullum, H.; Titlestad, K.E.; Pedersen, O.B.; Erikstrup, C.; Nielsen, K.R.; Melbye, M.; Nyren, O.; et al. ABO Blood Group and Risk of Thromboembolic and Arterial Disease: A Study of 1.5 Million Blood Donors. Circulation 2016, 133, 1449–1457, discussion 1457. [Google Scholar] [CrossRef] [PubMed]
- Moslemi, C.; Saekmose, S.; Larsen, R.; Brodersen, T.; Didriksen, M.; Hjalgrim, H.; Banasik, K.; Nielsen, K.R.; Bruun, M.T.; Dowsett, J.; et al. A large cohort study of the effects of Lewis, ABO, 13 other blood groups, and secretor status on COVID-19 susceptibility, severity, and long COVID-19. Transfusion 2023, 63, 47–58. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| c.385A>T | Fusion Gene | FUT2 Genotype | Secretor Phenotype | c.59T>G | c.314C>T | FUT3 Genotype | Lewis Phenotype | Number |
|---|---|---|---|---|---|---|---|---|
| A/A | - | Se/Se | Secretor | T/T | C/C | Le/Le | Le(a-b+) | 11 |
| C/T | Le/le202,314 | Le(a-b+) | 2 | |||||
| T/G | C/C | Le/le59 | Le(a-b+) | 7 | ||||
| C/T | le59/le202,314 | Le(a-b-) | 1 | |||||
| G/G | C/C | le59/le59 | Le(a-b-) | 3 | ||||
| A/T | - | Se/Sew | Secretor | T/T | C/C | Le/Le | Le(a-b+) | 13 |
| T/G | C/C | Le/le59 | Le(a-b+) | 19 * | ||||
| G/G | C/C | le59/le59 | Le(a-b-) | 6 | ||||
| T/T | - | Sew/Sew | Weak secretor | T/T | C/C | Le/Le | Le(a+b+) | 9 |
| T/G | C/C | Le/le59 | Le(a+b+) | 5 | ||||
| G/G | C/C | le59/le59 | Le(a-b-) | 8 | ||||
| A/A | one copy | Se/sefus | Secretor | T/T | C/C | Le/Le | Le(a-b+) | 2 |
| T/G | C/C | Le/le59 | Le(a-b+) | 6 | ||||
| G/G | C/C | le59/le59 | Le(a-b-) | 1 | ||||
| T/T | one copy | Sew/sefus | Weak secretor | T/G | C/C | Le/le59 | Le(a+b+) | 2 |
| A/A | two copies | sefus/sefus | Non-secretor | T/G | C/C | Le/le59 | Le(a+b-) | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soejima, M.; Koda, Y. Estimation of Lewis Blood Group Status by Fluorescence Melting Curve Analysis in Simultaneous Genotyping of c.385A>T and Fusion Gene in FUT2 and c.59T>G and c.314C>T in FUT3. Diagnostics 2023, 13, 931. https://doi.org/10.3390/diagnostics13050931
Soejima M, Koda Y. Estimation of Lewis Blood Group Status by Fluorescence Melting Curve Analysis in Simultaneous Genotyping of c.385A>T and Fusion Gene in FUT2 and c.59T>G and c.314C>T in FUT3. Diagnostics. 2023; 13(5):931. https://doi.org/10.3390/diagnostics13050931
Chicago/Turabian StyleSoejima, Mikiko, and Yoshiro Koda. 2023. "Estimation of Lewis Blood Group Status by Fluorescence Melting Curve Analysis in Simultaneous Genotyping of c.385A>T and Fusion Gene in FUT2 and c.59T>G and c.314C>T in FUT3" Diagnostics 13, no. 5: 931. https://doi.org/10.3390/diagnostics13050931