
Identification and Experimental Validation of LINC00582 Associated with B Cell Immune and Development of Pulpitis: Bioinformatics and In Vitro Analysis
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Analysis
2.2. Functional Enrichment Analysis
2.3. Immune Cell Infiltration Analysis
2.4. Correlation Analysis between LncRNA and mRNA
2.5. Tissue Samples
2.6. Cell Culture and Transfection
2.7. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.8. Cell Counting Kit-8 (CCK-8) Assay
2.9. Lactate Dehydrogenase (LDH) Release Detection Assay
2.10. Transwell Assay of BALL-1 Cells
2.11. Transwell Assay of BALL-1 Cells
3. Results
3.1. Identification and Characteristics of LncRNAs between Normal and Pulpitis Tissue
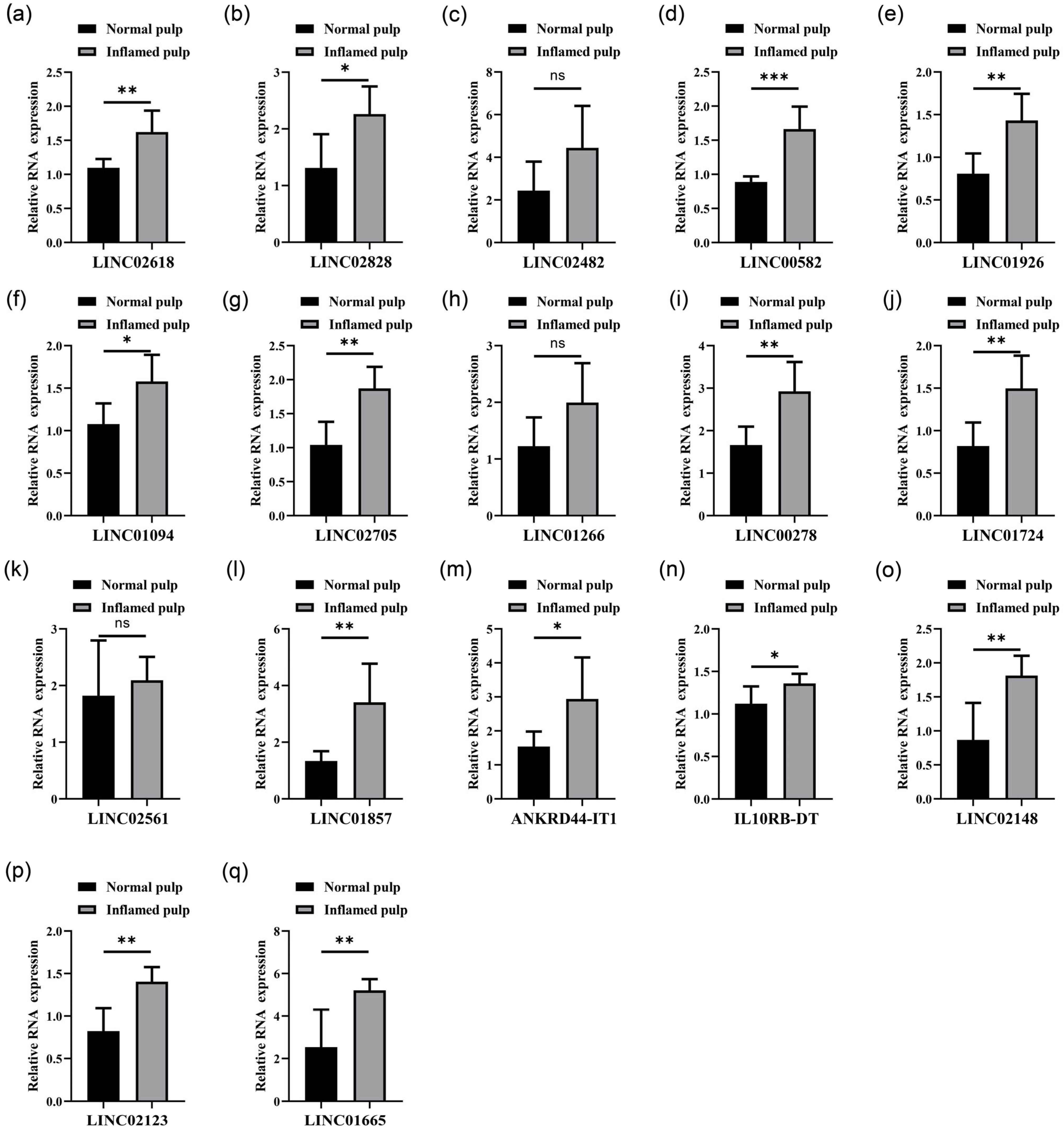
3.2. Analysis of LncRNAs Differentially Abundant in Normal Pulp Tissues Compared with Inflamed Pulp Tissues
3.3. Functional Annotation of Differentially Expressed Genes between Normal and Pulpitis Tissue
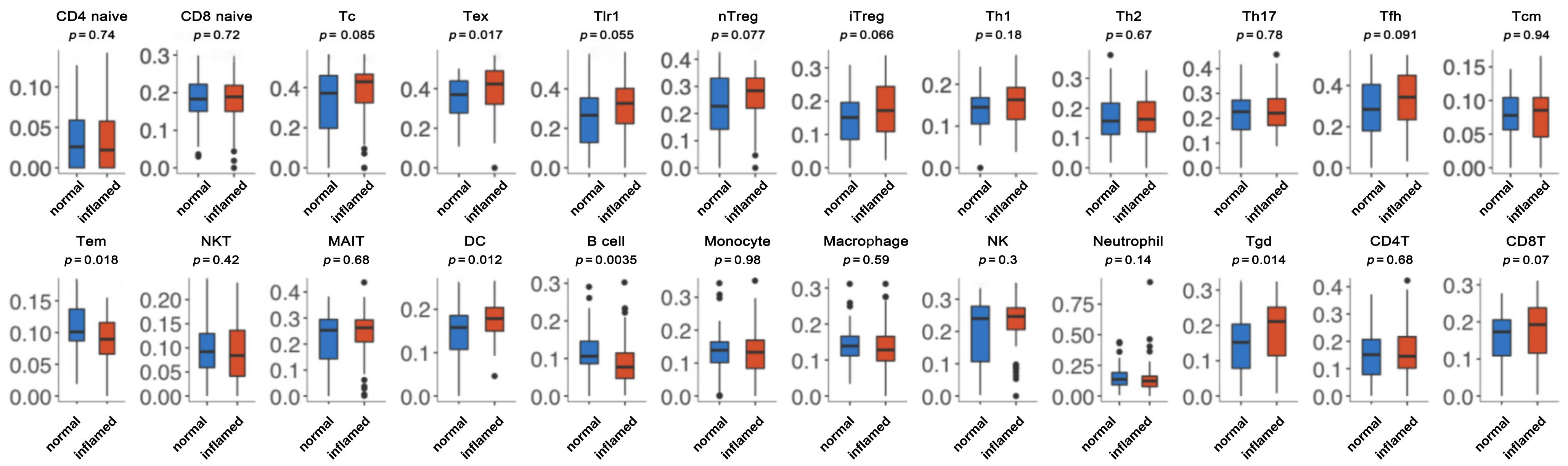
3.4. Immune Cell Abundance Identification between Normal and Inflamed Tissue
3.5. Correlation Analysis between LncRNAs and B Cell Activate Marker CD79B
3.6. Overexpression of LINC00582 Could Not Regulate Biological Function of HDPCs
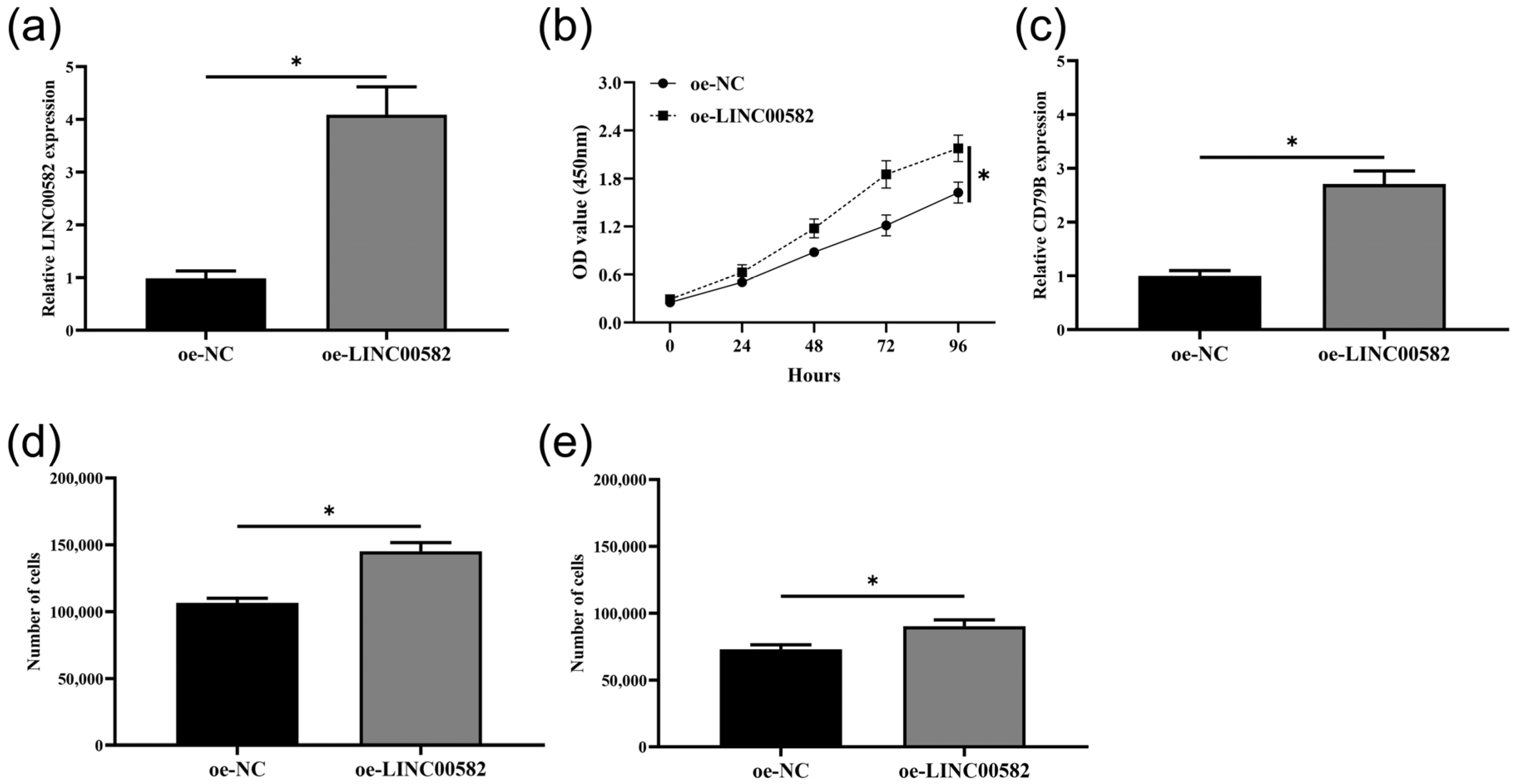
3.7. Overexpression of LINC00582 Promoted Proliferation, Migration, Invasion, and the Expression of CD79B in BALL-1 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leng, S.; Liu, L.; Xu, W.; Yang, F.; Du, J.; Ye, L.; Huang, D.; Zhang, L. Inflammation down regulates stromal cell-derived factor 1alpha in the early phase of pulpitis. Cytokine 2022, 158, 155983. [Google Scholar] [CrossRef] [PubMed]
- Gufran, K.; Mirza, M.B.; Robaian, A.; Alqahtani, A.S.; Alqhtani, N.R.; Alasqah, M.; Alsakr, A.M. A Prospective Clinical Study Evaluating the Efficacy of Intra-Ligamentary Anesthetic Solutions in Mandibular Molars Diagnosed as Symptomatic Irreversible Pulpitis with Symptomatic Apical Periodontitis. Healthcare 2022, 10, 1389. [Google Scholar] [CrossRef] [PubMed]
- Zhong, T.Y.; Zhang, Z.C.; Gao, Y.N.; Lu, Z.; Qiao, H.; Zhou, H.; Liu, Y. Loss of Wnt4 expression inhibits the odontogenic potential of dental pulp stem cells through JNK signaling in pulpitis. Am. J. Transl. Res. 2019, 11, 1819–1826. [Google Scholar] [PubMed]
- Yan, H.; Oshima, M.; Raju, R.; Raman, S.; Sekine, K.; Waskitho, A.; Inoue, M.; Inoue, M.; Baba, O.; Morita, T.; et al. Dentin-Pulp Complex Tissue Regeneration via Three-Dimensional Cell Sheet Layering. Tissue Eng. Part C Methods 2021, 27, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, Z.; Huang, J.; Liu, C. Therapeutic effect of one-time root canal treatment for irreversible pulpitis. J. Int. Med. Res. 2020, 48, 300060519879287. [Google Scholar] [CrossRef]
- Bamini, L.; Anand Sherwood, I.; Abbott, P.V.; Uthandakalaipandian, R.; Velu, V. Influence of anti-inflammatory irrigant on substance P expression for single-visit root canal treatment of teeth with irreversible pulpitis. Aust. Endod. J. 2020, 46, 73–81. [Google Scholar] [CrossRef]
- Bertossi, D.; Barone, A.; Iurlaro, A.; Marconcini, S.; De Santis, D.; Finotti, M.; Procacci, P. Odontogenic Orofacial Infections. J. Craniofac. Surg. 2017, 28, 197–202. [Google Scholar] [CrossRef]
- Ustiashvili, M.; Kordzaia, D.; Mamaladze, M.; Jangavadze, M.; Sanodze, L. Investigation of functional activity human dental pulp stem cells at acute and chronic pulpitis. Georgian Med. News 2014, 234, 19–24. [Google Scholar]
- Bhandi, S.; Alkahtani, A.; Mashyakhy, M.; Abumelha, A.S.; Albar, N.H.M.; Renugalakshmi, A.; Alkahtany, M.F.; Robaian, A.; Almeslet, A.S.; Patil, V.R.; et al. Effect of Ascorbic Acid on Differentiation, Secretome and Stemness of Stem Cells from Human Exfoliated Deciduous Tooth (SHEDs). J. Pers. Med. 2021, 11, 589. [Google Scholar] [CrossRef]
- Galler, K.M.; Weber, M.; Korkmaz, Y.; Widbiller, M.; Feuerer, M. Inflammatory Response Mechanisms of the Dentine-Pulp Complex and the Periapical Tissues. Int. J. Mol. Sci. 2021, 22, 1480. [Google Scholar] [CrossRef]
- Renard, E.; Gaudin, A.; Bienvenu, G.; Amiaud, J.; Farges, J.C.; Cuturi, M.C.; Moreau, A.; Alliot-Licht, B. Immune Cells and Molecular Networks in Experimentally Induced Pulpitis. J. Dent. Res. 2016, 95, 196–205. [Google Scholar] [CrossRef]
- Ghattas Ayoub, C.; Aminoshariae, A.; Bakkar, M.; Ghosh, S.; Bonfield, T.; Demko, C.; Montagnese, T.A.; Mickel, A.K. Comparison of IL-1beta, TNF-alpha, hBD-2, and hBD-3 Expression in the Dental Pulp of Smokers Versus Nonsmokers. J. Endod. 2017, 43, 2009–2013. [Google Scholar] [CrossRef] [PubMed]
- Kermeoglu, F.; Aksoy, U.; Sebai, A.; Savtekin, G.; Ozkayalar, H.; Sayiner, S.; Sehirli, A.O. Anti-Inflammatory Effects of Melatonin and 5-Methoxytryptophol on Lipopolysaccharide-Induced Acute Pulpitis in Rats. Biomed. Res. Int. 2021, 2021, 8884041. [Google Scholar] [CrossRef] [PubMed]
- Thanapati, S.; Ganu, M.; Giri, P.; Kulkarni, S.; Sharma, M.; Babar, P.; Ganu, A.; Tripathy, A.S. Impaired NK cell functionality and increased TNF-alpha production as biomarkers of chronic chikungunya arthritis and rheumatoid arthritis. Hum. Immunol. 2017, 78, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Shindo, S.; Kumagai, T.; Shirawachi, S.; Takeda, K.; Shiba, H. Semaphorin3A released from human dental pulp cells inhibits the increase in interleukin-6 and CXC chemokine ligand 10 production induced by tumor necrosis factor-alpha through suppression of nuclear factor-kappaB activation. Cell Biol. Int. 2021, 45, 238–244. [Google Scholar] [CrossRef]
- Lei, F.; Zhang, H.; Xie, X. Comprehensive analysis of an lncRNA-miRNA-mRNA competing endogenous RNA network in pulpitis. PeerJ 2019, 7, e7135. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Liu, L.; Yang, F.; Leng, S.; Zhang, L.; Huang, D. Identification of Immune-Related lncRNA Regulatory Network in Pulpitis. Dis. Markers 2022, 2022, 7222092. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chen, L.; Wu, J.; Lin, Z.; Huang, S. Long noncoding RNA MEG3 expressed in human dental pulp regulates LPS-Induced inflammation and odontogenic differentiation in pulpitis. Exp. Cell Res. 2021, 400, 112495. [Google Scholar] [CrossRef]
- Xia, L.; Wang, J.; Qi, Y.; Fei, Y.; Wang, D. Long Non-coding RNA PVT1 is Involved in the Pathological Mechanism of Pulpitis by Regulating miR-128-3p. Oral. Health Prev. Dent. 2022, 20, 263–270. [Google Scholar] [CrossRef]
- Wang, X.; Sun, H.; Hu, Z.; Mei, P.; Wu, Y.; Zhu, M. NUTM2A-AS1 silencing alleviates LPS-induced apoptosis and inflammation in dental pulp cells through targeting let-7c-5p/HMGB1 axis. Int. Immunopharmacol. 2021, 96, 107497. [Google Scholar] [CrossRef]
- Van de Vijfeijken, S.; Zigterman, B.G.R.; Helmers, R.; Dubois, L. The dentogenic inflammation, diagnosis, course and treatment. Ned. Tijdschr. Tandheelkd. 2022, 129, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Yong, D.; Cathro, P. Conservative pulp therapy in the management of reversible and irreversible pulpitis. Aust. Dent. J. 2021, 66 (Suppl. 1), S4–S14. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Z.; Li, W.; Tian, S. PECAM1 Combines With CXCR4 to Trigger Inflammatory Cell Infiltration and Pulpitis Progression Through Activating the NF-kappaB Signaling Pathway. Front. Cell Dev. Biol. 2020, 8, 593653. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhao, Y.; Wang, C.; Luo, H.; A, P.; Ye, L. Interleukin-17 plays a role in pulp inflammation partly by WNT5A protein induction. Arch. Oral. Biol. 2019, 103, 33–39. [Google Scholar] [CrossRef]
- Xin, B.; Lin, Y.; Tian, H.; Song, J.; Zhang, L.; Lv, J. Identification of Pulpitis-Related Potential Biomarkers Using Bioinformatics Approach. Comput. Math. Methods Med. 2021, 2021, 1808361. [Google Scholar] [CrossRef]
- Chen, M.; Zeng, J.; Yang, Y.; Wu, B. Diagnostic biomarker candidates for pulpitis revealed by bioinformatics analysis of merged microarray gene expression datasets. BMC Oral. Health 2020, 20, 279. [Google Scholar] [CrossRef]
- Hong, S.; Park, Y.H.; Lee, J.; Moon, J.; Kong, E.; Jeon, J.; Park, J.C.; Kim, H.R.; Kim, P. 3D Visualization of Dynamic Cellular Reaction of Pulpal CD11c+ Dendritic Cells against Pulpitis in Whole Murine Tooth. Int. J. Mol. Sci. 2021, 22, 2683. [Google Scholar] [CrossRef]
- Hahn, C.L.; Liewehr, F.R. Update on the adaptive immune responses of the dental pulp. J. Endod. 2007, 33, 773–781. [Google Scholar] [CrossRef]
- Lanzavecchia, A. Receptor-mediated antigen uptake and its effect on antigen presentation to class II-restricted T lymphocytes. Annu. Rev. Immunol. 1990, 8, 773–793. [Google Scholar] [CrossRef]
- Mix, E.; Meyer-Rienecker, H.; Hartung, H.P.; Zettl, U.K. Animal models of multiple sclerosis--potentials and limitations. Prog. Neurobiol. 2010, 92, 386–404. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Li, L.; Sad, S. Functions of CD8 T-cell subsets secreting different cytokine patterns. Semin. Immunol. 1997, 9, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Damoiseaux, J. Regulatory T cells: Back to the future. Neth. J. Med. 2006, 64, 4–9. [Google Scholar] [PubMed]
- Giardino, L.; Savoldi, E.; Pontieri, F.; Berutti, E. Russell bodies in dental pulp of permanent human teeth. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2004, 98, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Izumi, T.; Kobayashi, I.; Okamura, K.; Sakai, H. Immunohistochemical study on the immunocompetent cells of the pulp in human non-carious and carious teeth. Arch. Oral. Biol. 1995, 40, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Bednarczyk, M.; Medina-Montano, C.; Fittler, F.J.; Stege, H.; Roskamp, M.; Kuske, M.; Langer, C.; Vahldieck, M.; Montermann, E.; Tubbe, I.; et al. Complement-Opsonized Nano-Carriers Are Bound by Dendritic Cells (DC) via Complement Receptor (CR)3, and by B Cell Subpopulations via CR-1/2, and Affect the Activation of DC and B-1 Cells. Int. J. Mol. Sci. 2021, 22, 2869. [Google Scholar] [CrossRef]
- Lund, F.E.; Garvy, B.A.; Randall, T.D.; Harris, D.P. Regulatory roles for cytokine-producing B cells in infection and autoimmune disease. Curr. Dir. Autoimmun. 2005, 8, 25–54. [Google Scholar] [CrossRef]
- Hahn, C.L.; Falkler, W.A., Jr.; Siegel, M.A. A study of T and B cells in pulpal pathosis. J. Endod. 1989, 15, 20–26. [Google Scholar] [CrossRef]
- Speer, M.L.; Madonia, J.V.; Heuer, M.A. Quantitative evaluation of the immunocompetence of the dental pulp. J. Endod. 1977, 3, 418–423. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, L.; Deng, J.; Guo, B.; Li, F.; Wang, Y.; Wu, R.; Zhang, S.; Lu, J.; Zhou, Y. A Novel Micropeptide Encoded by Y-Linked LINC00278 Links Cigarette Smoking and AR Signaling in Male Esophageal Squamous Cell Carcinoma. Cancer Res. 2020, 80, 2790–2803. [Google Scholar] [CrossRef]
- Yang, C.; Cao, H.; Yang, J.W.; Wang, J.T.; Yu, M.M.; Wang, B.S. The ETS1-LINC00278 negative feedback loop plays a role in COL4A1/COL4A2 regulation in laryngeal squamous cell carcinoma. Neoplasma 2022, 69, 841–858. [Google Scholar] [CrossRef]
- Chen, M.; Zheng, Y.; Xie, J.; Zhen, E.; Zhou, X. Integrative profiling analysis identifies the oncogenic long noncoding RNA DUXAP8 in oral cancer. Anticancer Drugs 2020, 31, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Xu, Q.; Yin, J.; Wang, H.; Zhang, L. CircRNA, lncRNA, and mRNA profiles of umbilical cord blood exosomes from preterm newborns showing bronchopulmonary dysplasia. Eur. J. Pediatr. 2022, 181, 3345–3365. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.R.; Sun, W.; Zheng, K.; Zhu, W. LINC01857 promotes the development of gastric cancer by regulating microRNA-200b. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3648–3656. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, B.; Lu, C.L.; Wang, J.Y.; Gao, M.; Gao, W. LncRNA LINC01857 promotes cell growth and diminishes apoptosis via PI3K/mTOR pathway and EMT process by regulating miR-141-3p/MAP4K4 axis in diffuse large B-cell lymphoma. Cancer Gene Ther. 2021, 28, 1046–1057. [Google Scholar] [CrossRef]
- Hu, G.; Liu, N.; Wang, H.; Wang, Y.; Guo, Z. LncRNA LINC01857 promotes growth, migration, and invasion of glioma by modulating miR-1281/TRIM65 axis. J. Cell Physiol. 2019, 234, 22009–22016. [Google Scholar] [CrossRef]
- Liu, L.; Wang, T.; Huang, D.; Song, D. Comprehensive Analysis of Differentially Expressed Genes in Clinically Diagnosed Irreversible Pulpitis by Multiplatform Data Integration Using a Robust Rank Aggregation Approach. J. Endod. 2021, 47, 1365–1375. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Age (Years) | Diagnostic Information | Sample Information |
|---|---|---|---|
| Pulpitis tissue 1 | 26 | Irreversible pulpitis | Inflamed pulp tissue |
| Pulpitis tissue 2 | 27 | Irreversible pulpitis | Inflamed pulp tissue |
| Pulpitis tissue 3 | 29 | Irreversible pulpitis | Inflamed pulp tissue |
| Pulpitis tissue 4 | 25 | Irreversible pulpitis | Inflamed pulp tissue |
| Pulpitis tissue 5 | 27 | Irreversible pulpitis | Inflamed pulp tissue |
| Pulpitis tissue 6 | 29 | Irreversible pulpitis | Inflamed pulp tissue |
| Pulpitis tissue 7 | 24 | Irreversible pulpitis | Inflamed pulp tissue |
| Normal pulp tissue 1 | 25 | Healthy pulp tissue | Third molars extracted for orthodontic purposes |
| Normal pulp tissue 2 | 24 | Healthy pulp tissue | Third molars extracted for orthodontic purposes |
| Normal pulp tissue 3 | 22 | Healthy pulp tissue | Third molars extracted for orthodontic purposes |
| Normal pulp tissue 4 | 24 | Healthy pulp tissue | Third molars extracted for orthodontic purposes |
| Normal pulp tissue 5 | 25 | Healthy pulp tissue | Third molars extracted for orthodontic purposes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, W.; Hong, L.; Qian, Y. Identification and Experimental Validation of LINC00582 Associated with B Cell Immune and Development of Pulpitis: Bioinformatics and In Vitro Analysis. Diagnostics 2023, 13, 1678. https://doi.org/10.3390/diagnostics13101678
Gong W, Hong L, Qian Y. Identification and Experimental Validation of LINC00582 Associated with B Cell Immune and Development of Pulpitis: Bioinformatics and In Vitro Analysis. Diagnostics. 2023; 13(10):1678. https://doi.org/10.3390/diagnostics13101678
Chicago/Turabian StyleGong, Wenting, Lilin Hong, and Yi Qian. 2023. "Identification and Experimental Validation of LINC00582 Associated with B Cell Immune and Development of Pulpitis: Bioinformatics and In Vitro Analysis" Diagnostics 13, no. 10: 1678. https://doi.org/10.3390/diagnostics13101678