
The Revolution of Lateral Flow Assay in the Field of AMR Detection

 , , and
, , and
Abstract
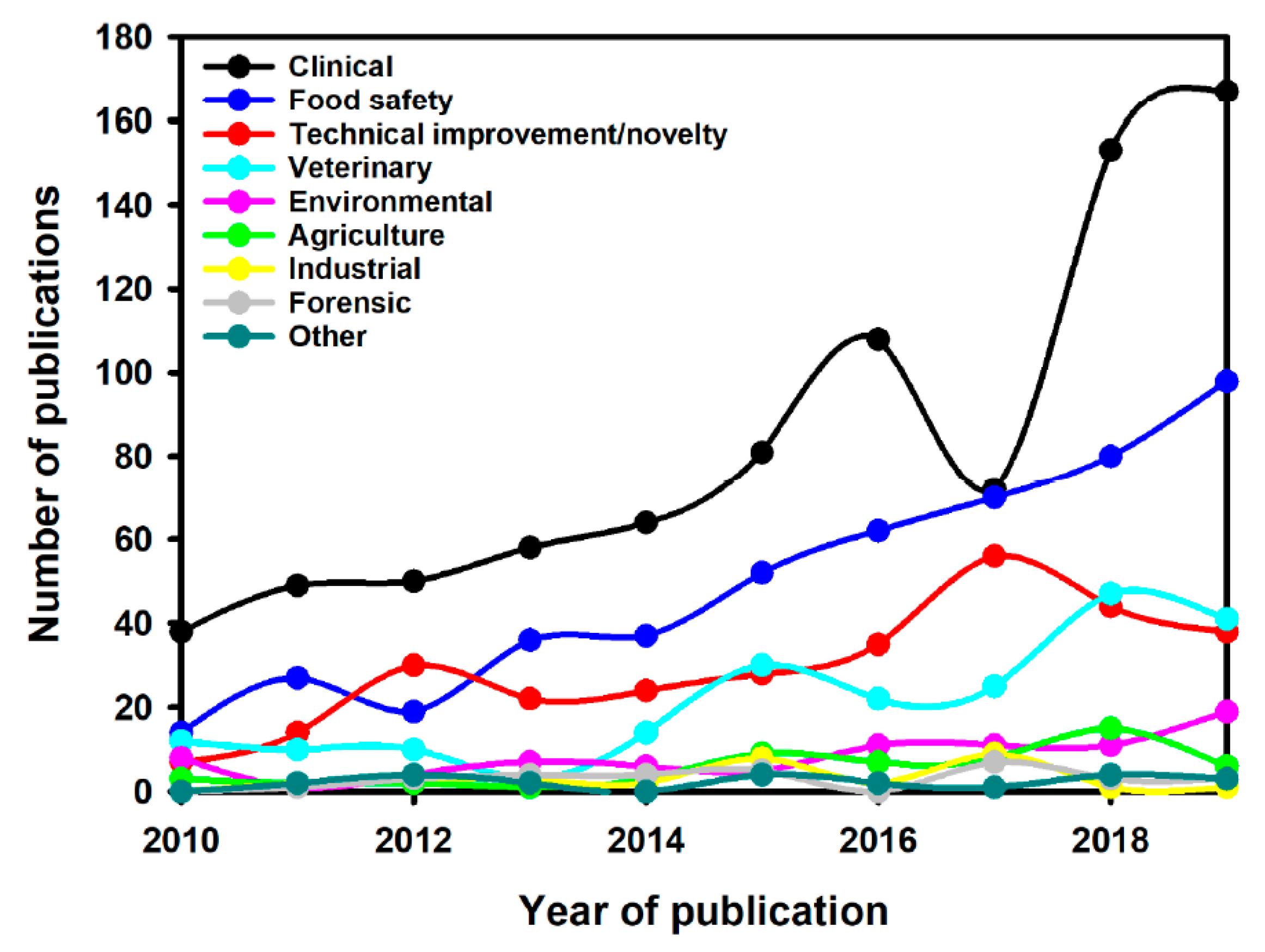
:1. Introduction
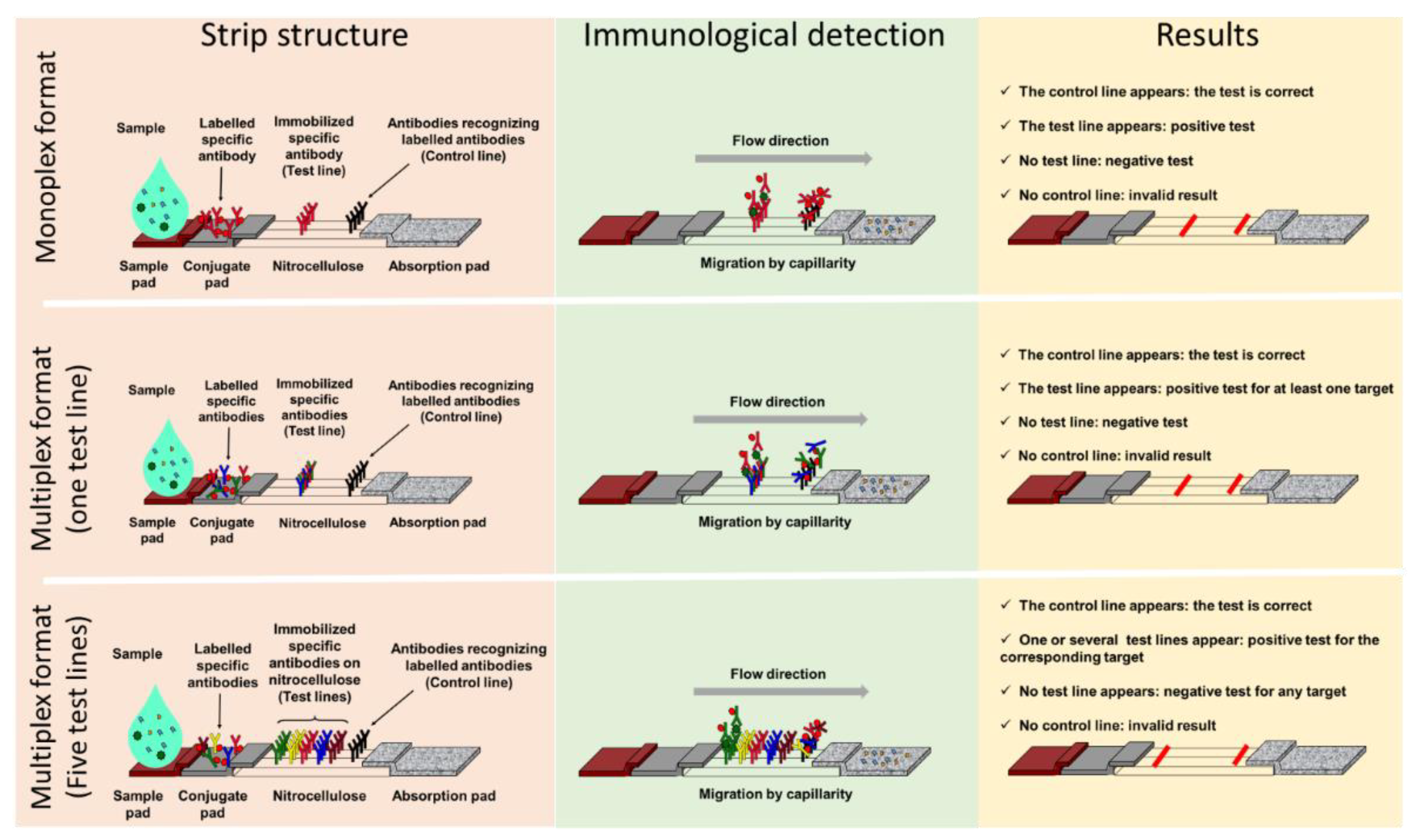
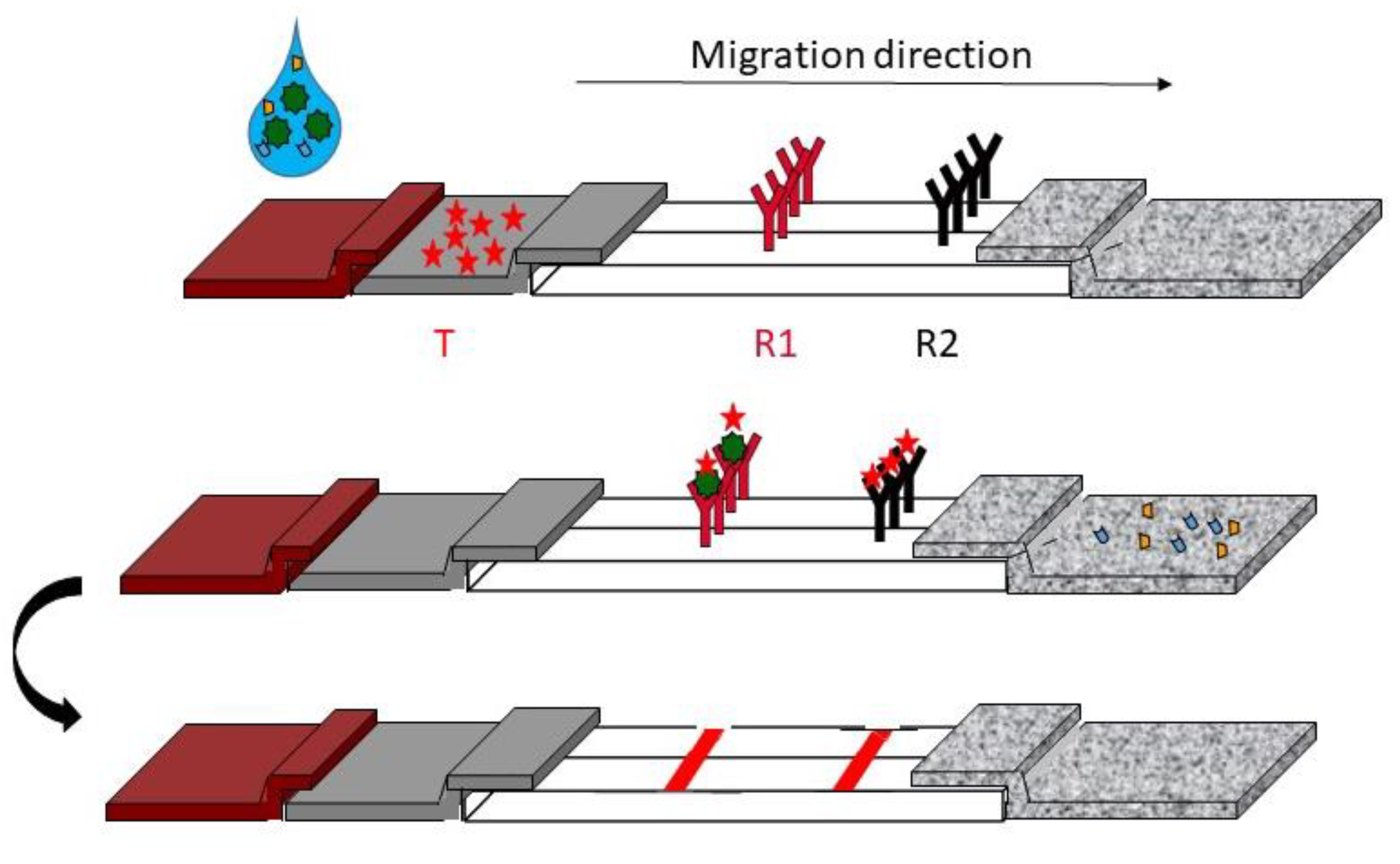
2. General Presentation of LFIA
2.1. Components and Principle
2.2. Advantages of LFIA
3. Classical Lateral Flow Assays and AMR
3.1. Monoplex LFIA to Address AMR Detection
3.2. Multiplex LFIA in the AMR Field
3.3. Limitations of LFIA in the Context of AMR
4. LFIA: A Phenotypic Method
4.1. Classical Phenotypic Methods
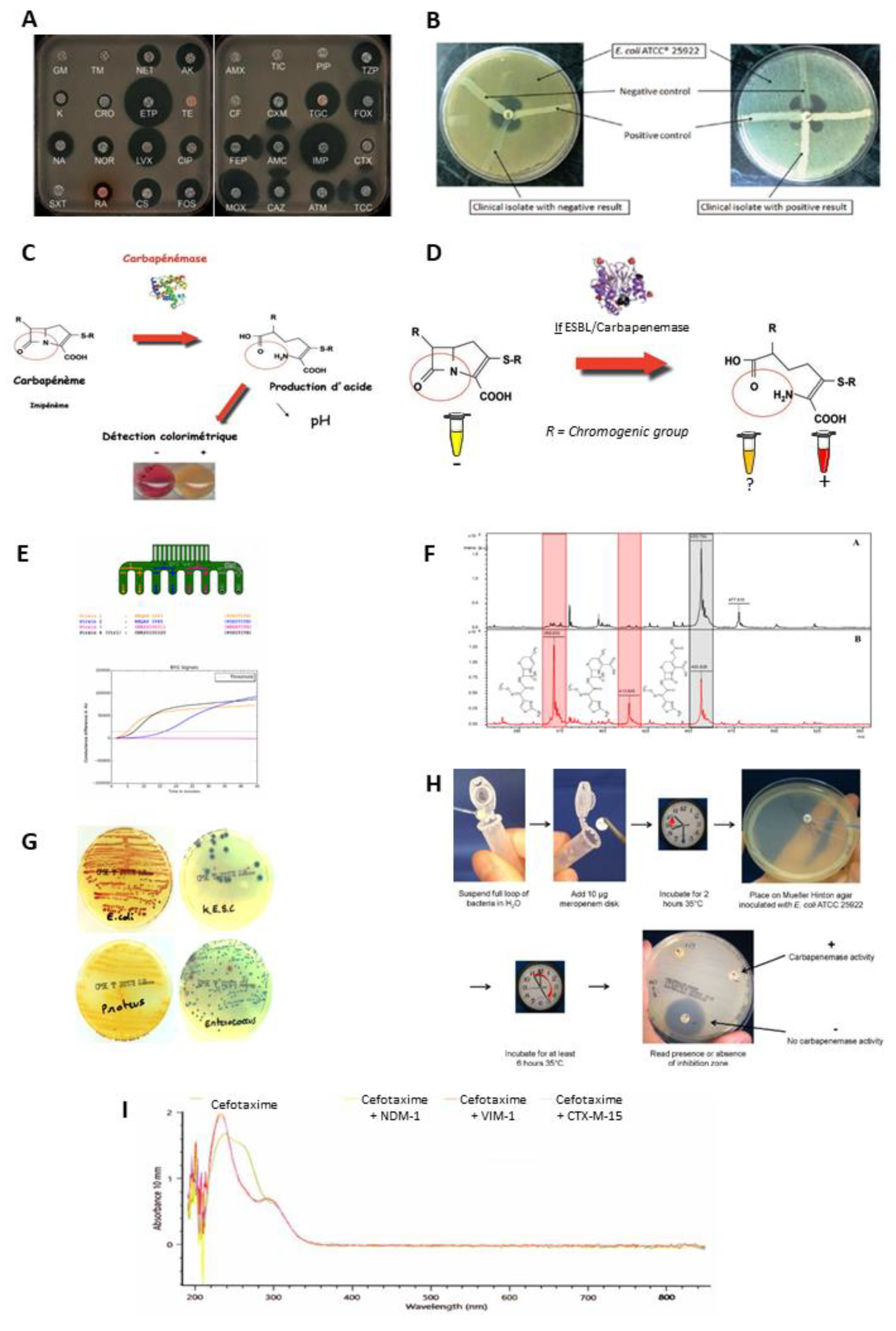
- The diffusion disc test (antibiogram, combined disc synergy test, inhibition test), is one of the reference methods [75,76,77,78]. The principle consists of placing the culture of bacteria in the presence of one or more antibiotics and observing the consequences on their development and survival in a Petri dish (Figure 3A).
- The modified Hodge test consists of inoculating Mueller–Hinton agar with the reference bacterial strain, wild type Escherichia coli ATCC 25,922, at 0.5 McFarland diluted 1:10. A disc containing carbapenem (meropenem or ertapenem) is placed in the centre of the agar. Colonies of the test strain are picked and plated in a line from the disc to the periphery of the plate. The presence of a carbapenemase is demonstrated by a cloverleaf indentation of Escherichia coli ATCC 25,922 that develops along the growth line of the isolate in the diffusion zone of the disc [79] (Figure 3B).
- The colorimetric tests detect β-lactamase activity via a color change in the reactive medium related to hydrolytic activity. This change in color is due to a biochemical change in the medium such as acidification. Here, three of them are presented: the Carba NP test, the β-Carba test, and the β-Lacta test [80,81,82] (Figure 3C,D).
- The electrochemical test is based on the analysis of conductivity variations within an electrode, composed of eight probes, coated with a conductive polymer, polyaniline. This variation is induced by changes in pH and redox potential related to an enzymatic hydrolysis reaction of imipenem [83] (Figure 3E).
- Mass spectrometry is used to detect the degradation of antibiotics by measuring their mass. Currently, among the different mass spectrometry technologies the MALDIO-TOF is the most used for this application. After ionization, the ionic molecules are accelerated in an electric field and projected towards a detector. This detector allows the ions to be separated and analysed according to their time of flight, which depends on their mass [84,85] (Figure 3F).
- The selective media tests are composed of chromogenic substances, rich nutrients, as well as specific antibiotic depending on the desired detection. Thus, they allow the identification of the strains involved through differential staining induced by the presence of characteristic enzymes [86] (Figure 3G).
- The carbapenem inactivation method consists of lysing the bacterial colonies to be tested in order to recover the suspension containing the possible β-lactamases. This lysate is divided into two separate tubes containing, respectively, a 10 µg meropenem disc (carbapenemase detection) and a 5 µg cefotaxime disc (ESBL detection). In parallel, two Mueller–Hinton agar plates were inoculated with an Escherichia coli ATCC 25,922 strain, known as a β-lactam sensitive strain. After incubation for 2 h at 36 °C, the meropenem and cefotaxime discs are transferred successively to the two plates. A further incubation is carried out for 4 h at 37 °C, followed by a reading and interpretation of the diameter of the inhibition zones. The absence of inhibition zones around the disc reflects degradation of the antibiotic during the first incubation, indicating hydrolytic activity of β-lactamases [87,88,89] (Figure 3G).

4.2. Evolution of LFIA in the Detection of Antibiotic Resistance
4.2.1. Detection of Enzymatic Activity Using LFIA
4.2.2. Enzymatic Activity Detection Coupled to Enzyme Detection Using LFIA
5. Direct Detection of Antibiotic-Resistant Bacteria in Clinical Samples
5.1. Direct Detection Using Centrifugation/Washing Steps
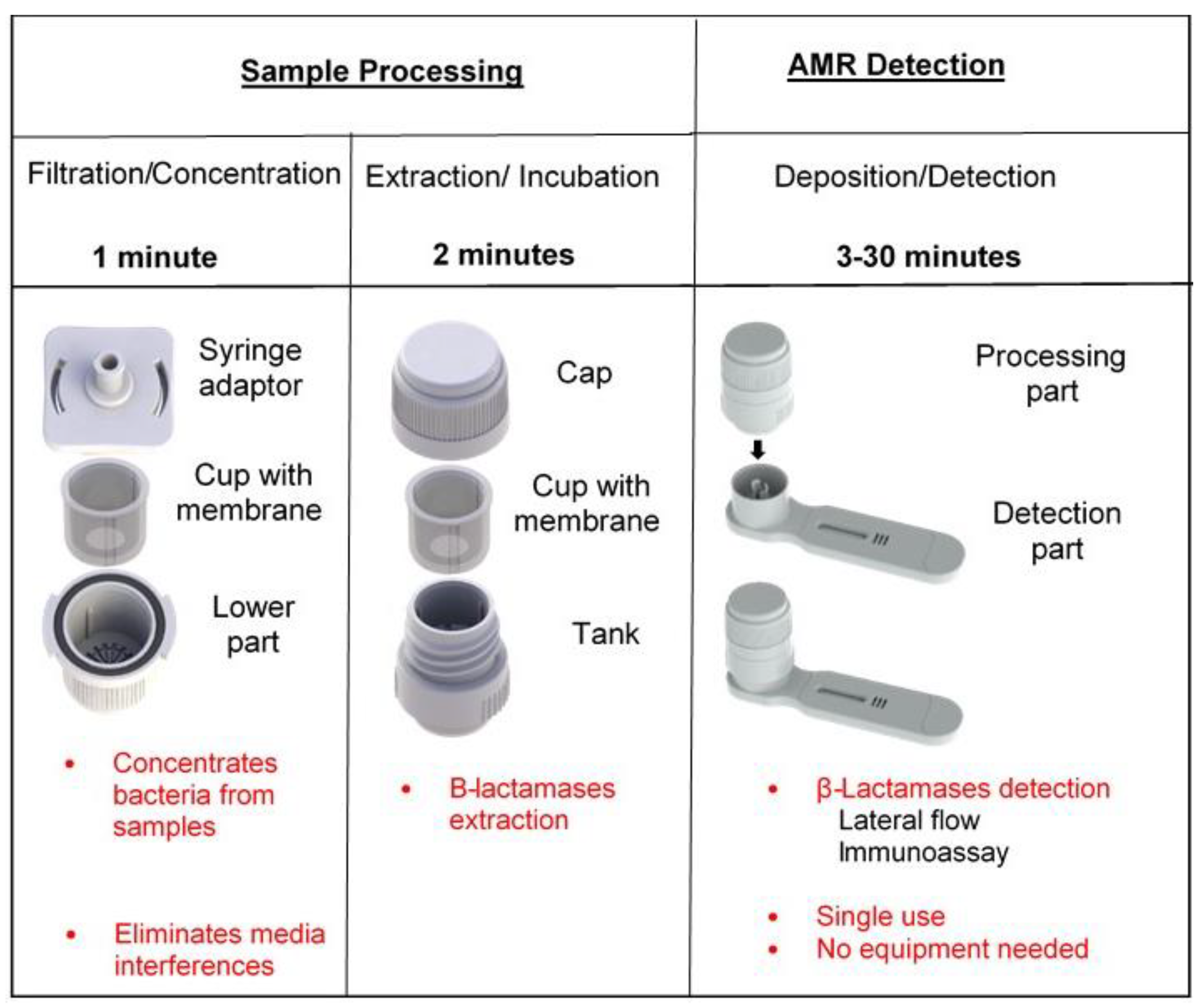
5.2. Direct Detection Using a Dedicated Device
6. Detection of Amplicons by LFIA
6.1. Polymerase Chain Reaction (PCR)
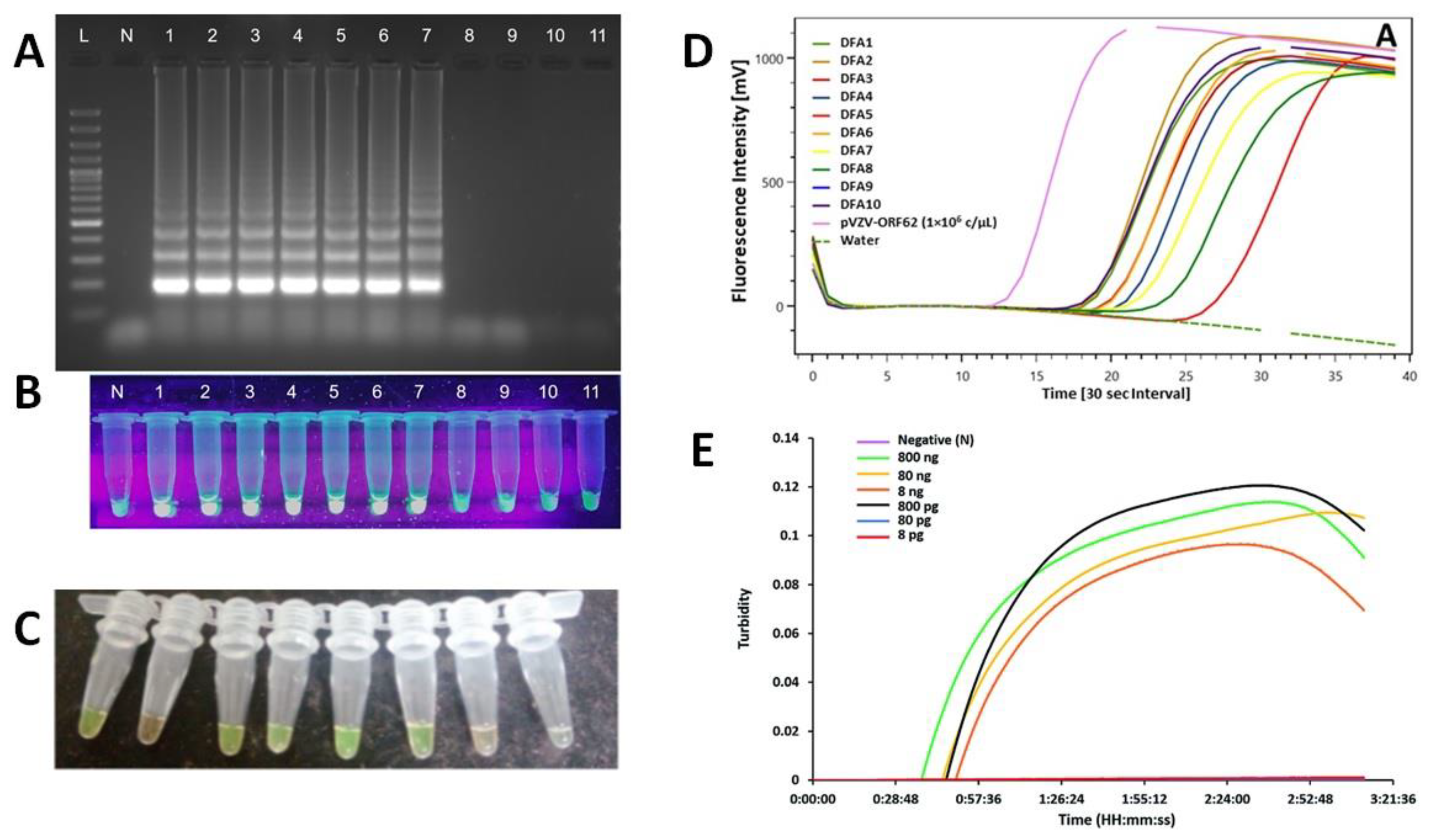
6.2. Loop-Mediated Isothermal Amplification (LAMP)
6.2.1. LAMP Detection Methods
6.2.2. LAMP Coupled to LFA Detection
6.2.3. Other Amplification Methods for AMR Detection
7. Conclusions
Funding
Conflicts of Interest
References
- Adedeji, W.A. The treasure called antibiotics. Ann. Ib. Postgrad. Med. 2016, 14, 56–57. [Google Scholar] [PubMed]
- Gould, K. Antibiotics: From Prehistory to the Present Day. J. Antimicrob. Chemother. 2016, 71, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Tangcharoensathien, V.; Chanvatik, S.; Sommanustweechai, A. Complex Determinants of Inappropriate Use of Antibiotics. Bull. World Health Organ. 2018, 96, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Golding, S.E.; Ogden, J.; Higgins, H.M. Shared Goals, Different Barriers: A Qualitative Study of UK Veterinarians’ and Farmers’ Beliefs About Antimicrobial Resistance and Stewardship. Front. Vet. Sci. 2019, 6, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Founou, R.C.; Founou, L.L.; Essack, S.Y. Clinical and Economic Impact of Antibiotic Resistance in Developing Countries: A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0189621. [Google Scholar] [CrossRef] [Green Version]
- White, A.; Hughes, J.M. Critical Importance of a One Health Approach to Antimicrobial Resistance. EcoHealth 2019, 16, 404–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, C.A.; Dominey-Howes, D.; Labbate, M. The Antimicrobial Resistance Crisis: Causes, Consequences, and Management. Front. Public Health 2014, 2, 145. [Google Scholar] [CrossRef]
- Zucca, M.; Savoia, D. The Post-Antibiotic Era: Promising Developments in the Therapy of Infectious Diseases. Int. J. Biomed. Sci. 2010, 6, 77–86. [Google Scholar]
- Naylor, N.R.; Atun, R.; Zhu, N.; Kulasabanathan, K.; Silva, S.; Chatterjee, A.; Knight, G.M.; Robotham, J.V. Estimating the Burden of Antimicrobial Resistance: A Systematic Literature Review. Antimicrob. Resist. Infect. Control 2018, 7, 58. [Google Scholar] [CrossRef]
- Ronat, J.-B.; Natale, A.; Kesteman, T.; Andremont, A.; Elamin, W.; Hardy, L.; Kanapathipillai, R.; Michel, J.; Langendorf, C.; Vandenberg, O.; et al. AMR in Low-Resource Settings: Médecins Sans Frontières Bridges Surveillance Gaps by Developing a Turnkey Solution, the Mini-Lab. Clin. Microbiol. Infect. 2021, 27, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Consumption and Resistance in Bacteria from Humans and Animals: Third Joint Inter-Agency Report on Integrated Analysis of Antimicrobial Agent Consumption and Occurrence of Antimicrobial Resistance in Bacteria from Humans and Food-Producing Animals in the EU/EEA: JIACRA III 2016–2018; European Food Safety Authority; European Centre for Disease Prevention and Control; European Medicines Agency: Stockholm, Sweden, 2021.
- Price, C.P. Regular Review: Point of Care Testing. BMJ 2001, 322, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Noster, J.; Thelen, P.; Hamprecht, A. Detection of Multidrug-Resistant Enterobacterales—From ESBLs to Carbapenemases. Antibiotics 2021, 10, 1140. [Google Scholar] [CrossRef] [PubMed]
- Dortet, L.; Tandé, D.; de Briel, D.; Bernabeu, S.; Lasserre, C.; Gregorowicz, G.; Jousset, A.B.; Naas, T. MALDI-TOF for the Rapid Detection of Carbapenemase-Producing Enterobacteriaceae: Comparison of the Commercialized MBT STAR®-Carba IVD Kit with Two in-House MALDI-TOF Techniques and the RAPIDEC® CARBA NP. J. Antimicrob. Chemother. 2018, 73, 2352–2359. [Google Scholar] [CrossRef] [Green Version]
- Naseri, M.; Ziora, Z.M.; Simon, G.P.; Batchelor, W. ASSURED-compliant Point-of-care Diagnostics for the Detection of Human Viral Infections. Rev. Med. Virol. 2021, 32, e2263. [Google Scholar] [CrossRef]
- Land, K.J.; Boeras, D.I.; Chen, X.-S.; Ramsay, A.R.; Peeling, R.W. REASSURED Diagnostics to Inform Disease Control Strategies, Strengthen Health Systems and Improve Patient Outcomes. Nat. Microbiol. 2019, 4, 46–54. [Google Scholar] [CrossRef]
- Tuena, C.; Semonella, M.; Fernández-Álvarez, J.; Colombo, D.; Cipresso, P. Predictive Precision Medicine: Towards the Computational Challenge. In P5 eHealth: An Agenda for the Health Technologies of the Future; Pravettoni, G., Triberti, S., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 71–86. ISBN 978-3-030-27993-6. [Google Scholar]
- Bahadır, E.B.; Sezgintürk, M.K. Lateral Flow Assays: Principles, Designs and Labels. TrAC Trends Anal. Chem. 2016, 82, 286–306. [Google Scholar] [CrossRef]
- Koczula, K.M.; Gallotta, A. Lateral Flow Assays. Essays Biochem. 2016, 60, 111–120. [Google Scholar] [CrossRef]
- Liu, Y.; Zhan, L.; Qin, Z.; Sackrison, J.; Bischof, J.C. Ultrasensitive and Highly Specific Lateral Flow Assays for Point-of-Care Diagnosis. ACS Nano 2021, 15, 3593–3611. [Google Scholar] [CrossRef]
- Di Nardo, F.; Chiarello, M.; Cavalera, S.; Baggiani, C.; Anfossi, L. Ten Years of Lateral Flow Immunoassay Technique Applications: Trends, Challenges and Future Perspectives. Sensors 2021, 21, 5185. [Google Scholar] [CrossRef] [PubMed]
- Posthuma-Trumpie, G.A.; Korf, J.; van Amerongen, A. Lateral Flow (Immuno)assay: Its Strengths, Weaknesses, Opportunities and Threats. A Literature Survey. Anal. Bioanal. Chem. 2009, 393, 569–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J. An Optimized Colorimetric Readout Method for Lateral Flow Immunoassays. Sensors 2018, 18, 4084. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Lu, L.; Rong, Z.; Wang, C.; Peng, Y.; Wang, F.; Wang, J.; Sun, M.; Dong, J.; Wang, D.; et al. Portable and Multiplexed Lateral Flow Immunoassay Reader Based on SERS for Highly Sensitive Point-of-Care Testing. Biosens. Bioelectron. 2020, 168, 112524. [Google Scholar] [CrossRef]
- Mühlberg, E.; Umstätter, F.; Kleist, C.; Domhan, C.; Mier, W.; Uhl, P. Renaissance of Vancomycin: Approaches for Breaking Antibiotic Resistance in Multidrug-Resistant Bacteria. Can. J. Microbiol. 2020, 66, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitao, T.; Miyoshi-Akiyama, T.; Shimada, K.; Tanaka, M.; Narahara, K.; Saito, N.; Kirikae, T. Development of an Immunochromatographic Assay for the Rapid Detection of AAC(6′)-Iae-Producing Multidrug-Resistant Pseudomonas Aeruginosa. J. Antimicrob. Chemother. 2010, 65, 1382–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshiro, S.; Tada, T.; Kameoka, Y.; Suzuki, K.; Ohmagari, N.; Miyoshi-Akiyama, T.; Kirikae, T. Development and Evaluation of Immunochromatography to Detect Gram-Negative Bacteria Producing ArmA 16S rRNA Methylase Responsible for Aminoglycoside Resistance. J. Microbiol. Methods 2015, 118, 159–163. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus Aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Wanchun, J.; Ohkura, T.; Murai, A.; Hayakawa, R.; Kinoshita, K.; Mizutani, M.; Okamoto, A.; Namikawa, T.; Ohta, M. Detection of Methicillin-Resistant Staphylococcus Aureus Using a Specific Anti-PBP2a Chicken IgY Antibody. Jpn. J. Infect. Dis. 2013, 66, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Amini, M.; Pourmand, M.R.; Faridi-Majidi, R.; Heiat, M.; Mohammad Nezhady, M.A.; Safari, M.; Noorbakhsh, F.; Baharifar, H. Optimising Effective Parameters to Improve Performance Quality in Lateral Flow Immunoassay for Detection of PBP2a in Methicillin-Resistant Staphylococcus Aureus (MRSA). J. Exp. Nanosci. 2020, 15, 266–279. [Google Scholar] [CrossRef]
- Cattoir, V.; Leclercq, R. Twenty-Five Years of Shared Life with Vancomycin-Resistant Enterococci: Is It Time to Divorce? J. Antimicrob. Chemother. 2013, 68, 731–742. [Google Scholar] [CrossRef]
- Shadel, B.N.; Puzniak, L.A.; Gillespie, K.N.; Lawrence, S.J.; Kollef, M.; Mundy, L.M. Surveillance for Vancomycin-Resistant Enterococci: Type, Rates, Costs, and Implications. Infect. Control Hosp. Epidemiol. 2006, 27, 1068–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prematunge, C.; MacDougall, C.; Johnstone, J.; Adomako, K.; Lam, F.; Robertson, J.; Garber, G. VRE and VSE Bacteremia Outcomes in the Era of Effective VRE Therapy: A Systematic Review and Meta-Analysis. Infect. Control Hosp. Epidemiol. 2016, 37, 26–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, J.K.; Cattoir, V.; Hegstad, K.; Sadowy, E.; Coque, T.M.; Westh, H.; Hammerum, A.M.; Schaffer, K.; Burns, K.; Murchan, S.; et al. Update on Prevalence and Mechanisms of Resistance to Linezolid, Tigecycline and Daptomycin in Enterococci in Europe: Towards a Common Nomenclature. Drug Resist. Updates 2018, 40, 25–39. [Google Scholar] [CrossRef]
- Oueslati, S.; Volland, H.; Cattoir, V.; Bernabeu, S.; Girlich, D.; Dulac, D.; Plaisance, M.; Laroche, M.; Dortet, L.; Simon, S.; et al. Development and Validation of a Lateral Flow Immunoassay for Rapid Detection of VanA-Producing Enterococci. J. Antimicrob. Chemother. 2021, 76, 146–151. [Google Scholar] [CrossRef]
- Oueslati, S.; Gonzalez, C.; Volland, H.; Cattoir, V.; Bernabeu, S.; Girlich, D.; Dulac, D.; Plaisance, M.; Boutigny, L.; Dortet, L.; et al. Rapid Detection of VanA/B-Producing Vancomycin-Resistant Enterococci Using Lateral Flow Immunoassay. Diagnosis 2021, 11, 1805. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A.; Munoz-Price, L.S. The New β-Lactamases. N. Engl. J. Med. 2005, 352, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Jacoby, G.A. Updated Functional Classification of β-Lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woerther, P.-L.; Burdet, C.; Chachaty, E.; Andremont, A. Trends in Human Fecal Carriage of Extended-Spectrum β-Lactamases in the Community: Toward the Globalization of CTX-M. Clin. Microbiol. Rev. 2013, 26, 744–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wareham, D.W.; Shah, R.; Betts, J.W.; Phee, L.M.; Momin, M.H.F.A. Evaluation of an Immunochromatographic Lateral Flow Assay (OXA-48 K -SeT) for Rapid Detection of OXA-48-Like Carbapenemases in Enterobacteriaceae. J. Clin. Microbiol. 2016, 54, 471–473. [Google Scholar] [CrossRef] [Green Version]
- Kitao, T.; Miyoshi-Akiyama, T.; Tanaka, M.; Narahara, K.; Shimojima, M.; Kirikae, T. Development of an Immunochromatographic Assay for Diagnosing the Production of IMP-Type Metallo-β-Lactamases That Mediate Carbapenem Resistance in Pseudomonas. J. Microbiol. Methods 2011, 87, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Notake, S.; Matsuda, M.; Tamai, K.; Yanagisawa, H.; Hiramatsu, K.; Kikuchi, K. Detection of IMP Metallo-β-Lactamase in Carbapenem-Nonsusceptible Enterobacteriaceae and Non-Glucose-Fermenting Gram-Negative Rods by Immunochromatography Assay. J. Clin. Microbiol. 2013, 51, 1762–1768. [Google Scholar] [CrossRef] [Green Version]
- Tada, T.; Sekiguchi, J.-I.; Watanabe, S.; Kuwahara-Arai, K.; Mizutani, N.; Yanagisawa, I.; Hishinuma, T.; Zan, K.N.; Mya, S.; Tin, H.H.; et al. Assessment of a Newly Developed Immunochromatographic Assay for NDM-Type Metallo-β-Lactamase Producing Gram-Negative Pathogens in Myanmar. BMC Infect. Dis. 2019, 19, 565. [Google Scholar] [CrossRef]
- Boutal, H.; Naas, T.; Devilliers, K.; Oueslati, S.; Dortet, L.; Bernabeu, S.; Simon, S.; Volland, H. Development and Validation of a Lateral Flow Immunoassay for Rapid Detection of NDM-Producing Enterobacteriaceae. J. Clin. Microbiol. 2017, 55, 2018–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Kasiakou, S.K.; Saravolatz, L.D. Colistin: The Revival of Polymyxins for the Management of Multidrug-Resistant Gram-Negative Bacterial Infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Ling, Z.; Yin, W.; Shen, Z.; Wang, Y.; Shen, J.; Walsh, T.R. Epidemiology of Mobile Colistin Resistance Genes Mcr-1 to Mcr-9. J. Antimicrob. Chemother. 2020, 75, 3087–3095. [Google Scholar] [CrossRef] [PubMed]
- Volland, H.; Dortet, L.; Bernabeu, S.; Boutal, H.; Haenni, M.; Madec, J.-Y.; Robin, F.; Beyrouthy, R.; Naas, T.; Simon, S. Development and Multicentric Validation of a Lateral Flow Immunoassay for Rapid Detection of MCR-1-Producing Enterobacteriaceae. J. Clin. Microbiol. 2019, 57, e01454-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernabeu, S.; Ratnam, K.C.; Boutal, H.; Gonzalez, C.; Vogel, A.; Devilliers, K.; Plaisance, M.; Oueslati, S.; Malhotra-Kumar, S.; Dortet, L.; et al. A Lateral Flow Immunoassay for the Rapid Identification of CTX-M-Producing Enterobacterales from Culture Plates and Positive Blood Cultures. Diagnostics 2020, 10, 764. [Google Scholar] [CrossRef] [PubMed]
- Bianco, G.; Boattini, M.; Iannaccone, M.; Cavallo, R.; Costa, C. Evaluation of the NG-Test CTX-M MULTI Immunochromatographic Assay for the Rapid Detection of CTX-M Extended-Spectrum-β-Lactamase Producers from Positive Blood Cultures. J. Hosp. Infect. 2020, 105, 341–343. [Google Scholar] [CrossRef]
- Nishida, S.; Nakagawa, M.; Ouchi, Y.; Sakuma, C.; Nakajima, Y.; Shimizu, H.; Shibata, T.; Kurosawa, Y.; Maruyama, T.; Okumura, C.J.; et al. A Rabbit Monoclonal Antibody-Mediated Lateral Flow Immunoassay for Rapid Detection of CTX-M Extended-Spectrum β-Lactamase-Producing Enterobacterales. Int. J. Biol. Macromol. 2021, 185, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anfossi, L.; Di Nardo, F.; Cavalera, S.; Giovannoli, C.; Baggiani, C. Multiplex Lateral Flow Immunoassay: An Overview of Strategies towards High-Throughput Point-of-Need Testing. Biosensors 2018, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Hamprecht, A.; Vehreschild, J.J.; Seifert, H.; Saleh, A. Rapid Detection of NDM, KPC and OXA-48 Carbapenemases Directly from Positive Blood Cultures Using a New Multiplex Immunochromatographic Assay. PLoS ONE 2018, 13, e0204157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glupczynski, Y.; Evrard, S.; Huang, T.-D.; Bogaerts, P. Evaluation of the RESIST-4 K-SeT Assay, a Multiplex Immunochromatographic Assay for the Rapid Detection of OXA-48-Like, KPC, VIM and NDM Carbapenemases. J. Antimicrob. Chemother. 2019, 74, 1284–1287. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Kang, D.; Kim, D. Performance Evaluation of the Newly Developed In Vitro Rapid Diagnostic Test for Detecting OXA-48-Like, KPC-, NDM-, VIM- and IMP-Type Carbapenemases: The RESIST-5 O.K.N.V.I. Multiplex Lateral Flow Assay. Antibiotics 2021, 10, 460. [Google Scholar] [CrossRef] [PubMed]
- El Kettani, A.; Maaloum, F.; Nzoyikorera, N.; Khalis, M.; Katfy, K.; Belabbes, H.; Zerouali, K. Evaluation of the Performances of the Rapid Test RESIST-5 O.O.K.N.V Used for the Detection of Carbapenemases-Producing Enterobacterales. Antibiotics 2021, 10, 953. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Guo, Y.; Peng, M.; Shi, Q.; Wu, S.; Yang, Y.; Zheng, Y.; Yin, D.; Hu, F. Evaluation of the Immunochromatographic NG-Test Carba 5, RESIST-5 O.O.K.N.V., and IMP K-SeT for Rapid Detection of KPC-, NDM-, IMP-, VIM-Type, and OXA-48-like Carbapenemase Among Enterobacterales. Front. Microbiol. 2021, 11, 3296. [Google Scholar] [CrossRef]
- Boutal, H.; Vogel, A.; Bernabeu, S.; Devilliers, K.; Creton, E.; Cotellon, G.; Plaisance, M.; Oueslati, S.; Dortet, L.; Jousset, A.; et al. A Multiplex Lateral Flow Immunoassay for the Rapid Identification of NDM-, KPC-, IMP- and VIM-Type and OXA-48-like Carbapenemase-Producing Enterobacteriaceae. J Antimicrob Chemother 2018, 73, 909–915. [Google Scholar] [CrossRef]
- Hopkins, K.L.; Meunier, D.; Naas, T.; Volland, H.; Woodford, N. Evaluation of the NG-Test CARBA 5 Multiplex Immunochromatographic Assay for the Detection of KPC, OXA-48-Like, NDM, VIM and IMP Carbapenemases. J. Antimicrob. Chemother. 2018, 73, 3523–3526. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.; Hopkins, K.L.; Meunier, D.; Woodford, N. Evaluation of Three Commercial Assays for Rapid Detection of Genes Encoding Clinically Relevant Carbapenemases in Cultured Bacteria. J. Antimicrob. Chemother. 2015, 70, 1338–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volland, H.; Girlich, D.; Laguide, M.; Gonzalez, C.; Paris, V.; Laroche, M.; Oueslati, S.; Dortet, L.; Simon, S.; Naas, T. Improvement of the Immunochromatographic NG-Test Carba 5 Assay for the Detection of IMP Variants Previously Undetected. Antimicrob. Agents Chemother. 2019, 64, e01940-19. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Fournier, D.; Emeraud, C.; Triponney, P.; Plésiat, P.; Naas, T.; Dortet, L. Evaluation of the Immunochromatographic NG-Test Carba 5 for Rapid Identification of Carbapenemase in Nonfermenters. Antimicrob. Agents Chemother. 2019, 63, e00968-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodendoerfer, E.; Keller, P.M.; Mancini, S. Rapid Identification of NDM-, KPC-, IMP-, VIM- and OXA-48-like Carbapenemase-Producing Enterobacteriales from Blood Cultures by a Multiplex Lateral Flow Immunoassay. J. Antimicrob. Chemother. 2019, 74, 1749–1751. [Google Scholar] [CrossRef] [PubMed]
- Giordano, L.; Fiori, B.; D’Inzeo, T.; Parisi, G.; Liotti, F.M.; Menchinelli, G.; De Angelis, G.; De Maio, F.; Luzzaro, F.; Sanguinetti, M.; et al. Simplified Testing Method for Direct Detection of Carbapenemase-Producing Organisms from Positive Blood Cultures Using the NG-Test Carba 5 Assay. Antimicrob. Agents Chemother. 2019, 63, e00550-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takissian, J.; Bonnin, R.A.; Naas, T.; Dortet, L. NG-Test Carba 5 for Rapid Detection of Carbapenemase-Producing Enterobacterales from Positive Blood Cultures. Antimicrob. Agents Chemother. 2019, 63, e00011-19. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, S.; Ledeboer, N.A.; Westblade, L.F.; Burnham, C.-A.D.; Faron, M.L.; Bergman, Y.; Yee, R.; Mesich, B.; Gerstbrein, D.; Wallace, M.A.; et al. Evaluation of NG-Test Carba 5 for Rapid Phenotypic Detection and Differentiation of Five Common Carbapenemase Families: Results of a Multicenter Clinical Evaluation. J. Clin. Microbiol. 2020, 58, e00344-20. [Google Scholar] [CrossRef]
- Huang, Y.-T.; Kuo, Y.-W.; Lee, N.-Y.; Tien, N.; Liao, C.-H.; Teng, L.-J.; Ko, W.-C.; Hsueh, P.-R. Evaluating NG-Test CARBA 5 Multiplex Immunochromatographic and Cepheid Xpert CARBA-R Assays among Carbapenem-Resistant Enterobacterales Isolates Associated with Bloodstream Infection. Microbiol. Spectr. 2022, 10, e01728-21. [Google Scholar] [CrossRef]
- Liu, Z.; Bai, L.; Liu, J.; Lei, J.; Gao, X.; Tenover, F.C.; Lei, K.; Tang, Y.-W.; Geng, Y.; He, A. Parallel Validation of the NG-Test Carba 5 and the Xpert Carba-R for Detection and Characterization of Carbapenem-Resistant Enterobacterales Causing Bloodstream Infections. J. Mol. Diagn. 2021, 23, 1007–1014. [Google Scholar] [CrossRef]
- Yoon, J.; Kim, C.H.; Yoon, S.-Y.; Lim, C.S.; Lee, C.K. Application of a Multiplex Immunochromatographic Assay for Rapid Identification of Carbapenemases in a Clinical Microbiology Laboratory: Performance and Turn-around-Time Evaluation of NG-Test Carba 5. BMC Microbiol. 2021, 21, 260. [Google Scholar] [CrossRef]
- Zhu, Y.; Jia, P.; Li, X.; Wang, T.; Zhang, J.; Zhang, G.; Duan, S.; Kang, W.; Xu, Y.; Yang, Q. Carbapenemase Detection by NG-Test CARBA 5—a Rapid Immunochromatographic Assay in Carbapenem-Resistant Enterobacterales Diagnosis. Ann. Transl. Med. 2021, 9, 769. [Google Scholar] [CrossRef]
- Baeza, L.L.; Pfennigwerth, N.; Greissl, C.; Göttig, S.; Saleh, A.; Stelzer, Y.; Gatermann, S.G.; Hamprecht, A. Comparison of Five Methods for Detection of Carbapenemases in Enterobacterales with Proposal of a New Algorithm. Clin. Microbiol. Infect. 2019, 25, 1286.e9–1286.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andryukov, B.G. Six Decades of Lateral Flow Immunoassay: From Determining Metabolic Markers to Diagnosing COVID-19. AIMS Microbiol. 2020, 6, 280–304. [Google Scholar] [CrossRef] [PubMed]
- Giske, C.G.; Martinez-Martinez, L.; Canton, R.; Stefani, S.; Skov, R.; Glupczynski, Y.; Nordmann, P.; Wootton, M.; Miriagou, V.; Skov Simonsen, G.; et al. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance; EUCAST—European Committee on Antimicrobial Susceptibility Testing: Växjö, Sweden, 2017; p. 43. [Google Scholar]
- Jorgensen, J.H.; Ferraro, M.J. Antimicrobial Susceptibility Testing: A Review of General Principles and Contemporary Practices. Clin. Infect. Dis. 2009, 49, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Drieux, L.; Brossier, F.; Sougakoff, W.; Jarlier, V. Phenotypic Detection of Extended-Spectrum Beta-Lactamase Production in Enterobacteriaceae: Review and Bench Guide. Clin. Microbiol. Infect. 2008, 14 (Suppl. 1), 90–103. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Poirel, L. Résistances aux antibiotiques émergentes et importantes chez les bactéries Gram négatif: Épidémiologie, aspects théoriques et détection. Rev. Méd. Suisse 2014, 10, 902–907. [Google Scholar]
- Cury, A.P.; Andreazzi, D.; Maffucci, M.; Caiaffa-Junior, H.H.; Rossi, F. The Modified Hodge Test Is a Useful Tool for Ruling out Klebsiella Pneumoniae Carbapenemase. Clinics 2012, 67, 1427–1431. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L.; Dortet, L. Rapid Detection of Carbapenemase-Producing Enterobacteriaceae. Emerg. Infect. Dis. 2012, 18, 1503–1507. [Google Scholar] [CrossRef] [Green Version]
- Shinde, S.; Gupta, R.; Raut, S.S.; Nataraj, G.; Mehta, P.R. Carba NP as a Simpler, Rapid, Cost-Effective, and a More Sensitive Alternative to Other Phenotypic Tests for Detection of Carbapenem Resistance in Routine Diagnostic Laboratories. J. Lab. Physic. 2017, 9, 100–103. [Google Scholar] [CrossRef]
- Vrioni, G.; Mamali, V.; Zarkotou, O.; Voulgari, E.; Dimitroulia, E.; Themeli-Digalaki, K.; Pournaras, S.; Tsakris, A. Performance of the β LACTATM Test for Rapid Detection of Expanded-Spectrum Cephalosporin-Non-Susceptible Enterobacteriaceae. J. Glob. Antimicrob. Resist. 2017, 10, 285–288. [Google Scholar] [CrossRef]
- Bogaerts, P.; Oueslati, S.; Meunier, D.; Nonhoff, C.; Yunus, S.; Massart, M.; Denis, O.; Woodford, N.; Hopkins, K.L.; Naas, T.; et al. Multicentre Evaluation of the BYG Carba v2.0 Test, a Simplified Electrochemical Assay for the Rapid Laboratory Detection of Carbapenemase-Producing Enterobacteriaceae. Sci. Rep. 2017, 7, 9937. [Google Scholar] [CrossRef] [Green Version]
- Hrabák, J.; Chudácková, E.; Walková, R. Matrix-Assisted Laser Desorption Ionization-Time of Flight (Maldi-Tof) Mass Spectrometry for Detection of Antibiotic Resistance Mechanisms: From Research to Routine Diagnosis. Clin. Microbiol. Rev. 2013, 26, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparbier, K.; Schubert, S.; Weller, U.; Boogen, C.; Kostrzewa, M. Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry-Based Functional Assay for Rapid Detection of Resistance against β-Lactam Antibiotics. J. Clin. Microbiol. 2012, 50, 927–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, J.D. A Decade of Development of Chromogenic Culture Media for Clinical Microbiology in an Era of Molecular Diagnostics. Clin. Microbiol. Rev. 2017, 30, 449–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, S.; Correa, A.; Hernández-Gómez, C.; De La Cadena, E.; Pallares, C.; Villegas, M.V. Detection of Carbapenemase-Producing Pseudomonas Aeruginosa: Evaluation of the Carbapenem Inactivation Method (CIM). Enferm. Infect. Microbiol. Clin. 2019, 37, 648–651. [Google Scholar] [CrossRef]
- Jing, X.; Zhou, H.; Min, X.; Zhang, X.; Yang, Q.; Du, S.; Li, Y.; Yu, F.; Jia, M.; Zhan, Y.; et al. The Simplified Carbapenem Inactivation Method (sCIM) for Simple and Accurate Detection of Carbapenemase-Producing Gram-Negative Bacilli. Front. Microbiol. 2018, 9, 2391. [Google Scholar] [CrossRef]
- Muntean, M.-M.; Muntean, A.-A.; Gauthier, L.; Creton, E.; Cotellon, G.; Popa, M.I.; Bonnin, R.A.; Naas, T. Evaluation of the Rapid Carbapenem Inactivation Method (rCIM): A Phenotypic Screening Test for Carbapenemase-Producing Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Laraki, N.; Franceschini, N.; Rossolini, G.M.; Santucci, P.; Meunier, C.; de Pauw, E.; Amicosante, G.; Frère, J.M.; Galleni, M. Biochemical Characterization of the Pseudomonas Aeruginosa 101/1477 Metallo-β-Lactamase IMP-1 Produced by Escherichia Coli. Antimicrob. Agents Chemother. 1999, 43, 902–906. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, D.; Akeda, Y.; Sugawara, Y.; Sakamoto, N.; Yamamoto, N.; Shanmugakani, R.K.; Ishihara, T.; Shintani, A.; Tomono, K.; Hamada, S. Establishment of a Dual-Wavelength Spectrophotometric Method for Analysing and Detecting Carbapenemase-Producing Enterobacteriaceae. Sci. Rep. 2018, 8, 15689. [Google Scholar] [CrossRef]
- Datta, S.; Dey, R.; Dey, J.B.; Ghosh, S. A Comparative Study of Modified Hodge Test and Carba NP Test for Detecting Carbapenemase Production in Gram-Negative Bacteria. Med. J. Dr. D. Y. Patil Univ. 2017, 10, 365. [Google Scholar] [CrossRef]
- Bogaerts, P.; Yunus, S.; Massart, M.; Huang, T.-D.; Glupczynski, Y. Evaluation of the BYG Carba Test, a New Electrochemical Assay for Rapid Laboratory Detection of Carbapenemase-Producing Enterobacteriaceae. J. Clin. Microbiol. 2016, 54, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Oviaño, M.; Rodicio, M.R.; Heinisch, J.J.; Rodicio, R.; Bou, G.; Fernández, J. Analysis of the Degradation of Broad-Spectrum Cephalosporins by OXA-48-Producing Enterobacteriaceae Using MALDI-TOF MS. Microorganisms 2019, 7, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milieu de Culture. Wikipédia 2021. Available online: https://fr.wikipedia.org/w/index.php?title=Milieu_de_culture&oldid=185004580 (accessed on 19 June 2022).
- Van der Zwaluw, K.; de Haan, A.; Pluister, G.; Bootsma, H.; de Neeling, A.D.; Schouls, L. The Carbapenem Inactivation Method (CIM), a Simple and Low-Cost Alternative for the Carba NP Test to Assess Phenotypic Carbapenemase Activity in Gram-Negative Rods. PLoS ONE 2015, 10, e0213690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre-Quiñonero, A.; Martínez-Martínez, L. Non-Molecular Detection of Carbapenemases in Enterobacteriaceae Clinical Isolates. J. Infect. Chemother. 2017, 23, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choquet, M.; Guiheneuf, R.; Castelain, S.; Cattoir, V.; Auzou, M.; Pluquet, E.; Decroix, V. Comparison of MALDI-ToF MS with the Rapidec Carba NP Test for the Detection of Carbapenemase-Producing Enterobacteriaceae. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Moguet, C.; Gonzalez, C.; Sallustrau, A.; Gelhaye, S.; Naas, T.; Simon, S.; Volland, H. Detection of Expanded-Spectrum Cephalosporin Hydrolysis by Lateral Flow Immunoassay. Microb. Biotechnol. 2021, 15, 603–612. [Google Scholar] [CrossRef]
- Gonzalez, C.; Moguet, C.; Chalin, A.; Oueslati, S.; Dortet, L.; Simon, S.; Volland, H.; Naas, T. Comparison of Three Expanded-Spectrum Cephalosporin Hydrolysis Assays and the NG-Test CTX-M Multi Assay That Detects All CTX-M-Like Enzymes. Diagnostics 2022, 12, 197. [Google Scholar] [CrossRef]
- Moguet, C.; Gonzalez, C.; Naas, T.; Simon, S.; Volland, H. Multiplex Lateral Flow Immunoassay for the Detection of Expanded-Spectrum Hydrolysis and CTX-M Enzymes. Diagnostics 2022, 12, 190. [Google Scholar] [CrossRef]
- Hitchens, A.P.; Leikind, M.C. The Introduction of Agar-Agar into Bacteriology. J. Bacteriol. 1939, 37, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Kawanishi, T.; Shiraishi, T.; Okano, Y.; Sugawara, K.; Hashimoto, M.; Maejima, K.; Komatsu, K.; Kakizawa, S.; Yamaji, Y.; Hamamoto, H.; et al. New Detection Systems of Bacteria Using Highly Selective Media Designed by SMART: Selective Medium-Design Algorithm Restricted by Two Constraints. PLoS ONE 2011, 6, e16512. [Google Scholar] [CrossRef]
- Oviaño, M.; de la Ramírez, C.L.; Barbeyto, L.P.; Bou, G. Rapid Direct Detection of Carbapenemase-Producing Enterobacteriaceae in Clinical Urine Samples by MALDI-TOF MS Analysis. J. Antimicrob. Chemother. 2017, 72, 1350–1354. [Google Scholar] [CrossRef]
- Fleurbaaij, F.; Goessens, W.; van Leeuwen, H.C.; Kraakman, M.E.M.; Bernards, S.T.; Hensbergen, P.J.; Kuijper, E.J. Direct Detection of Extended-Spectrum Beta-Lactamases (CTX-M) from Blood Cultures by LC-MS/MS Bottom-up Proteomics. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1621–1628. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Hamprecht, A. Systematic Comparison of Four Methods for Detection of Carbapenemase-Producing Enterobacterales Directly from Blood Cultures. J. Clin. Microbiol. 2019, 57, e00709-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, S.; Ching, J.; Xi, L.; Hughes, S.J.; Chang, R.; Wong, W.; McMillan, W.; Gooding, W.E.; McCarty, K.S.; Chestney, M.; et al. Technology for Automated, Rapid, and Quantitative PCR or Reverse Transcription-PCR Clinical Testing. Clin. Chem. 2005, 51, 882–890. [Google Scholar] [CrossRef]
- Hagan, K.A.; Reedy, C.R.; Uchimoto, M.L.; Basu, D.; Engel, D.A.; Landers, J.P. An Integrated, Valveless System for Microfluidic Purification and Reverse Transcription-PCR Amplification of RNA for Detection of Infectious Agents. Lab. Chip. 2011, 11, 957–961. [Google Scholar] [CrossRef]
- Xu, G.; Hsieh, T.-M.; Lee, D.Y.S.; Ali, E.M.; Xie, H.; Looi, X.L.; Koay, E.S.-C.; Li, M.-H.; Ying, J.Y. A Self-Contained All-in-One Cartridge for Sample Preparation and Real-Time PCR in Rapid Influenza Diagnosis. Lab. Chip. 2010, 10, 3103–3111. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-K.; Kim, H.S.; Pinto, N.; Jeon, J.; D’Souza, R.; Kim, M.S.; Choi, J.Y.; Yong, D.; Jeong, S.H.; Lee, K. Xpert CARBA-R Assay for the Detection of Carbapenemase-Producing Organisms in Intensive Care Unit Patients of a Korean Tertiary Care Hospital. Ann. Lab. Med. 2016, 36, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-L.; Luo, Q.-B.; Xiao, S.-S.; Lin, Z.-H.; Liu, Y.-L.; Han, M.-Y.; Zhong, J.-H.; Ji, T.-X.; Guo, X.-G. Evaluation of GeneXpert vanA/vanB in the Early Diagnosis of Vancomycin-Resistant Enterococci Infection. PLoS Negl. Trop. Dis. 2021, 15, e0009869. [Google Scholar] [CrossRef]
- Bianco, G.; Boattini, M.; van Asten, S.A.V.; Iannaccone, M.; Zanotto, E.; Zaccaria, T.; Bernards, A.T.; Cavallo, R.; Costa, C. RESIST-5 O.O.K.N.V. and NG-Test Carba 5 Assays for the Rapid Detection of Carbapenemase-Producing Enterobacterales from Positive Blood Cultures: A Comparative Study. J. Hosp. Infect. 2020, 105, 162–166. [Google Scholar] [CrossRef]
- Wareham, D.W.; Phee, L.M.; Abdul Momin, M.H.F. Direct Detection of Carbapenem Resistance Determinants in Clinical Specimens Using Immunochromatographic Lateral Flow Devices. J. Antimicrob. Chemother. 2018, 73, 1997–1998. [Google Scholar] [CrossRef]
- Baer, D.; Azrad, M.; Saleh, N.; Peretz, A. Detection of Carbapenem-Resistant Enterobacterales in Simulated Blood Culture in 15 Minutes. Life 2021, 11, 145. [Google Scholar] [CrossRef]
- Boattini, M.; Bianco, G.; Iannaccone, M.; Ghibaudo, D.; Almeida, A.; Cavallo, R.; Costa, C. Fast-Track Identification of CTX-M-Extended-Spectrum-β-Lactamase- and Carbapenemase-Producing Enterobacterales in Bloodstream Infections: Implications on the Likelihood of Deduction of Antibiotic Susceptibility in Emergency and Internal Medicine Departments. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- Vasilakopoulou, A.; Karakosta, P.; Vourli, S.; Kalogeropoulou, E.; Pournaras, S. Detection of KPC, NDM and VIM-Producing Organisms Directly from Rectal Swabs by a Multiplex Lateral Flow Immunoassay. Microorganisms 2021, 9, 942. [Google Scholar] [CrossRef] [PubMed]
- Fauconnier, C.; Dodemont, M.; Depouhon, A.; Anantharajah, A.; Verroken, A.; Rodriguez-Villalobos, H. Lateral Flow Immunochromatographic Assay for Rapid Screening of Faecal Carriage of Carbapenemase-Producing Enterobacteriaceae. J. Antimicrob. Chemother. 2019, 74, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Vasala, A.; Hytönen, V.P.; Laitinen, O.H. Modern Tools for Rapid Diagnostics of Antimicrobial Resistance. Front. Cell. Infect. Microbiol. 2020, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Volkmann, H.; Schwartz, T.; Bischoff, P.; Kirchen, S.; Obst, U. Detection of Clinically Relevant Antibiotic-Resistance Genes in Municipal Wastewater Using Real-Time PCR (TaqMan). J. Microbiol. Methods 2004, 56, 277–286. [Google Scholar] [CrossRef] [PubMed]
- PCR en Microbiologie: De l’Amplification de l’ADN à l’Interprétation du Résultat. Available online: https://www.revmed.ch/revue-medicale-suisse/2007/revue-medicale-suisse-106/pcr-en-microbiologie-de-l-amplification-de-l-adn-a-l-interpretation-du-resultat (accessed on 28 June 2021).
- Singh, J.; Birbian, N.; Sinha, S.; Goswami, A. Reaction (pcr, P.C. Review Article A Critical Review on PCR, Its Types and Applications. Int. J. Adv. Res. Biol. Sci. 2014, 1, 65–80. [Google Scholar]
- Multiplex PCR Assay for Simultaneous Detection of Nine Clinically Relevant Antibiotic Resistance Genes in Staphylococcus Aureus. Available online: https://journals.asm.org/doi/epub/10.1128/JCM.41.9.4089-4094.2003 (accessed on 16 September 2021).
- Aktas, G.B.; Wichers, J.H.; Skouridou, V.; van Amerongen, A.; Masip, L. Nucleic Acid Lateral Flow Assays Using a Conjugate of a DNA Binding Protein and Carbon Nanoparticles. Microchim. Acta 2019, 186, 426. [Google Scholar] [CrossRef]
- Kostoulas, P.; Eusebi, P.; Hartnack, S. Diagnostic Accuracy Estimates for COVID-19 Real-Time Polymerase Chain Reaction and Lateral Flow Immunoassay Tests With Bayesian Latent-Class Models. Am. J. Epidemiol. 2021, 190, 1689–1695. [Google Scholar] [CrossRef]
- Pecchia, S.; Da Lio, D. Development of a Rapid PCR-Nucleic Acid Lateral Flow Immunoassay (PCR-NALFIA) Based on rDNA IGS Sequence Analysis for the Detection of Macrophomina Phaseolina in Soil. J. Microbiol. Methods 2018, 151, 118–128. [Google Scholar] [CrossRef]
- Moers, A.P.H.A.; Hallett, R.L.; Burrow, R.; Schallig, H.D.F.H.; Sutherland, C.J.; van Amerongen, A. Detection of Single-Nucleotide Polymorphisms in Plasmodium Falciparum by PCR Primer Extension and Lateral Flow Immunoassay. Antimicrob. Agents Chemother. 2015, 59, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, V.K.; Bakthavathsalam, P.; Bergquist, P.L.; Sunna, A. Smartphone Detection of Antibiotic Resistance Using Convective PCR and a Lateral Flow Assay. Sens. Actuators B Chem. 2019, 298, 126849. [Google Scholar] [CrossRef]
- Kievits, T.; van Gemen, B.; van Strijp, D.; Schukkink, R.; Dircks, M.; Adriaanse, H.; Malek, L.; Sooknanan, R.; Lens, P. NASBATM Isothermal Enzymatic in Vitro Nucleic Acid Amplification Optimized for the Diagnosis of HIV-1 Infection. J. Virol. Methods 1991, 35, 273–286. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.Q. Rolling Replication of Short DNA Circles. Proc. Natl. Acad. Sci. USA 1995, 92, 4641–4645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, G.T.; Fraiser, M.S.; Schram, J.L.; Little, M.C.; Nadeau, J.G.; Malinowski, D.P. Strand Displacement Amplification—An Isothermal, in Vitro DNA Amplification Technique. Nucl. Acids Res. 1992, 20, 1691–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA Detection Using Recombination Proteins. PLoS Biol. 2006, 4, e204. [Google Scholar] [CrossRef]
- Vincent, M.; Xu, Y.; Kong, H. Helicase-Dependent Isothermal DNA Amplification. EMBO Rep. 2004, 5, 795–800. [Google Scholar] [CrossRef]
- Notomi, T. Loop-Mediated Isothermal Amplification of DNA. Nucl. Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [Green Version]
- Wong, Y.-P.; Othman, S.; Lau, Y.-L.; Radu, S.; Chee, H.-Y. Loop-Mediated Isothermal Amplification (LAMP): A Versatile Technique for Detection of Micro-Organisms. J. Appl. Microbiol. 2018, 124, 626–643. [Google Scholar] [CrossRef] [Green Version]
- Loop-Mediated Isothermal Amplification NEB. Available online: https://international.neb.com/applications/dna-amplification-pcr-and-qpcr/isothermal-amplification/loop-mediated-isothermal-amplification-lamp (accessed on 2 September 2021).
- Liu, D.; He, W.; Jiang, M.; Zhao, B.; Ou, X.; Liu, C.; Xia, H.; Zhou, Y.; Wang, S.; Song, Y.; et al. Development of a Loop-Mediated Isothermal Amplification Coupled Lateral Flow Dipstick Targeting erm(41) for Detection of Mycobacterium Abscessus and Mycobacterium Massiliense. AMB Express 2019, 9, 11. [Google Scholar] [CrossRef]
- Foo, P.C.; Nurul Najian, A.B.; Muhamad, N.A.; Ahamad, M.; Mohamed, M.; Yean Yean, C.; Lim, B.H. Loop-Mediated Isothermal Amplification (LAMP) Reaction as Viable PCR Substitute for Diagnostic Applications: A Comparative Analysis Study of LAMP, Conventional PCR, Nested PCR (nPCR) and Real-Time PCR (qPCR) Based on Entamoeba Histolytica DNA Derived from Faecal Sample. BMC Biotechnol. 2020, 20, 34. [Google Scholar] [CrossRef]
- Mori, Y.; Kitao, M.; Tomita, N.; Notomi, T. Real-Time Turbidimetry of LAMP Reaction for Quantifying Template DNA. J. Biochem. Biophys. Methods 2004, 59, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Nancy, K.; Lillian, W.; Wilkinson, M.; Claire, M.; Luna, K.; Dorcas, W.; Rosemary, G.; Japheth, L.; Christine, I.; Emily, W.; et al. Optimization of a Loop-Mediated Isothermal Amplification Assay as a Point-of-Care Tool for the Detection of Wuchereria Bancrofti in Human Blood in Tana River Delta, Kenya. J. Parasitol. Res. 2021, 2021, e6650870. [Google Scholar] [CrossRef] [PubMed]
- Gadkar, V.J.; Goldfarb, D.M.; Gantt, S.; Tilley, P.A.G. Real-Time Detection and Monitoring of Loop Mediated Amplification (LAMP) Reaction Using Self-Quenching and De-Quenching Fluorogenic Probes. Sci. Rep. 2018, 8, 5548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ma, K.; Yi, X.; Xiong, L.; Wang, Y.; Li, S. The Rapid and Visual Detection of Methicillin-Susceptible and Methicillin-Resistant Staphylococcus Aureus Using Multiplex Loop-Mediated Isothermal Amplification Linked to a Nanoparticle-Based Lateral Flow Biosensor. Antimicrob. Resist. Infect. Control 2020, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Tang, F.; Liu, E.; Liu, X.; Xu, H.; Wang, Y.; Song, Y.; Liang, J. Development of a Loop-Mediated Isothermal Amplification Assay Combined with a Nanoparticle-Based Lateral Flow Biosensor for Rapid Detection of Plasmid-Mediated Colistin Resistance Gene Mcr-1. PLoS ONE 2021, 16, e0249582. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Li, D.; Xu, J.; Ye, C. Detection of Nucleic Acids and Elimination of Carryover Contamination by Using Loop-Mediated Isothermal Amplification and Antarctic Thermal Sensitive Uracil-DNA-Glycosylase in a Lateral Flow Biosensor: Application to the Detection of Streptococcus Pneumoniae. Microchim. Acta 2018, 185, 212. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, N.; Xu, Y.; Huang, K.; Luo, Y.; Xu, W. Point-of-Care and Visual Detection of P. Aeruginosa and Its Toxin Genes by Multiple LAMP and Lateral Flow Nucleic Acid Biosensor. Biosens. Bioelectron. 2016, 81, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Njiru, Z.K. Rapid and Sensitive Detection of Human African Trypanosomiasis by Loop-Mediated Isothermal Amplification Combined with a Lateral-Flow Dipstick. Diagn. Microbiol. Infect. Dis. 2011, 69, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, Y.T.; Lee, J.W.; Kim, D.H.; Seo, T.S. An Integrated Direct Loop-Mediated Isothermal Amplification Microdevice Incorporated with an Immunochromatographic Strip for Bacteria Detection in Human Whole Blood and Milk without a Sample Preparation Step. Biosens. Bioelectron. 2016, 79, 273–279. [Google Scholar] [CrossRef]
- Loop Mediated Isothermal Amplification (LAMP)—Lateral Flow. Available online: https://www.milenia-biotec.com/en/isothermal-amplification-lateral-flow/ (accessed on 5 May 2021).
- Gong, L.; Liu, E.; Che, J.; Li, J.; Liu, X.; Xu, H.; Liang, J. Multiple Cross Displacement Amplification Coupled With Gold Nanoparticles-Based Lateral Flow Biosensor for Detection of the Mobilized Colistin Resistance Gene Mcr-1. Front. Cell. Infect. Microbiol. 2019, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, Q.; Guo, J.; Li, Y.; Chen, Q. Identification of Methicillin-Resistant Staphylococcus Aureus (MRSA) Using Simultaneous Detection of mecA, Nuc, and femB by Loop-Mediated Isothermal Amplification (LAMP). Curr. Microbiol. 2017, 74, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.R.; Stedtfeld, R.D.; Waseem, H.; Stedtfeld, T.; Upham, B.; Khalife, W.; Etchebarne, B.; Hughes, M.; Tiedje, J.M.; Hashsham, S.A. Implications of Direct Amplification for Measuring Antimicrobial Resistance Using Point-of-Care Devices. Anal. Methods 2017, 9, 1229–1241. [Google Scholar] [CrossRef] [PubMed]
- Alamolhoda, S.Z.; Zarghami, N.; Kahroba, H.; Mehdipour, A.; Pourhassan-Moghaddam, M.; Jahanban-Esfahlan, R.; Milani, M. Isothermal Amplification of Nucleic Acids Coupled with Nanotechnology and Microfluidic Platforms for Detecting Antimicrobial Drug Resistance and Beyond. Adv. Pharm. Bull. 2021, 12, 58–76. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests | Products | Time | Intrinsic Performance (Variable between Studies) | Information Provided | Easy to Implement | Main Limitation(s) |
|---|---|---|---|---|---|---|
| Hydrolysis of β-lactams | Antibiotic susceptibility test (from bioMérieux, Servilab…) | 16–24 h | High | Points to ESBL or carbapenemases | Little expertise needed | Accumulation of large amount of information |
| Chromogenic media (from bioMérieux, Oxoïd, CHROMagar) | 16–24 h | Medium to high | Points to ESBL or carbapenemases | Little expertise needed | Non-detection of enzymes with low activity | |
| Modified Hodge Test * | 16–24 h | Medium to high | Points to ESBL or carbapenemases | Medium expertise needed | Recurrent FP appearance + Difficulty in detecting MBLs | |
| Carba NP (from bioMérieux, Rosco Diagnostics) | 2 h | Medium to high | Points to carbapenemases | Medium expertise needed | No standardisation Faulty interpretation (FN possible) | |
| Blue Carba (from Rosco Diagnostics) | 2 h | High | Points to carbapenemases | Medium expertise needed | No standardisation Distorted interpretation (FN possible) | |
| β-Carba (from Bio-Rad) | 0.5 h | High | Points to carbapenemases | Little expertise needed | Incubation >0.5 h for strains with 0XA-48 enzymes | |
| ESBL NDP (from Rosco Diagnostics) | 2 h | High | Points to ESBL | Medium expertise needed | No standardisation Faulty interpretation (FN possible) | |
| β-Lacta (from Bio-Rad) | 0.25 h | High | Points to ESBL | Little expertise needed | Overexpression of AmpC can lead to → FP | |
| Mass spectrometry (Bruker, BioMérieux, Beckman Coulter) | 0.5–3 h | High | Points to ESBL or carbapenemases | Equipment needed + significant expertise needed | Visualisation of degradation products sometimes problematic | |
| UV spectroscopy * | 1 h | High | Points to ESBL or carbapenemases | Equipment needed + significant expertise needed | Interference present + standardisation of OD difficult | |
| BYG Carba * | 0.5 h | High | Points to carbapenemases | Equipment needed + significant expertise needed | No commercialised kit, nor evaluation of the technique | |
| Carbapenem inactivation method * | 6 h | Medium to high | Points to ESBL or carbapenemases | Little expertise needed | Variability of inhibition zones → FN | |
| Inhibition of β-lactamases | Combined discs (from Rosco Diagnostics, Mast Groups) | 16–24 h | High | Points to ESBL or carbapenemases | Little expertise needed | Variable antibiotic distribution + some inhibitors not very effective |
| Double synergy (from bioMérieux, Servilab…) | 16–24 h | High | Points to ESBL or carbapenemases | Little expertise needed | Questionable zones of interpretation + some inhibitors not very effective | |
| E-Test (from Rosco Diagnostics, bioMérieux) | 16–24 h | High | Points to ESBL or carbapenemases | Little expertise needed | Difficulty in detecting OXA-48 type enzymes + some inhibitors not very effective | |
| Lateral flow iiimmunoassays | RESIST (from Coris) | 0.25 h | High | Four of the five major carbapenemases in two tests (VIM, OXA-48, NDM, KPC) | Little expertise needed | Does not detect IMP enzymes or all new variants + use of 2 tests |
| NG-Test (from NG-Biotech) | 0.25 h | High | All five carbapenemases in one test + CTX-M | Little expertise needed | Does not detect all new variants |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boutal, H.; Moguet, C.; Pommiès, L.; Simon, S.; Naas, T.; Volland, H. The Revolution of Lateral Flow Assay in the Field of AMR Detection. Diagnostics 2022, 12, 1744. https://doi.org/10.3390/diagnostics12071744
Boutal H, Moguet C, Pommiès L, Simon S, Naas T, Volland H. The Revolution of Lateral Flow Assay in the Field of AMR Detection. Diagnostics. 2022; 12(7):1744. https://doi.org/10.3390/diagnostics12071744
Chicago/Turabian StyleBoutal, Hervé, Christian Moguet, Lilas Pommiès, Stéphanie Simon, Thierry Naas, and Hervé Volland. 2022. "The Revolution of Lateral Flow Assay in the Field of AMR Detection" Diagnostics 12, no. 7: 1744. https://doi.org/10.3390/diagnostics12071744