
miRNAs and Their Gene Targets—A Clue to Differentiate Pregnancies with Small for Gestational Age Newborns, Intrauterine Growth Restriction, and Preeclampsia

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics of Patients with PE, IUGR, and SGA and Healthy Pregnant Women
2.2. miRNA Expression Signature in Placenta and Placenta Bed Tissue Samples
2.3. Protein Expression Profiles in Placenta and Placenta Bed Tissue Samples
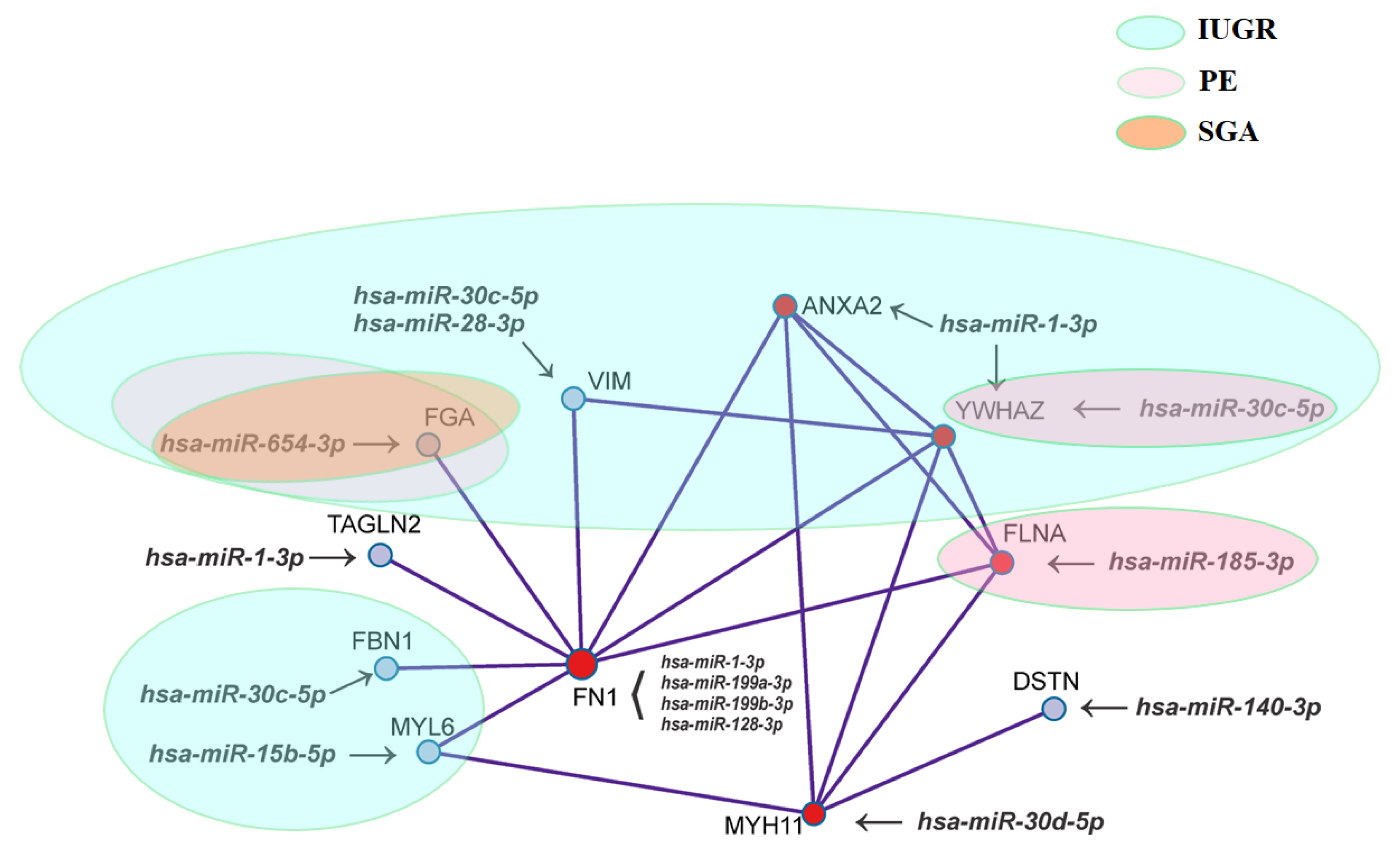
2.4. Bioinformatic Search for miRNA–Target Gene Pairs that Differentiate IUGR from PE
2.5. Assessment of the Expression Level of miRNAs and Their Target Genes in the Placental Bed Samples by Quantitative RT-PCR
2.6. Partial Least Squares Discriminant Analysis (PLS-DA)
2.7. Functional Annotation of miRNA Target Genes
3. Discussion
4. Materials and Methods
4.1. Patient Cohort
4.2. RNA Isolation from the Placenta and Placenta Bed Samples
4.3. miRNA Deep Sequencing
4.4. Reverse Transcription and Quantitative Real-Time PCR
4.5. Proteomic Tissue Analysis (HPLC-MS/MS)
4.6. Statistical Analysis of the Obtained Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Burton, G.J.; Fowden, A.L.; Thornburg, K.L. Placental Origins of Chronic Disease. Physiol. Rev. 2016, 96, 1509–1565. [Google Scholar] [CrossRef] [PubMed]
- Bassily, E.; Bell, C.; Verma, S.; Patel, N.; Patel, A. Significance of Obstetrical History with Future Cardiovascular Disease Risk. Am. J. Med. 2019, 132, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Bronson, S.L.; Bale, T.L. The Placenta as a Mediator of Stress Effects on Neurodevelopmental Reprogramming. Neuropsychopharmacology 2016, 41, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Steegers, E.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-eclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef]
- Walker, J.J. Management of Severe Pre-Eclampsia/Eclampsia. In Best Practice in Labour and Delivery; Arulkumaran, S.S., Ed.; Cambridge University Press: Cambridge, UK, 2016; pp. 301–331. ISBN 9781107472341. [Google Scholar]
- Lausman, A.; McCarthy, F.P.; Walker, M.; Kingdom, J. Screening, diagnosis, and management of intrauterine growth restriction. J. Obstet. Gynaecol. Can. 2012, 34, 17–28. [Google Scholar] [CrossRef]
- Nardozza, L.M.M.; Caetano, A.C.R.; Zamarian, A.C.P.; Mazzola, J.B.; Silva, C.P.; Marçal, V.M.G.; Lobo, T.F.; Peixoto, A.B.; Júnior, E.A. Fetal growth restriction: Current knowledge. Arch. Gynecol. Obstet. 2017, 295, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- Schlembach, D. Fetal Growth Restriction—Diagnostic Work-up, Management and Delivery. Geburtshilfe Frauenheilkd. 2020, 80, 1016–1025. [Google Scholar] [CrossRef]
- Aplin, J.D.; Myers, J.E.; Timms, K.; Westwood, M. Tracking placental development in health and disease. Nat. Rev. Endocrinol. 2020, 16, 479–494. [Google Scholar] [CrossRef]
- Knöfler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.K.J.B.; James, J. Human placenta and trophoblast development: Key molecular mechanisms and model systems. Cell. Mol. Life Sci. 2019, 76, 3479–3496. [Google Scholar] [CrossRef] [Green Version]
- Pijnenborg, R.; Bland, J.; Robertson, W.; Brosens, I. Uteroplacental arterial changes related to interstitial trophoblast migration in early human pregnancy. Placenta 1983, 4, 397–413. [Google Scholar] [CrossRef]
- Huppertz, B. Placental Origins of Preeclampsia. Hypertension 2008, 51, 970–975. [Google Scholar] [CrossRef]
- Ekropp, J.; Salih, S.M.; Ekhatib, H. Expression of microRNAs in bovine and human pre-implantation embryo culture media. Front. Genet. 2014, 5, 91. [Google Scholar] [CrossRef]
- Sánchez-Ribas, I.; Díaz-Gimeno, P.; Quiñonero, A.; Ojeda, M.; Larreategui, Z.; Ballesteros, A.; Domínguez, F. NGS Analysis of Human Embryo Culture Media Reveals miRNAs of Extra Embryonic Origin. Reprod. Sci. 2018, 26, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Abu Khaizaran, Z.; Ayesh, B.M.; Fischer, U.; Abu Khaizaran, S.; Al-Battah, F.; Hammadeh, M.; Keller, A.; Meese, E. MicroRNAs in combined spent culture media and sperm are associated with embryo quality and pregnancy outcome. Fertil. Steril. 2020, 113, 970–980.e2. [Google Scholar] [CrossRef]
- Cuman, C.; Van Sinderen, M.; Gantier, M.P.; Rainczuk, K.; Sorby, K.; Rombauts, L.; Osianlis, T.; Dimitriadis, E. Human Blastocyst Secreted microRNA Regulate Endometrial Epithelial Cell Adhesion. EBioMedicine 2015, 2, 1528–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, C.C.; Calhoun, S.H.; Wieden, H.-J. Non-coding RNAs: What are we missing? Biochem. Cell Biol. 2020, 98, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, A.V.; Chagovets, V.V.; Drapkina, Y.S.; Makarova, N.P.; Kalinina, E.A.; Sukhikh, G.T. Cell-Free, Embryo-Specific sncRNA as a Molecular Biological Bridge between Patient Fertility and IVF Efficiency. Int. J. Mol. Sci. 2019, 20, 2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timofeeva, A.; Drapkina, Y.; Fedorov, I.; Chagovets, V.; Makarova, N.; Shamina, M.; Kalinina, E.; Sukhikh, G. Small Noncoding RNA Signatures for Determining the Developmental Potential of an Embryo at the Morula Stage. Int. J. Mol. Sci. 2020, 21, 9399. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-Q.; Zhang, L. Hu MicroRNAs in Uteroplacental Vascular Dysfunction. Cells 2019, 8, 1344. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timofeeva, A.V.; Gusar, V.A.; Kan, N.E.; Prozorovskaya, K.N.; Karapetyan, A.O.; Bayev, O.R.; Chagovets, V.V.; Kliver, S.F.; Iakovishina, D.Y.; Frankevich, V.E.; et al. Identification of potential early biomarkers of preeclampsia. Placenta 2018, 61, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Paquette, A.G.; Shynlova, O.; Kibschull, M.; Price, N.D.; Lye, S.J. Global Alliance to Prevent Prematurity and Stillbirth Systems Biology of Preterm Birth Team Comparative analysis of gene expression in maternal peripheral blood and monocytes during spontaneous preterm labor. Am. J. Obstet. Gynecol. 2018, 218, 345.e1–345.e30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishnyakova, P.; Elchaninov, A.; Fatkhudinov, T.; Sukhikh, G. Role of the Monocyte–Macrophage System in Normal Pregnancy and Preeclampsia. Int. J. Mol. Sci. 2019, 20, 3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrolia, S.A.; Mazor, M.; Loverro, G.; Klaitman, V.; Erez, O. Placental vascular pathology and increased thrombin generation as mechanisms of disease in obstetrical syndromes. PeerJ 2014, 2, e653. [Google Scholar] [CrossRef] [Green Version]
- Myatt, L. Review: Reactive oxygen and nitrogen species and functional adaptation of the placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.; Woods, A.; Jauniaux, E.; Kingdom, J. Rheological and Physiological Consequences of Conversion of the Maternal Spiral Arteries for Uteroplacental Blood Flow during Human Pregnancy. Placenta 2009, 30, 473–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindström, T.M.; Bennett, P.R. The role of nuclear factor kappa B in human labour. Reproduction 2005, 130, 569–581. [Google Scholar] [CrossRef]
- Johnstone, E.D.; Sibley, C.P.; Lowen, B.; Guilbert, L.J. Epidermal Growth Factor Stimulation of Trophoblast Differentiation Requires MAPK11/14 (p38 MAP Kinase) Activation. Biol. Reprod. 2005, 73, 1282–1288. [Google Scholar] [CrossRef]
- Dikshit, S. Fresh look at the doppler changes in pregnancies with placental-based complications. J. Postgrad. Med. 2011, 57, 138–140. [Google Scholar] [CrossRef]
- Spinillo, A.; Gardella, B.; Bariselli, S.; Alfei, A.; Silini, E.; Bello, B.D. Placental histopathological correlates of umbilical artery Doppler velocimetry in pregnancies complicated by fetal growth restriction. Prenat. Diagn. 2012, 32, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Viscardi, R.M.; Sun, C.-C.J. Placental lesion multiplicity: Risk factor for IUGR and neonatal cranial ultrasound abnormalities. Early Hum. Dev. 2001, 62, 1–10. [Google Scholar] [CrossRef]
- Mone, F.; Thompson, A.; Stewart, M.C.; Ong, S.; Shields, M.D. Fetal umbilical artery Doppler pulsatility index as a predictor of cardiovascular risk factors in children—A long-term follow up study. J. Matern. Neonatal Med. 2014, 27, 1633–1636. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Guevara, O.E.; Warburton, R.R.; Hill, N.S.; Gaestel, M.; Kayyali, U.S. Regulation of vimentin intermediate filaments in endothelial cells by hypoxia. Am. J. Physiol. Physiol. 2010, 299, C363–C373. [Google Scholar] [CrossRef] [Green Version]
- Ivaska, J.; Pallari, H.-M.; Nevo, J.; Eriksson, J.E. Novel functions of vimentin in cell adhesion, migration, and signaling. Exp. Cell Res. 2007, 313, 2050–2062. [Google Scholar] [CrossRef]
- Lui, S.; Duval, C.; Farrokhnia, F.; Girard, S.; Harris, L.K.; Tower, C.L.; Stevens, A.; Jones, R.L. Delineating differential regulatory signatures of the human transcriptome in the choriodecidua and myometrium at term labor. Biol. Reprod. 2018, 98, 422–436. [Google Scholar] [CrossRef]
- Sak, M.; Deveci, E.; Turgut, A.; Sak, S.; Evsen, M.S.; Gul, T.; Kalkanli, S. Placental expression of vimentin, desmin and ultrastructural changes in the villi in patients with HELLP syndrome. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 874–878. [Google Scholar]
- Du, L.; Kuang, L.; He, F.; Tang, W.; Sun, W.; Chen, D. Mesenchymal-to-epithelial transition in the placental tissues of patients with preeclampsia. Hypertens. Res. 2016, 40, 67–72. [Google Scholar] [CrossRef]
- Pieters, M.; Wolberg, A.S. Fibrinogen and fibrin: An illustrated review. Res. Pr. Thromb. Haemost. 2019, 3, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Iwaki, T.; Castellino, F.J. Maternal Fibrinogen is Necessary for Embryonic Development. Curr. Drug Targets 2005, 6, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Snir, A.; Brenner, B.; Paz, B.; Ohel, G.; Lanir, N. The role of fibrin matrices and tissue factor in early-term trophoblast proliferation and spreading. Thromb. Res. 2013, 132, 477–483. [Google Scholar] [CrossRef]
- Iwaki, T.; Sandoval-Cooper, M.J.; Paiva, M.; Kobayashi, T.; Ploplis, V.A.; Castellino, F.J. Fibrinogen Stabilizes Placental-Maternal Attachment During Embryonic Development in the Mouse. Am. J. Pathol. 2002, 160, 1021–1034. [Google Scholar] [CrossRef] [Green Version]
- Sucak, G.T.; Acar, K.; Sucak, A.; Kirazl, S.; Haznedar, R. Increased global fibrinolytic capacity as a clue for activated fibrinolysis in pre-eclampsia. Blood Coagul. Fibrinolysis 2006, 17, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Zhou, Q.; Jiang, F.; Zhou, S.; Lou, Y.; Guo, Q.; Liang, W.; Kong, D.; Ma, D.; Li, X. Changes of plasma and placental tissue factor pathway inhibitor-2 in women with preeclampsia and normal pregnancy. Thromb. Res. 2010, 125, e317–e322. [Google Scholar] [CrossRef]
- Aharon, A.; Lanir, N.; Drugan, A.; Brenner, B. Placental TFPI is decreased in gestational vascular complications and can be restored by maternal enoxaparin treatment. J. Thromb. Haemost. 2005, 3, 2355–2357. [Google Scholar] [CrossRef] [PubMed]
- Erez, O.; Romero, R.; Vaisbuch, E.; Than, N.G.; Kusanovic, J.P.; Mazaki-Tovi, S.; Gotsch, F.; Mittal, P.; Dong, Z.; Chaiworapongsa, T.; et al. Tissue factor activity in women with preeclampsia or SGA: A potential explanation for the excessive thrombin generation in these syndromes. J. Matern. Neonatal Med. 2018, 31, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- Cesarman-Maus, G.; Hajjar, K.A. Molecular mechanisms of fibrinolysis. Br. J. Haematol. 2005, 129, 307–321. [Google Scholar] [CrossRef]
- Hajjar, K. Cellular Receptors in the Regulation of Plasmin Generation. Thromb. Haemost. 1995, 74, 294–301. [Google Scholar] [CrossRef]
- Cesarman, G.; Guevara, C.; Hajjar, K. An endothelial cell receptor for plasminogen/tissue plasminogen activator (t-PA). II. Annexin II-mediated enhancement of t-PA-dependent plasminogen activation. J. Biol. Chem. 1994, 269, 21198–21203. [Google Scholar] [CrossRef]
- Dassah, M.; Deora, A.B.; He, K.; Hajjar, K.A. The endothelial cell annexin A2 system and vascular fibrinolysis. Gen. Physiol. Biophys. 2009, 28, F20–F28. [Google Scholar]
- Ling, Q.; Jacovina, A.T.; Deora, A.; Febbraio, M.; Simantov, R.; Silverstein, R.L.; Hempstead, B.; Mark, W.H.; Hajjar, K.A. Annexin II regulates fibrin homeostasis and neoangiogenesis in vivo. J. Clin. Investig. 2004, 113, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Sano, M.; Matsumoto, M.; Terada, H.; Wang, H.; Kurihara, Y.; Wada, N.; Yamamoto, H.; Kira, Y.; Tachibana, D.; Koyama, M. Increased annexin A2 expression in the placenta of women with acute worsening of preeclampsia. Osaka City Med. J. 2014, 60, 87–93. [Google Scholar]
- Kaczanbourgois, D.; Salles, J.; Hulling, F.; Fauvel, J.; Moisand, A.; Duganeulat, I.; Berrebi, A.; Campistron, G.; Chap, H. Increased content of annexin II (p36) and p11 in human placentabrush-border membrane vesicles during syncytiotrophoblast maturation and differentiation. Placenta 1996, 17, 669–676. [Google Scholar] [CrossRef]
- Hoang, V.M.; Foulk, R.; Clauser, K.; Burlingame, A.; Gibson, B.W.; Fisher, S.J. Functional Proteomics: Examining the Effects of Hypoxia on the Cytotrophoblast Protein Repertoire. Biochemistry 2001, 40, 4077–4086. [Google Scholar] [CrossRef]
- Hu, R.; Jin, H.; Zhou, S.; Yang, P.; Li, X. Proteomic Analysis of Hypoxia-Induced Responses in the Syncytialization of Human Placental Cell Line BeWo. Placenta 2007, 28, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Langbein, M.; Strick, R.; Strissel, P.L.; Vogt, N.; Parsch, H.; Beckmann, M.W.; Schild, R.L. Impaired cytotrophoblast cell–cell fusion is associated with reduced Syncytin and increased apoptosis in patients with placental dysfunction. Mol. Reprod. Dev. 2007, 75, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Longtine, M.S.; Nelson, D.M. Placental Dysfunction and Fetal Programming: The Importance of Placental Size, Shape, Histopathology, and Molecular Composition. Semin. Reprod. Med. 2011, 29, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitken, A. 14-3-3 and its possible role in co-ordinating multiple signalling pathways. Trends Cell Biol. 1996, 6, 341–347. [Google Scholar] [CrossRef]
- Park, J.; Kang, M.; Kim, Y.; Jo, H.; Shin, J.; Choi, W.; Lee, S.; Lee, J.; Paik, W. PKC δ in Preeclamptic Placentas Promotes Bax Dissociation from 14-3-3 ζ through 14-3-3 ζ Phosphorylation. Placenta 2008, 29, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Goossens, K.; Van Soom, A.; Van Poucke, M.; Vandaele, L.; Vandesompele, J.; Van Zeveren, A.; Peelman, L.J. Identification and expression analysis of genes associated with bovine blastocyst formation. BMC Dev. Biol. 2007, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Peñagaricano, F.; Chen, H.; Khatib, H. Novel transcripts and alternatively spliced genes are associated with early development in bovine embryos. Animals 2012, 6, 1199–1205. [Google Scholar] [CrossRef] [Green Version]
- Krendel, M.; Mooseker, M.S. Myosins: Tails (and Heads) of Functional Diversity. Physiology 2005, 20, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Guo, Y.; Shi, H.; Liu, C.-L.; Panganiban, R.A.; Chung, W.; O’Connor, L.J.; Himes, B.E.; Gazal, S.; Hasegawa, K.; et al. Shared genetic and experimental links between obesity-related traits and asthma subtypes in UK Biobank. J. Allergy Clin. Immunol. 2020, 145, 537–549. [Google Scholar] [CrossRef] [Green Version]
- Stossel, T.P.; Condeelis, J.; Cooley, L.; Hartwig, J.H.; Noegel, A.; Schleicher, M.; Shapiro, S.S. Filamins as integrators of cell mechanics and signalling. Nat. Rev. Mol. Cell Biol. 2001, 2, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.-X.; Hartwig, J.H.; Akyürek, L.M. Filamins in cell signaling, transcription and organ development. Trends Cell Biol. 2010, 20, 113–123. [Google Scholar] [CrossRef]
- Bedolla, R.G.; Wang, Y.; Asuncion, A.; Chamie, K.; Siddiqui, S.; Mudryj, M.M.; Prihoda, T.J.; Siddiqui, J.; Chinnaiyan, A.M.; Mehra, R.; et al. Nuclear versus Cytoplasmic Localization of Filamin A in Prostate Cancer: Immunohistochemical Correlation with Metastases. Clin. Cancer Res. 2009, 15, 788–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Fu, Y.; Mao, X.; Jing, Y.; Guo, J.; Ye, Y. Decreased Filamin b expression regulates trophoblastic cells invasion through ERK/MMP-9 pathway in pre-eclampsia. Ginekol. Polska 2019, 90, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Abbas, Y.; Carnicer-Lombarte, A.; Gardner, L.; Thomas, J.; Brosens, J.J.; Moffett, A.; Sharkey, A.M.; Franze, K.; Burton, G.J.; Oyen, M.L. Tissue stiffness at the human maternal–fetal interface. Hum. Reprod. 2019, 34, 1999–2008. [Google Scholar] [CrossRef] [Green Version]
- Halper, J.; Kjaer, M. Basic Components of Connective Tissues and Extracellular Matrix: Elastin, Fibrillin, Fibulins, Fibrinogen, Fibronectin, Laminin, Tenascins and Thrombospondins. Adv. Exp. Med. Biol. 2013, 802, 31–47. [Google Scholar] [CrossRef]
- Korhonen, M.; Virtanen, I. The Distribution of Laminins and Fibronectins Is Modulated During Extravillous Trophoblastic Cell Differentiation and Decidual Cell Response to Invasion in the Human Placenta. J. Histochem. Cytochem. 1997, 45, 569–581. [Google Scholar] [CrossRef]
- Korhonen, M.; Virtanen, I. Immunohistochemical Localization of Laminin and Fibronectin Isoforms in Human Placental Villi. J. Histochem. Cytochem. 2001, 49, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fruscalzo, A.; Londero, A.P.; Orsaria, M.; Marzinotto, S.; Driul, L.; Di Loreto, C.; Mariuzzi, L. Placental fibronectin staining is unaffected in pregnancies complicated by late-onset intrauterine growth restriction and small for gestational age fetuses. Int. J. Gynecol. Obstet. 2019, 148, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Allison, B.J.; Castillo-Melendez, M.; Jenkin, G.; Polglase, G.R.; Miller, S.L. Neonatal Morbidities of Fetal Growth Restriction: Pathophysiology and Impact. Front. Endocrinol. 2019, 10, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jazwiec, P.A.; Li, X.; Matushewski, B.; Richardson, B.S.; Sloboda, D.M. Fetal Growth Restriction Is Associated With Decreased Number of Ovarian Follicles and Impaired Follicle Growth in Young Adult Guinea Pig Offspring. Reprod. Sci. 2019, 26, 1557–1567. [Google Scholar] [CrossRef]
- American College of Obstetricians and Gynecologists ACOG Practice Bulletin. Diagnosis and management of preeclampsia and eclampsia. Number 33, January 2002. American College of Obstetricians and Gynecologists. Int. J. Gynaecol. Obstet. 2002, 77, 67–75. [Google Scholar] [CrossRef]
- Lees, C.C.; Stampalija, T.; Baschat, A.A.; Costa, F.D.S.; Ferrazzi, E.; Figueras, F.; Hecher, K.; Kingdom, J.; Poon, L.C.; Salomon, L.J.; et al. ISUOG Practice Guidelines: Diagnosis and management of small-for-gestational-age fetus and fetal growth restriction. Ultrasound Obstet. Gynecol. 2020, 56, 298–312. [Google Scholar] [CrossRef]
- Burton, G.; Sebire, N.; Myatt, L.; Tannetta, D.; Wang, Y.-L.; Sadovsky, Y.; Staff, A.; Redman, C. Optimising sample collection for placental research. Placenta 2014, 35, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, 25. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 10 March 2021).
- Starodubtseva, N.L.; Brzhozovskiy, A.G.; Bugrova, A.E.; Kononikhin, A.S.; Indeykina, M.I.; Gusakov, K.I.; Chagovets, V.V.; Nazarova, N.M.; Frankevich, V.E.; Sukhikh, G.T.; et al. Label-free cervicovaginal fluid proteome profiling reflects the cervix neoplastic transformation. J. Mass Spectrom. 2019, 54, 693–703. [Google Scholar] [CrossRef]
- Kononikhin, A.S.; Zakharova, N.V.; Sergeeva, V.A.; Indeykina, M.I.; Starodubtseva, N.L.; Bugrova, A.E.; Muminova, K.T.; Khodzhaeva, Z.S.; Popov, I.A.; Shao, W.; et al. Differential Diagnosis of Preeclampsia Based on Urine Peptidome Features Revealed by High Resolution Mass Spectrometry. Diagnostics 2020, 10, 1039. [Google Scholar] [CrossRef]
- R Studio Team. R Studio: Integrated Development for R; R Studio, Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 23 March 2021).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | IUGR < 34 n = 10 | IUGR > 34 n = 13 | PE < 34 n = 12 | PE > 34 n = 7 | SGA > 34 n = 8 | N > 34 n = 7 | p-Value: IUGR < 34 vs. IUGR > 34 PE < 34 vs. PE > 34 IUGR > 34 vs. N > 34 PE > 34 vs. N > 34 SGA > 34 vs. N > 34 |
|---|---|---|---|---|---|---|---|
| Maternal age, years 1 | 30 (28, 35) | 30 (28, 34) | 34 (30, 36) | 35 (35, 37) | 32 (27, 35) | 34 (32, 34) | 0.827 0.468 0.421 0.092 0.598 |
| Diastolic pressure, mm Hg 1 | 70 (69, 74) | 72 (70, 80) | 100 (100, 101) | 100 (100, 100) | 70 (64, 76) | 70 (70, 80) | 0.359 1 0.935 0.003 0.284 |
| Systolic pressure, mm Hg 1 | 112 (110, 120) | 116 (113, 120) | 160 (150, 164) | 163 (155, 173) | 117 (108, 119) | 120 (110, 120) | 0.406 0.416 0.898 0.004 0.398 |
| APGAR 1 min 1 | 6 (5, 7) | 7 (7, 8) | 6.5 (6, 7) | 8 (7, 8) | 8 (8, 8) | 8 (8, 8) | 0.003 0.024 0.098 0.29 0.677 |
| APGAR 5 min 1 | 7 (7, 7) | 8 (8, 9) | 8 (7, 8) | 8 (8, 9) | 8.5 (8, 9) | 8 (8, 9) | <0.001 0.151 0.925 0.762 0.454 |
| BMI, (kg/m2) 1 | 25 (22, 29) | 23 (21, 26) | 26 (24, 30) | 29 (27, 32) | 26 (24, 27) | 23 (22, 26) | 0.304 0.219 0.873 0.054 0.52 |
| IFC, CTG 1 | 0.16 (0.15, 0.61) | 0.42 (0.32, 0.67) | 0.38 (0.29, 0.55) | 0.4 (0.28, 0.57) | 0.56 (0.21, 0.95) | 0.15 (0.09, 0.18) | 0.336 0.832 0.007 <0.001 0.072 |
| PI of Left Uterine Artery 1 | 1.03 (0.83, 1.34) | 0.91 (0.82, 1.03) | 1.06 (0.94, 1.09) | 0.95 (0.76, 1.14) | 0.85 (0.76, 0.9) | 0.91 (0.91, 1) | 0.513 0.523 0.936 0.898 0.092 |
| PI of Right Uterine Artery 1 | 1.05 (0.85, 1.06) | 1.06 (0.98, 1.07) | 1.33 (1.27, 1.39) | 1.3 (0.92, 1.58) | 0.82 (0.78, 0.88) | 0.85 (0.8, 0.92) | 0.851 0.55 0.039 0.095 0.642 |
| Birth weight, g 1 | 1144 (921, 1455) | 2170 (1960, 2228) | 1100 (989, 1421) | 2080 (1735, 2617) | 2300 (2231, 2375) | 2660 (2505, 2700) | <0.001 <0.001 <0.001 0.259 0.056 |
| Placenta mass, g 1 | 225 (216, 251) | 216 (216, 265) | 287 (270, 287) | 287 (281, 324) | 348 (322, 365) | 352 (352, 392) | 0.332 0.48 <0.001 0.122 0.323 |
| PI of umbilical artery 1 | 1.15 (1, 1.31) | 1.05 (0.97, 1.07) | 0.96 (0.94, 1.06) | 0.96 (0.88, 1) | 1.01 (0.92, 1.12) | 0.85 (0.8, 0.85) | 0.123 0.525 <0.001 0.303 0.048 |
| Estimated fetal weight, g 1 | 1324 (1041, 1611) | 1936 (1737, 2114) | 1153 (1033, 1595) | 1985 (1759, 2462) | 2277 (2188, 2349) | 2515 (2400, 2617) | <0.001 <0.001 <0.001 0.097 0.072 |
| Percentile of estimated fetal weight 1 | 2.12 (1.52, 2.25) | 1 (0.2, 2.3) | 15 (3.75, 23.2) | 13 (8.7, 25.39) | 4.3 (3.38, 6.8) | 25 (22.05, 27.75) | 0.418 0.966 <0.001 0.165 <0.001 |
| Gestational age at the time of delivery, weeks 1 | 31 (30, 32) | 37 (37, 37) | 31 (29, 32) | 35 (34, 36) | 37 (36, 38) | 35 (35, 37) | <0.001 <0.001 0.056 0.335 0.063 |
| PI in MCA 1 | 1.87 (1.6, 2.13) | 1.75 (1.51, 1.87) | 1.79 (1.66, 1.79) | 1.68 (1.56, 1.86) | 1.64 (1.49, 1.8) | 1.63 (1.56, 1.63) | 0.437 0.965 0.475 0.335 0.684 |
| Cerebroplacental ratio 1 | 1.43 (0.99, 1.77) | 1.64 (1.54, 1.64) | 1.55 (1.51, 1.55) | 1.61 (1.55, 1.85) | 1.55 (1.41, 1.78) | 2.23 (2.03, 2.23) | 0.434 0.061 0.005 <0.001 0.023 |
| Intraventricular hemorrhage, % 2 | 2 (20%) | 2 (15.4%) | 0 | 0 | 1 (12.5%) | 0 | - |
| Respiratory distress syndrome, % 2 | 10 (100%) | 1 (7.7%) | 12 (100%) | 3 (42.9%) | 0 | 1 (14.3%) | - |
| Proteinuria, % 2 | 0 | 0 | 10 (83.3%) | 4 (57.14%) | 0 | 0 | - |
| miRNA | log2(FC) * | p ** | Gene-Target Name | Protein Name (Protein ID, UniProtKB) | log2(FC) * | p ** |
|---|---|---|---|---|---|---|
| hsa-miR-1-3p | 1.18 | 0.00012 | ANXA2 | Annexin A2 (P07355) | −2.15 | <0.0001 |
| hsa-miR-1-3p | 1.18 | 0.00012 | TAGLN2 | Transgelin-2 (P37802) | −2.088 | <0.0001 |
| hsa-miR-1-3p | 1.18 | 0.00012 | YWHAZ | 14-3-3 protein zeta/delta (P63104) | −1.032 | 0.00193 |
| hsa-miR-1-3p | 1.18 | 0.00012 | FN1 | Fibronectin (P02751) | −6.999 | 0.00504 |
| hsa-miR-199a-3p | 0.72 | 0.00026 | FN1 | Fibronectin (P02751) | −6.999 | 0.00504 |
| hsa-miR-199b-3p | 0.72 | 0.00027 | FN1 | Fibronectin (P02751) | −6.999 | 0.00504 |
| hsa-miR-185-3p | −0.95 | 0.00060 | FLNA | Filamin-A (P21333) | 0.9255 | 0.1388 |
| hsa-miR-128-3p | 0.80 | 0.0024 | FN1 | Fibronectin (P02751) | −6.999 | 0.00504 |
| hsa-miR-140-3p | −0.71 | 0.0025 | DSTN | Destrin (P60981) | 0.878 | 0.00531 |
| hsa-miR-30c-5p | 0.62 | 0.0075 | FBN1 | Fibrillin-1 (P35555) | −6.985 | 0.0689 |
| hsa-miR-30c-5p | 0.62 | 0.0075 | VIM | Vimentin (P08670) | −1.695 | 0.0012 |
| hsa-miR-30c-5p | 0.62 | 0.0075 | YWHAZ | 14-3-3 protein zeta/delta (P63104) | −1.032 | 0.00193 |
| hsa-miR-28-3p | 0.56 | 0.0140 | VIM | Vimentin (P08670) | −1.695 | 0.0012 |
| hsa-miR-30d-5p | −0.59 | 0.02025 | MYH11 | Myosin-11 (P35749) | 1.9163 | <0.0001 |
| hsa-miR-654-3p | 0.57 | 0.0421 | FGA | Fibrinogen alpha chain (P02671) | −7.686 | 0.0002243 |
| hsa-miR-15b-5p | −0.53 | 0.0495 | MYL6 | Myosin light polypeptide 6 (P60660) | 1.5447 | 0.000172 |
| miRNA or mRNA | Groups to Be Compared 1 | p-Value | |
|---|---|---|---|
| IUGR < 34 | IUGR > 34 | ||
| hsa-miR-1-3p | 5.58 (4.71, 6.82) | 7.04 (6.28, 8.9) | 0.0374 |
| hsa-miR-128-3p | 6.9 (5.84, 7.23) | 7.87 (6.98, 8.28) | 0.0268 |
| YWHAZ | 8.76 (8.53, 8.92) | 8.07 (7.49, 8.39) | 0.0268 |
| ANXA2 | 1 (0.57, 1.21) | 0.66 (0.18, 0.77) | 0.0374 |
| FBN1 | 3.73 (3.28, 3.95) | 2.99 (2.58, 3.42) | 0.0439 |
| PE < 34 | PE > 34 | ||
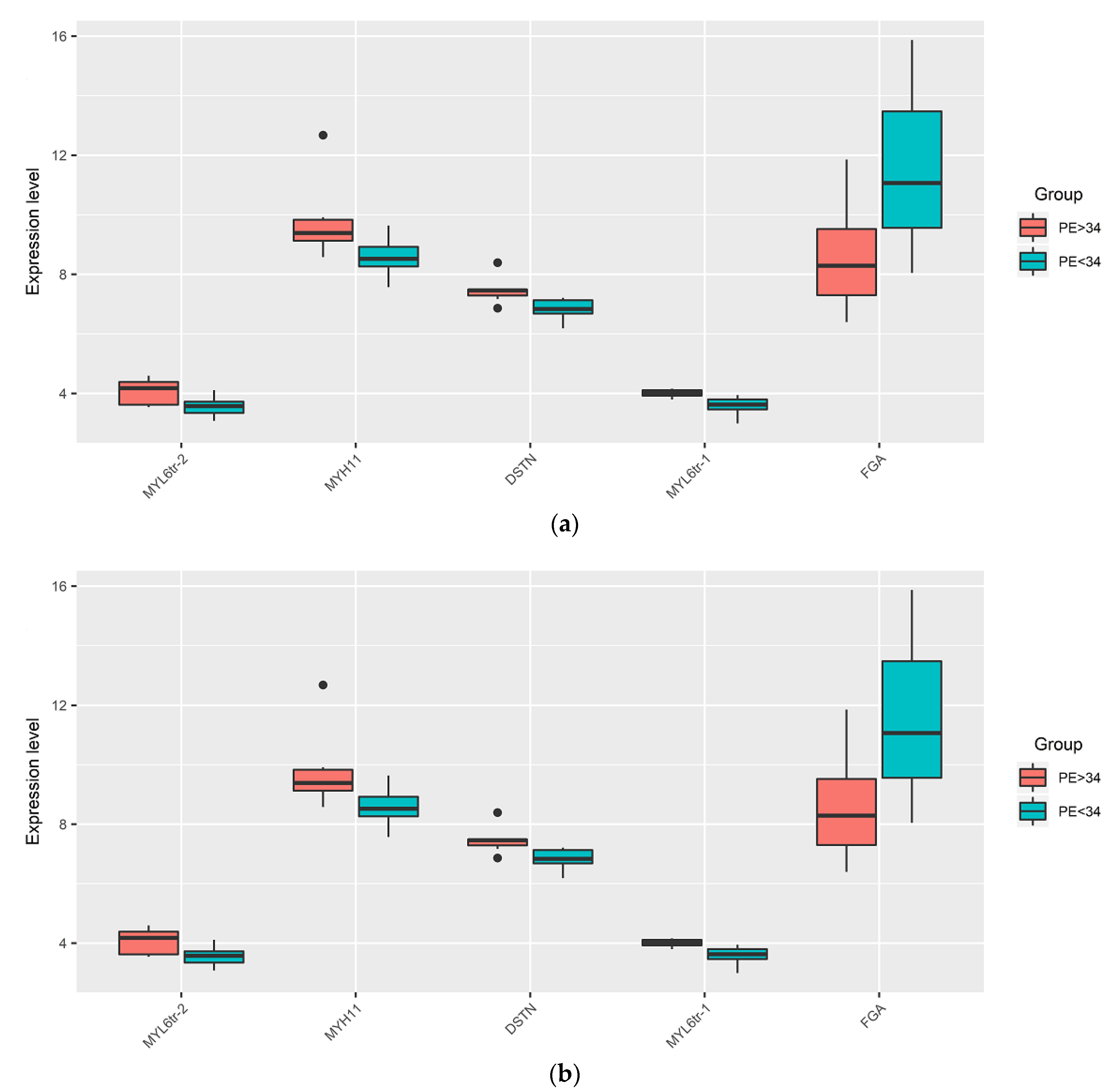
| DSTN | 6.84 (6.68, 7.13) | 7.46 (7.29, 7.49) | 0.0026 |
| FGA | 11.07 (9.56, 13.47) | 8.29 (7.3, 9.52) | 0.0221 |
| MYH11 | 8.52 (8.26, 8.92) | 9.39 (9.13, 9.83) | 0.0098 |
| MYL6 (tr.var.1) | 3.62 (3.46, 3.8) | 4.01 (3.92, 4.11) | 0.0008 |
| MYL6 (tr.var.2) | 3.57 (3.34, 3.72) | 4.17 (3.62, 4.38) | 0.0358 |
| IUGR > 34 | N > 34 | ||
| hsa-miR-1-3p | 7.04 (6.28, 8.9) | 5.37 (3.64, 5.96) | 0.0221 |
| hsa-miR-140-3p | 0.08 (−0.4,0.48) | −0.88 (−1.63, −0.36) | 0.0449 |
| hsa-miR-15b-5p | 2.12 (1.64, 2.46) | 0.77 (0.27, 0.92) | 0.0008 |
| hsa-miR-199a-3p | 5.21 (4.71, 5.79) | 3.89 (2.69, 4.18) | 0.0037 |
| hsa-miR-199b-3p | 5.26 (4.75, 5.7) | 3.58 (2.59, 4.27) | 0.0098 |
| hsa-miR-28-3p | 5.88 (5.53, 6.75) | 4.33 (4.01, 4.8) | 0.0001 |
| hsa-miR-30c-5p | 5.72 (5.56, 6.42) | 4.04 (2.75, 4.6) | 0.0026 |
| hsa-miR-654-3p | 10.85 (9.52, 11.91) | 9.38 (8.49, 9.72) | 0.0449 |
| VIM | 1.06 (0.94, 1.31) | 1.47 (1.39, 1.75) | 0.0221 |
| ANXA2 | 0.66 (0.18, 0.77) | 0.85 (0.81, 1.13) | 0.0130 |
| FGA | 8.59 (5.94, 11.9) | 13.48 (12.06, 17.19) | 0.0098 |
| MYL 6 tr.v.1 | 3.84 (3.5, 4.09) | 4.06 (3.9, 4.33) | 0.0373 |
| MYL 6 tr.v.2 | 3.62 (3.29, 4.1) | 4.03 (3.67, 4.06) | 0.0498 |
| YWHAZ | 8.07 (7.49, 8.39) | 8.7 (8.08, 9.18) | 0.0414 |
| FBN1 | 2.99 (2.58, 3.42) | 3.28 (2.82, 3.78) | 0.0374 |
| SGA > 34 | N > 34 | ||
| FGA | 8.67 (7.53, 12.52) | 13.48 (12.06, 17.19) | 0.0229 |
| MYL 6 tr.v.1 | 3.67 (3.54, 4.11) | 4.06 (3.9, 4.33) | 0.0274 |
| hsa-miR-654-3p | 9.74 (9.06, 12.25) | 9.38 (8.49, 9.72) | 0.0269 |
| hsa-miR-15b-5p | 1.09 (0.95, 1.39) | 0.77 (0.27, 0.92) | 0.0453 |
| hsa-miR-185-3p | 7.87 (7.23, 8.16) | 6.73 (6.36, 7.49) | 0.0302 |
| hsa-miR-140-3p | 0.06 (−0.2, 0.43) | −0.88 (−1.63, −0.36) | 0.0189 |
| PE > 34 | N > 34 | ||
| hsa-miR-30c-5p | 5.82 (4.95, 6.36) | 4.04 (2.75, 4.6) | 0.0379 |
| hsa-miR-654-3p | 9.89 (9.27, 10.99) | 9.38 (8.49, 9.72) | 0.0494 |
| hsa-miR-185-3p | 7.88 (7.45, 8.2) | 6.73 (6.36, 7.49) | 0.0433 |
| FGA | 8.29 (7.3, 9.52) | 13.48 (12.06, 17.19) | 0.0023 |
| FLNA | 7.85 (7.27, 7.87) | 8.13 (7.63, 8.29) | 0.0498 |
| YWHAZ | 8.02 (7.76, 8.43) | 8.7 (8.08, 9.18) | 0.0493 |
| MYH11 | 9.39 (9.13, 9.83) | 8.32 (7.77, 8.96) | 0.0281 |
| miRNA/mRNA Pair | Groups to Be Compared 1 | p-Value | |
|---|---|---|---|
| IUGR > 34 | N > 34 | ||
| miR-28-3p/VIM | −4.74 (−5.79, −4.22) | −2.49 (−3.48, −2.43) | 0.001 |
| miR-654-3p/FGA | −0.81 (−3.93, 0.83) | 4.43 (2.46, 9.85) | 0.006 |
| miR-15b-5p/MYL6 tr.v.2 | 2.13 (1.02, 2.5) | 3.32 (2.7, 3.72) | 0.006 |
| miR-15b-5p/MYL6 tr.v.1 | 1.57 (1.53, 2.46) | 3.29 (2.87, 3.9) | 0.0007 |
| miR-1-3p/ANXA2 | −5.78 (−8.25, −5.23) | −4.13 (−5.04, −2.61) | 0.037 |
| miR-1-3p/YWHAZ | 1.09 (−0.55, 2.43) | 4.28 (2.39, 5.73) | 0.03 |
| miR-30c-5p/FBN1 | −2.73 (−3.7, −2.55) | 0.24 (−1.53, 0.79) | 0.02 |
| miR-30c-5p/YWHAZ | 2 (0.89, 2.63) | 5.81 (3.84, 6.43) | 0.006 |
| miR-30c-5p/VIM | −4.61 (−5.18, −4.32) | −2.37 (−3.2, −1.18) | 0.006 |
| SGA > 34 | N > 34 | ||
| miR-654-3p/FGA | −1.46 (−2.5, 1.18) | 4.43 (2.46, 9.85) | 0.006 |
| PE > 34 | N > 34 | ||
| miR-654-3p/FGA | −1.84 (−2.88, 0.1) | 4.43 (2.46, 9.85) | 0.001 |
| miR-185-3p/FLNA | −0.49 (−0.66, 0.13) | 1.4 (0.53, 1.93) | 0.0026 |
| miR-30c-5p/YWHAZ | 2.82 (1.55, 3.39) | 5.81 (3.84, 6.43) | 0.0026 |
| IUGR > 34 | SGA > 34 | ||
| miR-30c-5p/VIM | −4.61 (−5.18, −4.32) | −3.16 (−3.78, −2.87) | 0.016 |
| miR-15b-5p/MYL6 tr.v.1 | 1.57 (1.53, 2.46) | 2.58 (2.1, 3.06) | 0.01 |
| miR-30c-5p/FBN1 | −2.73 (−3.7, −2.55) | −1.62 (−1.98, −0.98) | 0.02 |
| PE > 34 | IUGR > 34 | ||
| miR-15b-5p/MYL6 tr.v.2 | 2.8 (2.74, 3.29) | 2.13 (1.02, 2.5) | 0.001 |
| miR-15b-5p/MYL6 tr.v.1 | 3.22 (2.31, 3.29) | 1.57 (1.53, 2.46) | 0.0046 |
| ID | Category | Description | Hits 1 | Lg(P) | Lg(Q) |
|---|---|---|---|---|---|
| 1 | GO Biological Processes | supramolecular fiber organization | ANXA2|FLNA|MYH11|VIM|DSTN | −5.35 | −2.35 |
| 2 | Reactome Gene Sets | Signaling by Interleukins | ANXA2|FN1|VIM|YWHAZ | −4.66 | −1.86 |
| 3 | Reactome Gene Sets | RHO GTPases activate PAKs | FLNA|MYH11|MYL6 | −7.04 | −3.42 |
| 4 | Reactome Gene Sets | Platelet activation, signaling and aggregation | FGA|FLNA|FN1|YWHAZ|TAGLN2 | −7.51 | −3.42 |
| 5 | GO Biological Processes | blood vessel development | ANXA2|FN1|YWHAZ | −2.50 | 0.00 |
| 6 | GO Biological Processes | blood vessel morphogenesis | ANXA2|FN1|YWHAZ | −2.64 | −0.07 |
| 7 | GO Biological Processes | protein localization to membrane | ANXA2|FLNA|YWHAZ | −2.74 | −0.15 |
| 8 | GO Biological Processes | angiogenesis | ANXA2|FN1|YWHAZ | −2.82 | −0.22 |
| 9 | GO Biological Processes | positive regulation of organelle organization | ANXA2|FLNA|YWHAZ|DSTN | −4.10 | −1.36 |
| 10 | GO Biological Processes | actin cytoskeleton organization | FLNA|MYH11|DSTN | −2.64 | −0.07 |
| 11 | GO Biological Processes | actin filament-based movement | FLNA|MYL6|VIM | −4.64 | −1.85 |
| 12 | GO Biological Processes | muscle system process | FLNA|MYH11|MYL6|VIM | −4.66 | −1.86 |
| 13 | Reactome Gene Sets | Signaling by Rho GTPases | FLNA|MYH11|MYL6|YWHAZ | −4.68 | −1.86 |
| 14 | GO Biological Processes | actin-mediated cell contraction | FLNA|MYL6|VIM | −4.93 | −2.06 |
| 15 | GO Biological Processes | muscle contraction | FLNA|MYH11|MYL6|VIM | −5.09 | −2.17 |
| 16 | Reactome Gene Sets | RHO GTPases activate PKNs | MYH11|MYL6|YWHAZ | −5.21 | −2.26 |
| 17 | Reactome Gene Sets | RHO GTPase Effectors | FLNA|MYH11|MYL6|YWHAZ | −5.25 | −2.28 |
| 18 | Reactome Gene Sets | Muscle contraction | ANXA2|MYH11|MYL6|VIM | −6.04 | −2.89 |
| 19 | Reactome Gene Sets | Smooth Muscle Contraction | ANXA2|MYH11|MYL6 | −6.35 | −3.09 |
| 20 | GO Biological Processes | platelet activation | FGA|FLNA|FN1|YWHAZ | −6.49 | −3.19 |
| 21 | GO Biological Processes | extracellular matrix organization | ANXA2|FBN1|FGA|FN1|MYH11 | −6.62 | −3.27 |
| 22 | Reactome Gene Sets | Response to elevated platelet cytosolic Ca2+ | FGA|FLNA|FN1|TAGLN2 | −6.79 | −3.40 |
| 23 | Reactome Gene Sets | Platelet degranulation | FGA|FLNA|FN1|TAGLN2 | −6.86 | −3.42 |
| 24 | GO Biological Processes | platelet degranulation | FGA|FLNA|FN1|TAGLN2 | −6.86 | −3.42 |
| 25 | GO Biological Processes | coagulation | ANXA2|FGA|FLNA|FN1|YWHAZ | −6.89 | −3.42 |
| 26 | GO Biological Processes | hemostasis | ANXA2|FGA|FLNA|FN1|YWHAZ | −6.90 | −3.42 |
| 27 | GO Biological Processes | blood coagulation | ANXA2|FGA|FLNA|FN1|YWHAZ | −6.93 | −3.42 |
| 28 | Reactome Gene Sets | Hemostasis | ANXA2|FGA|FLNA|FN1|YWHAZ|TAGLN2 | −7.31 | −3.42 |
| miRNA or mRNA | miRNA or mRNA Accession Number (miRbase, Unigene) | Nucleotide Sequence of PCR Primer, 5’-3’ | PCR Primers Annealing Temperature, °C |
|---|---|---|---|
| hsa-miR-1-3p | MIMAT0000416 | TGGAATGTAAAGAAGTATGTAT | 52.7 |
| hsa-miR-199a-3p | MIMAT0000232 | ACAGTAGTCTGCACATTGGTTA | 57.6 |
| hsa-miR-199b-3p | MIMAT0004563 | ACAGTAGTCTGCACATTGGTTA | 57.6 |
| hsa-miR-185-3p | MIMAT0004611 | AGGGGCTGGCTTTCCTCTGGTC | 48.9 |
| hsa-miR-140-3p | MIMAT0004597 | TACCACAGGGTAGAACCACGG | 48.9 |
| hsa-miR-30c-5p | MIMAT0000244 | TGTAAACATCCTACACTCTCAGC | 57.6 |
| hsa-miR-654-3p | MIMAT0004814 | TATGTCTGCTGACCATCACCTT | 52.7 |
| hsa-miR-15b-5p | MIMAT0000417 | TAGCAGCACATCATGGTTTACA | 57.6 |
| hsa-miR-128-3p | MIMAT0000424 | TCACAGTGAACCGGTCTCTTT | 59 |
| hsa-miR-28-3p | MIMAT0004502 | CACTAGATTGTGAGCTCCTGGA | 54 |
| hsa-miR-30d-5p | MIMAT0000245 | TGTAAACATCCCCGACTGGAAG | 54 |
| SNORD68 | NR_002450 | Hs_SNORD68_11 miScript Primer Assay, Qiagen | 55 |
| ANXA2 F | NM_004039.3 | CAGCATTTGGGGACGCTCTCA | 48.9 |
| ANXA2 R | AATGGTGACCTCATCCACACC | ||
| TAGLN2 F | NM_003564.3 | CCCTCACTGTGCTGCTCTTT | 52.7 |
| TAGLN2 R | GCCATCCTTGAGCCAGTTCT | ||
| YWHAZ F | NM_003406.4 | ACGACGTCCCTCAAACCTTG | 52.7 |
| YWHAZ R | TGACCTACGGGCTCCTACAA | ||
| FN1 F | XM_017003692.1 | TGGTGCCATGACAATGGTGT | 48.9 |
| FN1 R | CGGGAATCTTCTCTGTCAGCC | ||
| FLNA F | NM_001456.4 | AGTGTCAATCGGAGGTCACG | 61.6 |
| FLNA R | CTGGTCACATCCAGCCCATT | ||
| DSTN F | NM_001011546.2 | AGTGTTGGGCCAGGCTTTAG | 57.6 |
| DSTN R | TGCATCCTTGGAGCTTGCAT | ||
| FBN1 F | NM_000138.5 | ACATCTCCGCGTGTATCGAC | 57.6 |
| FBN1 R | CACAGGTCCCACTTAGGCAG | ||
| VIM F | NM_003380.5 | GGACCAGCTAACCAACGACA | 46.2 |
| VIM R | AAGGTCAAGACGTGCCAGAG | ||
| MYH11 F | NM_002474.3 | AGCGGTACTTCTCAGGGCTA | 52.7 |
| MYH11 R | AGATGGGCCTTGCGTGATAC | ||
| FGA F | NM_000508.5 | GCAAAGATTCAGACTGGCCC | 61.6 |
| FGA R | CTACTGCATGACCCTCGACA | ||
| MYL6 tr.v.1 F | NM_021019.5 | ACTTCACCGAAGACCAGACC | 57.6 |
| MYL6 tr.v.1 R | TATGCCTCACAAACGCTTCATAG | ||
| MYL6 tr.v.2 F | NM_079423.4 | ACTTCACCGAAGACCAGACC | 52.7 |
| MYL6 tr.v.2 R | GCGGACGAGCTCTTCATAGT | ||
| GAPDH F | NM_002046 | ACCACAGTCCATGCCATCAC | 60 |
| GAPDH R | TCCACCACCCTGTTGCTGTA | ||
| ACTB F | NM_001101 | GGACTTCGAGCAAGAGATGG | 60 |
| ACTB R | AGCACTGTGTTGGCGTACAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timofeeva, A.V.; Fedorov, I.S.; Brzhozovskiy, A.G.; Bugrova, A.E.; Chagovets, V.V.; Volochaeva, M.V.; Starodubtseva, N.L.; Frankevich, V.E.; Nikolaev, E.N.; Shmakov, R.G.; et al. miRNAs and Their Gene Targets—A Clue to Differentiate Pregnancies with Small for Gestational Age Newborns, Intrauterine Growth Restriction, and Preeclampsia. Diagnostics 2021, 11, 729. https://doi.org/10.3390/diagnostics11040729
Timofeeva AV, Fedorov IS, Brzhozovskiy AG, Bugrova AE, Chagovets VV, Volochaeva MV, Starodubtseva NL, Frankevich VE, Nikolaev EN, Shmakov RG, et al. miRNAs and Their Gene Targets—A Clue to Differentiate Pregnancies with Small for Gestational Age Newborns, Intrauterine Growth Restriction, and Preeclampsia. Diagnostics. 2021; 11(4):729. https://doi.org/10.3390/diagnostics11040729
Chicago/Turabian StyleTimofeeva, Angelika V., Ivan S. Fedorov, Alexander G. Brzhozovskiy, Anna E. Bugrova, Vitaliy V. Chagovets, Maria V. Volochaeva, Natalia L. Starodubtseva, Vladimir E. Frankevich, Evgeny N. Nikolaev, Roman G. Shmakov, and et al. 2021. "miRNAs and Their Gene Targets—A Clue to Differentiate Pregnancies with Small for Gestational Age Newborns, Intrauterine Growth Restriction, and Preeclampsia" Diagnostics 11, no. 4: 729. https://doi.org/10.3390/diagnostics11040729