
Imbalance of Mg Homeostasis as a Potential Biomarker in Colon Cancer

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Sample Preparation
2.3. Synchrotron Analysis and Microscope Setup
2.4. Acquisition Protocol and Post-Processing
2.5. Literature and Database Analysis of Mg Transporters
2.6. Statistical Analysis
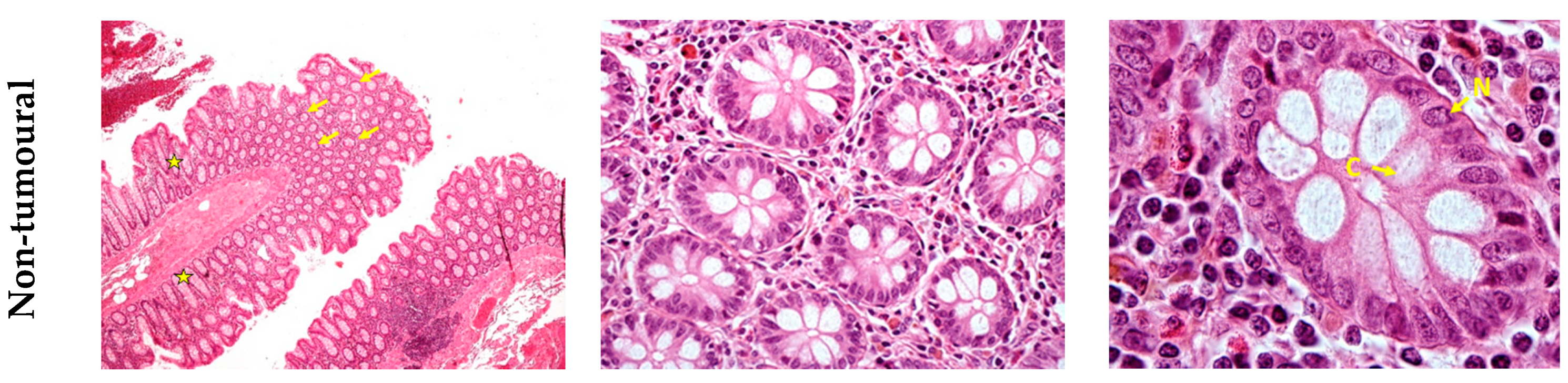
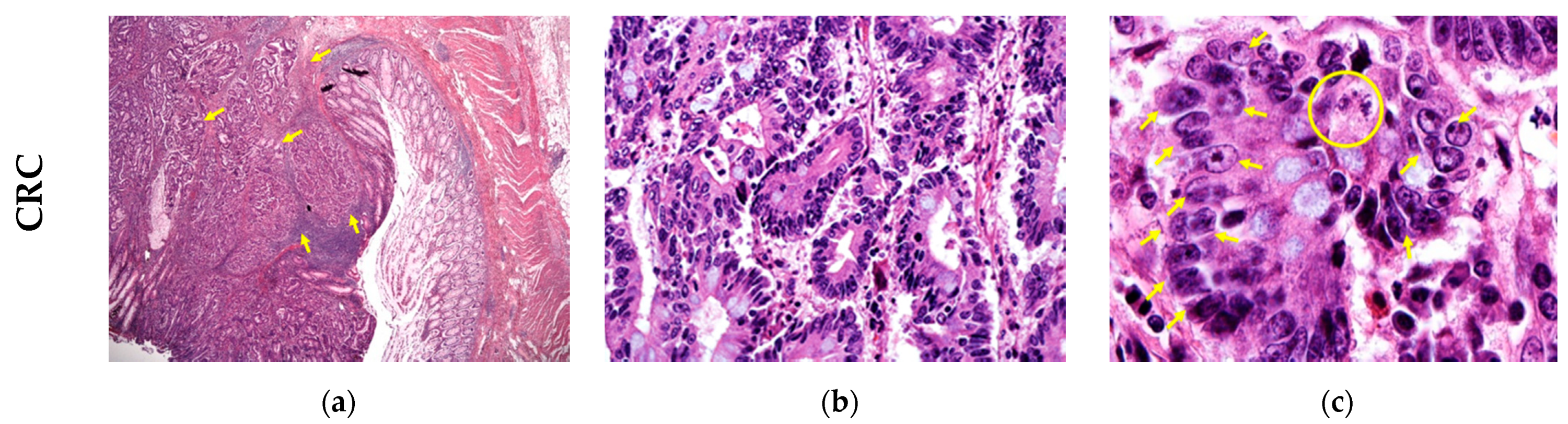
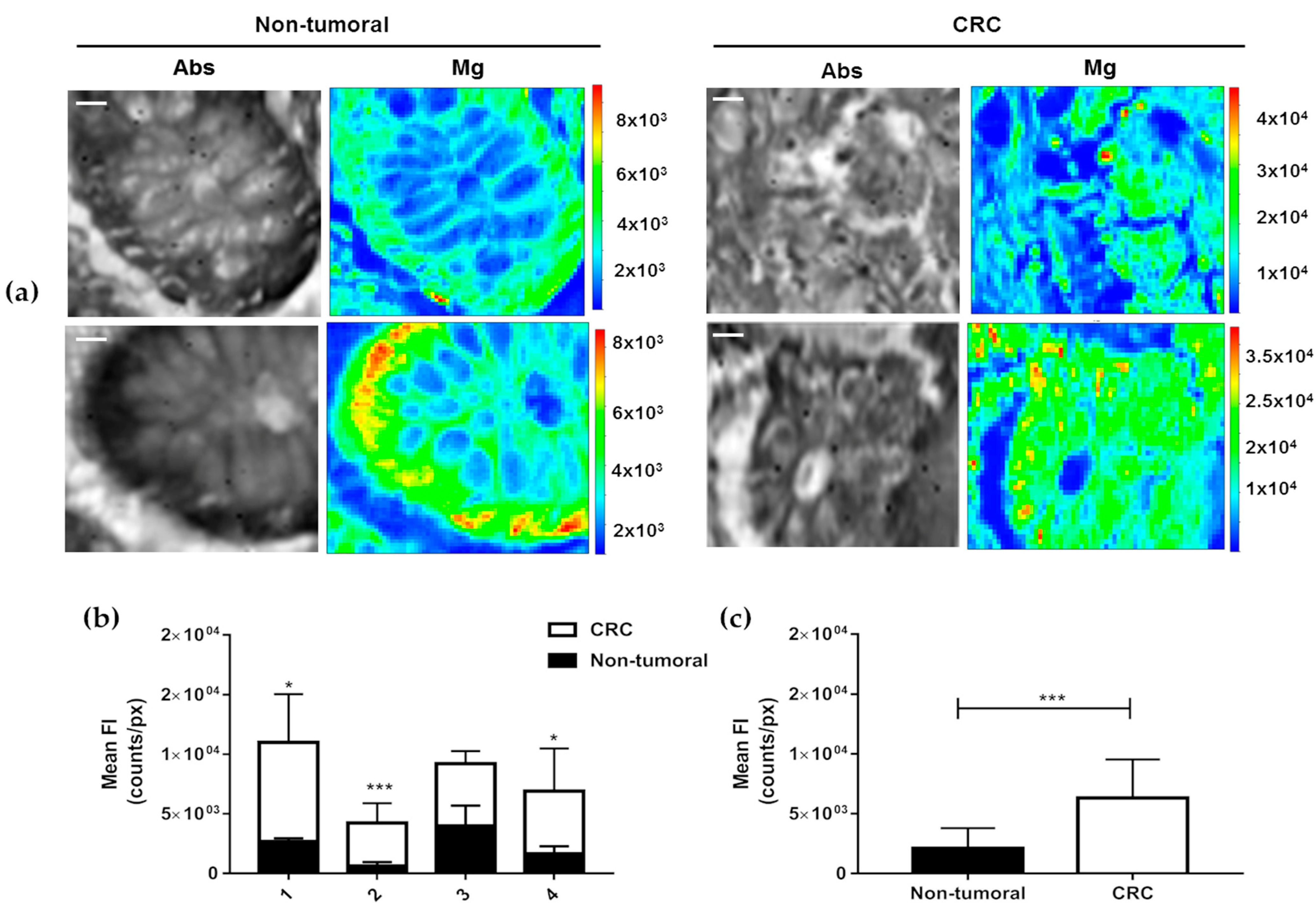
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jung, G.; Hernández-Illán, E.; Moreira, L.; Balaguer, F.; Goel, A. Epigenetics of colorectal cancer: Biomarker and therapeutic potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [CrossRef]
- Binefa, G.; Rodríguez-Moranta, F.; Teule, A.; Medina-Hayas, M. Colorectal cancer: From prevention to personalized medicine. World J. Gastroenterol. 2014, 20, 6786–6808. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, H.; Ushijima, T. Accumulation of genetic and epigenetic alterations in normal cells and cancer risk. NPJ Precis. Oncol. 2019, 3, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardhan, K.; Liu, K. Epigenetics and colorectal cancer pathogenesis. Cancers 2013, 5, 676–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Wark, P.A.; Lau, R.; Norat, T.; Kampman, E. Magnesium intake and colorectal tumor risk: A case-control study and meta-analysis. Am. J. Clin. Nutr. 2012, 96, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Pang, Z.; Liu, Q. Magnesium intake and risk of colorectal cancer: A meta-analysis of prospective studies. Eur. J. Clin. Nutr. 2012, 66, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Sun, J.; Yu, J.; Wang, C.; Su, J. Dietary intakes of calcium, iron, magnesium, and potassium elements and the risk of colorectal cancer: A meta-analysis. Biol. Trace Elem. Res. 2019, 189, 325–335. [Google Scholar] [CrossRef]
- Kuno, T.; Hatano, Y.; Tomita, H.; Hara, A.; Hirose, Y.; Hirata, A.; Mori, H.; Terasaki, M.; Masuda, S.; Tanaka, T. Organomagnesium suppresses inflammation-associated colon carcinogenesis in male Crj: CD-1 mice. Carcinogenesis 2013, 34, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaszczyk, U.; Duda-Chodak, A. Magnesium: Its role in nutrition and carcinogenesis. Rocz. Panstw. Zakl. Hig. 2013, 64, 165–171. [Google Scholar] [PubMed]
- Sartori, S.; Nielsen, I.; Tassinari, D.; Mazzotta, D.; Vecchiatti, G.; Sero, A.; Abbasciano, V. Serum and erythrocyte magnesium concentrations in solid tumours: Relationship with stage of malignancy. Magnes. Res. 1992, 5, 189–192. [Google Scholar] [PubMed]
- Polter, E.J.; Onyeaghala, G. Prospective Association of Serum and Dietary Magnesium with Colorectal Cancer Incidence. Cancer Epidemiol. Prev. Biomark. 2019, 28, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Vincenzi, B.; Galluzzo, S.; Santini, D.; Rocci, L.; Loupakis, F.; Correale, P.; Addeo, R.; Zoccoli, A.; Napolitano, A.; Graziano, F.; et al. Early magnesium modifications as a surrogate marker of efficacy of cetuximab-based anticancer treatment in KRAS wild-type advanced colorectal cancer patients. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2011, 22, 1141–1146. [Google Scholar] [CrossRef]
- Vincenzi, B.; Santini, D.; Galluzzo, S.; Russo, A.; Fulfaro, F.; Silletta, M.; Battistoni, F.; Rocci, L.; Zobel, B.B.; Adamo, V.; et al. Early magnesium reduction in advanced colorectal cancer patients treated with cetuximab plus irinotecan as predictive factor of efficacy and outcome. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 4219–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, C.; Heinemann, V.; Heinrich, K.; Haas, M.; Holch, J.W.; Fraccaroli, A.; Held, S.; von Einem, J.C.; Modest, D.P.; Fischer von Weikersthal, L.; et al. Predictive and prognostic value of magnesium serum level in FOLFIRI plus cetuximab or bevacizumab treated patients with stage IV colorectal cancer: Results from the FIRE-3 (AIO KRK-0306) study. Anti Cancer Drugs 2020, 31, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Tejpar, S.; Piessevaux, H.; Claes, K.; Piront, P.; Hoenderop, J.G.; Verslype, C.; van Cutsem, E. Magnesium wasting associated with epidermal-growth-factor receptor-targeting antibodies in colorectal cancer: A prospective study. Lancet Oncol. 2007, 8, 387–394. [Google Scholar] [CrossRef]
- Trapani, V.; Wolf, F.I. Dysregulation of Mg2+ homeostasis contributes to acquisition of cancer hallmarks. Cell Calcium 2019, 83, 102078. [Google Scholar] [CrossRef]
- Wolf, F.I.; Trapani, V.; Cittadini, A. Magnesium and the control of cell proliferation: Looking for a needle in a haystack. Magnes. Res. 2008, 21, 83–91. [Google Scholar] [PubMed]
- Li, F.-Y.; Chaigne-Delalande, B.; Kanellopoulou, C.; Davis, J.C.; Matthews, H.F.; Douek, D.C.; Cohen, J.I.; Uzel, G.; Su, H.C.; Lenardo, M.J. Second messenger role for Mg2+ revealed by human T-cell immunodeficiency. Nature 2011, 475, 471–476. [Google Scholar] [CrossRef]
- Stangherlin, A.; O’Neill, J.S. Signal transduction: Magnesium manifests as a second messenger. Curr. Biol. 2018, 28, R1403–R1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapani, V.; Arduini, D.; Cittadini, A.; Wolf, F.I. From magnesium to magnesium transporters in cancer: TRPM7, a novel signature in tumour development. Magnes. Res. 2013, 26, 149–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.J.; Cormier, R.T.; Scott, P.M. Role of ion channels in gastrointestinal cancer. World J. Gastroenterol. 2019, 25, 5732–5772. [Google Scholar] [CrossRef]
- Liu, L.; Wu, N.; Wang, Y.; Zhang, X.; Xia, B.; Tang, J.; Cai, J.; Zhao, Z.; Liao, Q.; Wang, J. TRPM7 promotes the epithelial-mesenchymal transition in ovarian cancer through the calcium-related PI3K/AKT oncogenic signaling. J. Exp. Clin. Cancer Res. CR 2019, 38, 106. [Google Scholar] [CrossRef] [PubMed]
- Yee, N.S. Role of TRPM7 in Cancer: Potential as Molecular Biomarker and Therapeutic Target. Pharmaceuticals 2017, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Stokłosa, P.; Borgström, A.; Kappel, S.; Peinelt, C. TRP channels in digestive tract cancers. Int. J. Mol. Sci. 2020, 21, 1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farruggia, G.; Castiglioni, S.; Sargenti, A.; Marraccini, C.; Cazzaniga, A.; Merolle, L.; Iotti, S.; Cappadone, C.; Maier, J.A.M. Effects of supplementation with different Mg salts in cells: Is there a clue? Magnes. Res. 2014, 27, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Castiglioni, S.; Cazzaniga, A.; Trapani, V.; Cappadone, C.; Farruggia, G.; Merolle, L.; Wolf, F.I.; Iotti, S.; Maier, J.A.M. Magnesium homeostasis in colon carcinoma LoVo cells sensitive or resistant to doxorubicin. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef]
- Merolle, L.; Sponder, G.; Sargenti, A.; Mastrototaro, L.; Cappadone, C.; Farruggia, G.; Procopio, A.; Malucelli, E.; Parisse, P.; Gianoncelli, A.; et al. Overexpression of the mitochondrial Mg channel MRS2 increases total cellular Mg concentration and influences sensitivity to apoptosis. Metallomics 2018, 10, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Fatholahi, M.; LaNoue, K.; Romani, A.; Scarpa, A. Relationship between total and free cellular Mg(2+) during metabolic stimulation of rat cardiac myocytes and perfused hearts. Arch. Biochem. Biophys. 2000, 374, 395–401. [Google Scholar] [CrossRef]
- Merolle, L.; Ragazzi, M.; Gianoncelli, A.; Altissimo, M.; Ciarrocchi, A.; Bedolla, D.E.; Marraccini, C.; Baricchi, R.; Pertinhez, T.A. Mapping fundamental life elements in papillary thyroid carcinoma tissue. J. Instrum. 2018, 13, C05018. [Google Scholar] [CrossRef]
- Nagtegaal, I.D.; Odze, R.D.; Klimstra, D.; Paradis, V.; Rugge, M.; Schirmacher, P.; Washington, K.M.; Carneiro, F.; Cree, I.A. The 2019 WHO classification of tumours of the digestive system. Histopathology 2020, 76, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gianoncelli, A.; Kourousias, G.; Merolle, L.; Altissimo, M.; Bianco, A. Current status of the TwinMic beamline at Elettra: A soft X-ray transmission and emission microscopy station. J. Synchrotron Radiat. 2016, 23, 1526–1537. [Google Scholar] [CrossRef] [PubMed]
- Gianoncelli, A.; Kourousias, G.; Stolfa, A.; Kaulich, B. Recent developments at the TwinMic beamline at ELETTRA: An 8 SDD detector setup for low energy X-ray Fluorescence. J. Phys. Conf. Ser. 2013, 425, 182001. [Google Scholar] [CrossRef]
- Solé, V.; Papillon, E.; Cotte, M.; Walter, P.; Susini, J. A multiplatform code for the analysis of energy-dispersive X-ray fluorescence spectra. Spectrochim. Acta B At. Spectrosc. 2007, 62, 63–68. [Google Scholar] [CrossRef]
- Sabates-Bellver, J.; Van der Flier, L.G.; de Palo, M.; Cattaneo, E.; Maake, C.; Rehrauer, H.; Laczko, E.; Kurowski, M.A.; Bujnicki, J.M.; Menigatti, M.; et al. Transcriptome profile of human colorectal adenomas. Mol. Cancer Res. MCR 2007, 5, 1263–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khamas, A.; Ishikawa, T.; Shimokawa, K.; Mogushi, K.; Iida, S.; Ishiguro, M.; Mizushima, H.; Tanaka, H.; Uetake, H.; Sugihara, K. Screening for epigenetically masked genes in colorectal cancer Using 5-Aza-2′-deoxycytidine, microarray and gene expression profile. Cancer Genom. Proteom. 2012, 9, 67–75. [Google Scholar]
- Hong, Y.; Ho, K.S.; Eu, K.W.; Cheah, P.Y. A susceptibility gene set for early onset colorectal cancer that integrates diverse signaling pathways: Implication for tumorigenesis. Clin. Cancer Res. 2007, 13, 1107–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auwercx, J.; Rybarczyk, P.; Kischel, P.; Dhennin-Duthille, I.; Chatelain, D.; Sevestre, H.; van Seuningen, I.; Ouadid-Ahidouch, H.; Jonckheere, N.; Gautier, M. Mg2+ Transporters in Digestive Cancers. Nutrients 2021, 13, 210. [Google Scholar] [CrossRef]
- Schlingmann, K.P.; Weber, S.; Peters, M.; Nejsum, L.N.; Vitzthum, H.; Klingel, K.; Kratz, M.; Haddad, E.; Ristoff, E.; Dinour, D.; et al. Hypomagnesemia with secondary hypocalcemia is caused by mutations in TRPM6, a new member of the TRPM gene family. Nat. Genet. 2002, 31, 166–170. [Google Scholar] [CrossRef]
- Pugliese, D.; Armuzzi, A.; Castri, F.; Benvenuto, R.; Mangoni, A.; Guidi, L.; Gasbarrini, A.; Rapaccini, G.L.; Wolf, F.I.; Trapani, V. TRPM7 is overexpressed in human IBD-related and sporadic colorectal cancer and correlates with tumor grade. Dig. Liver Dis. 2020, 52, 1188–1194. [Google Scholar] [CrossRef]
- Su, F.; Wang, B.-F.; Zhang, T.; Hou, X.-M.; Feng, M.-H. TRPM7 deficiency suppresses cell proliferation, migration, and invasion in human colorectal cancer via regulation of epithelial-mesenchymal transition. Cancer Biomark. 2019, 26, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Zhao, R.; Bai, B.; Wu, Y.; Xu, Y.; Lu, S.; Fang, Y.; Wang, Z.; Maswikiti, E.P.; Zhou, X.; et al. Identification of key tumorigenesis-related genes and their microRNAs in colon cancer. Oncol. Rep. 2018, 40, 3551–3560. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Yang, Q.; Xie, L.; Qiu, Z.; Huang, Y.; Lin, Y.; Tu, L.; Cui, C. Overexpression of MAGT1 is associated with aggressiveness and poor prognosis of colorectal cancer. Oncol. Lett. 2019, 18, 3857–3862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funato, Y.; Yamazaki, D.; Mizukami, S.; Du, L.; Kikuchi, K.; Miki, H. Membrane protein CNNM4–dependent Mg2+ efflux suppresses tumor progression. J. Clin. Investig. 2014, 124, 5398–5410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, F.I.; Cittadini, A.R.; Maier, J.A. Magnesium and tumors: Ally or foe? Cancer Treat. Rev. 2009, 35, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Wolf, F.I.; Trapani, V. Magnesium and its transporters in cancer: A novel paradigm in tumour development. Clin. Sci. 2012, 123, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Castiglioni, S.; Maier, J.A. Magnesium and cancer: A dangerous liason. Magnes. Res. 2011, 24, S92–S100. [Google Scholar] [CrossRef] [PubMed]
- Sohrabi, M.; Gholami, A.; Azar, M.H.; Yaghoobi, M.; Shahi, M.M.; Shirmardi, S.; Nikkhah, M.; Kohi, Z.; Salehpour, D.; Khoonsari, M.R.; et al. Trace Element and Heavy Metal Levels in Colorectal Cancer: Comparison Between Cancerous and Non-cancerous Tissues. Biol. Trace Elem. Res. 2018, 183, 1–8. [Google Scholar] [CrossRef]
- Witkowski, M.; Hubert, J.; Mazur, A. Methods of assessment of magnesium status in humans: A systematic review. Magnes. Res. 2011, 24, 163–180. [Google Scholar] [CrossRef] [Green Version]
- Cappadone, C.; Merolle, L.; Marraccini, C.; Farruggia, G.; Sargenti, A.; Locatelli, A.; Morigi, R.; Iotti, S. Intracellular magnesium content decreases during mitochondria-mediated apoptosis induced by a new indole-derivative in human colon cancer cells. Magnes. Res. 2012, 25, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Kubota, T.; Shindo, Y.; Tokuno, K.; Komatsu, H.; Ogawa, H.; Kudo, S.; Kitamura, Y.; Suzuki, K.; Oka, K. Mitochondria are intracellular magnesium stores: Investigation by simultaneous fluorescent imagings in PC12 cells. Biochim. Biophys. Acta 2005, 1744, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Mitri, E.; Millucci, L.; Merolle, L.; Bernardini, G.; Vaccari, L.; Gianoncelli, A.; Santucci, A. A new light on Alkaptonuria: A Fourier-transform infrared microscopy (FTIRM) and low energy X-ray fluorescence (LEXRF) microscopy correlative study on a rare disease. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Lagomarsino, S.; Iotti, S.; Farruggia, G.; Cedola, A.; Trapani, V.; Fratini, M.; Bukreeva, I.; Notargiacomo, A.; Mastrototaro, L.; Marraccini, C.; et al. Intracellular concentration map of magnesium in whole cells by combined use of X-ray fluorescence microscopy and atomic force microscopy. Spectrochim. Acta B At. Spectrosc. 2011, 66, 834–840. [Google Scholar] [CrossRef]
- Malucelli, E.; Iotti, S.; Gianoncelli, A.; Fratini, M.; Merolle, L.; Notargiacomo, A.; Marraccini, C.; Sargenti, A.; Cappadone, C.; Farruggia, G.; et al. Quantitative chemical imaging of the intracellular spatial distribution of fundamental elements and light metals in single cells. Analyt. Chem. 2014, 86, 5108–5115. [Google Scholar] [CrossRef]
- Popescu, B.F.G.; George, M.J.; Bergmann, U.; Garachtchenko, A.V.; Kelly, M.E.; McCrea, R.P.; Lüning, K.; Devon, R.M.; George, G.N.; Hanson, A.D.; et al. Mapping metals in Parkinson’s and normal brain using rapid-scanning X-ray fluorescence. Phys. Med. Biol. 2009, 54, 651. [Google Scholar] [CrossRef] [PubMed]
- James, S.A.; Myers, D.E.; de Jonge, M.D.; Vogt, S.; Ryan, C.G.; Sexton, B.A.; Hoobin, P.; Paterson, D.; Howard, D.L.; Mayo, S.C.; et al. Quantitative comparison of preparation methodologies for x-ray fluorescence microscopy of brain tissue. Anal. Bioanal. Chem. 2011, 401, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Paunesku, T.; Wanzer, M.; Kirillova, E.; Muksinova, K.; Revina, V.; Romanov, S.; Lyubchansky, E.; Grosche, B.; Birschwilks, M.; Vogt, S.; et al. X-ray fluorescence microscopy for investigation of archival tissues. Health Phys. 2012, 103, 181. [Google Scholar] [CrossRef] [PubMed]
- Carboni, E.; Nicolas, J.-D.; Töpperwien, M.; Stadelmann-Nessler, C.; Lingor, P.; Salditt, T. Imaging of neuronal tissues by x-ray diffraction and x-ray fluorescence microscopy: Evaluation of contrast and biomarkers for neurodegenerative diseases. Biomed. Opt. Express 2017, 8, 4331–4347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gherase, M.R.; Fleming, D.E. Probing Trace Elements in Human Tissues with Synchrotron Radiation. Crystals 2020, 10, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malucelli, E.; Procopio, A.; Fratini, M.; Gianoncelli, A.; Notargiacomo, A.; Merolle, L.; Sargenti, A.; Castiglioni, C.; Cappadone, C.; Farruggia, G.; et al. Single cell versus large population analysis: Cell variability in elemental intracellular concentration and distribution. Anal. Bioanal. Chem. 2018, 410, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.T.; Soman, S.S.; Yee, J. Magnesium Balance and Measurement. Adv. Chronic Kidney Dis. 2018, 25, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Mohammed, A. Magnesium: The forgotten electrolyte—A review on hypomagnesemia. Med. Sci. 2019, 7, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Z.G.; Rios, F.J.; Montezano, A.C.; Touyz, R.M. TRPM7, Magnesium, and Signaling. Int. J. Mol. Sci. 2019, 20, 1877. [Google Scholar] [CrossRef] [Green Version]
- Romani, A.M. Magnesium homeostasis in mammalian cells. Front. Biosci. J. Virtual Libr. 2007, 12, 308–331. [Google Scholar] [CrossRef] [PubMed]
- Maguire, M.E. Magnesium transporters: Properties, regulation and structure. Front. Biosci. J. Virtual Libr. 2006, 11, 3149–3163. [Google Scholar] [CrossRef] [PubMed]
- Sargenti, A.; Farruggia, G.; Zaccheroni, N.; Marraccini, C.; Sgarzi, M.; Cappadone, C.; Malucelli, E.; Procopio, A.; Prodi, L.; Lombardo, M.; et al. Synthesis of a highly Mg(2+)-selective fluorescent probe and its application to quantifying and imaging total intracellular magnesium. Nat. Protoc. 2017, 12, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Tao, T.; Li, H.; Zhu, X. mTOR signaling pathway and mTOR inhibitors in cancer: Progress and challenges. Cell Biosci. 2020, 10, 31. [Google Scholar] [CrossRef]
- Narayanankutty, A. PI3K/ Akt/ mTOR Pathway as a Therapeutic Target for Colorectal Cancer: A Review of Preclinical and Clinical Evidence. Curr. Drug Targets 2019, 20, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, R.; Shindo, Y.; Hotta, K.; Suzuki, K.; Oka, K. GABA-induced intracellular Mg2+ mobilization integrates and coordinates cellular information processing for the maturation of neural networks. Curr. Biol. 2018, 28, 3984–3991.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinowska, J.; Małecka, M.; Ciepiela, O. Variations in Magnesium Concentration Are Associated with Increased Mortality: Study in an Unselected Population of Hospitalized Patients. Nutrients 2020, 12, 1836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; del Gobbo, L.C.; Hruby, A.; Rosanoff, A.; He, K.; Dai, Q.; Costello, R.B.; Zhang, W.; Song, Y. The circulating concentration and 24-h urine excretion of magnesium dose-and time-dependently respond to oral magnesium supplementation in a meta-analysis of randomized controlled trials. J. Nutr. 2016, 146, 595–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| x | Expression in Normal Tissue | Expression in Cancer Tissue | |||||
|---|---|---|---|---|---|---|---|
| GTEx | Human Protein Atlas | Human Protein Atlas | TCGA | GEPIA2 | GEO Profiles | Literature | |
| TRPM6 | Mainly expressed in colon, Brain and Testis | Not detected | Not detected | Down-regulated in COAD (FC 30) | Down-regulated in CRC (FC 1.25) [37] Down-regulated in CA (FC 15) [36] | Higher in CRC (IHC) [41] Down-regulated in CRC [43] | |
| TRPM7 | Ubiquitous, expressed in colon | Moderate in colon, only glandular cells | No expression in 11 CRC patients | Higher in CRC (IHC) [41] Higher in CRC (IF) [42] | |||
| MRS2 | Ubiquitous, expressed in colon | High in colon glandular cells and moderate in endothelial cells | Strong and moderate expression in 11/12 CRC patients | Down-regulated in early-onset CRC (FC 1.3) [38] Up-regulated in CA (FC 1.3) [36] | |||
| MAGT1 | Ubiquitous, expressed in colon | High in colon glandular cells and moderate endothelial cells | Moderate expression in 10/12 CRC patients | Up-regulated in COAD (FC 2.5) | Up-regulated in CRC (mRNA) [44] | ||
| SLC41A1 | Ubiquitous, expressed in colon | High in colon glandular cells and moderate in endothelial cells | Strong and moderate expression in 12/12 CRC patients | Up-regulated in early-onset CRC (FC 2.15) [38] Up-regulated in CA (FC 1.21) [36] | |||
| CNNM1 | Low expression in colon | Moderate expression in colon, in both glandular and endothelial cells | Low and moderate expression in 7/12 CRC patients | Down-regulated in CA (FC 1.33) [36] | |||
| CNNM3 | Ubiquitous, expressed in colon | High expression in colon glandular cells, moderate in Colon endothelial cells | Strong and moderate expression in 12/12 CRC patients | ||||
| CNNM4 | Ubiquitous, expressed in colon | High expression in colon, only in glandular cells | Strong and moderate expression in 10/12 CRC patients | Down-regulated in COAD (FC 3.4) | Up-regulated in COAD (FC 3) | Down-regulated in CRC (FC 1.18) [37] Down-regulated in early-onset CRC (FC 1.94) [38] Down-regulated in CA (FC 2.6) [36] | Lower in colon cancer-derived Metastases (IHC) [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiroli, D.; Marraccini, C.; Zanetti, E.; Ragazzi, M.; Gianoncelli, A.; Quartieri, E.; Gasparini, E.; Iotti, S.; Baricchi, R.; Merolle, L. Imbalance of Mg Homeostasis as a Potential Biomarker in Colon Cancer. Diagnostics 2021, 11, 727. https://doi.org/10.3390/diagnostics11040727
Schiroli D, Marraccini C, Zanetti E, Ragazzi M, Gianoncelli A, Quartieri E, Gasparini E, Iotti S, Baricchi R, Merolle L. Imbalance of Mg Homeostasis as a Potential Biomarker in Colon Cancer. Diagnostics. 2021; 11(4):727. https://doi.org/10.3390/diagnostics11040727
Chicago/Turabian StyleSchiroli, Davide, Chiara Marraccini, Eleonora Zanetti, Moira Ragazzi, Alessandra Gianoncelli, Eleonora Quartieri, Elisa Gasparini, Stefano Iotti, Roberto Baricchi, and Lucia Merolle. 2021. "Imbalance of Mg Homeostasis as a Potential Biomarker in Colon Cancer" Diagnostics 11, no. 4: 727. https://doi.org/10.3390/diagnostics11040727