Identification of Brain Damage after Seizures Using an MR-Based Electrical Conductivity Imaging Method

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and N-Methyl-D-Aspartic Acid (NMDA) Treatment
2.2. Assessment of Seizure Severity
2.3. Conductivity Image of Ultra-High-Field (9.4 T) MRI
2.4. Tissue Collection
2.5. Western Blot Analysis
2.6. Nissl Staining
2.7. Immunohistochemistry
2.8. Terminal, Deoxynucleotidyl Transferase-Mediated, dUTP–Biotin, Nick End Labeling (TUNEL) Staining
2.9. Statistical Analysis
3. Results
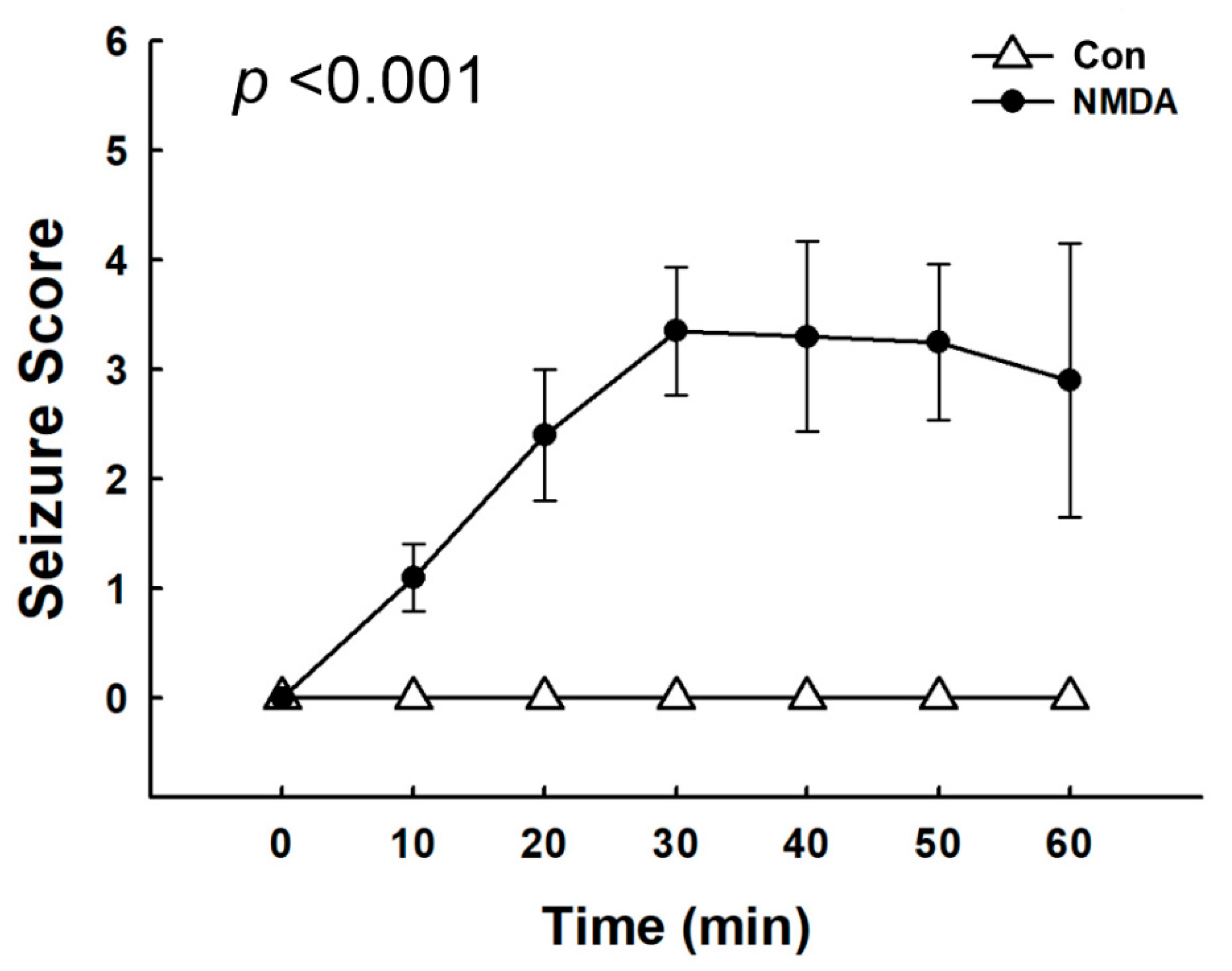
3.1. Seizure Severity after NMDA Treatment
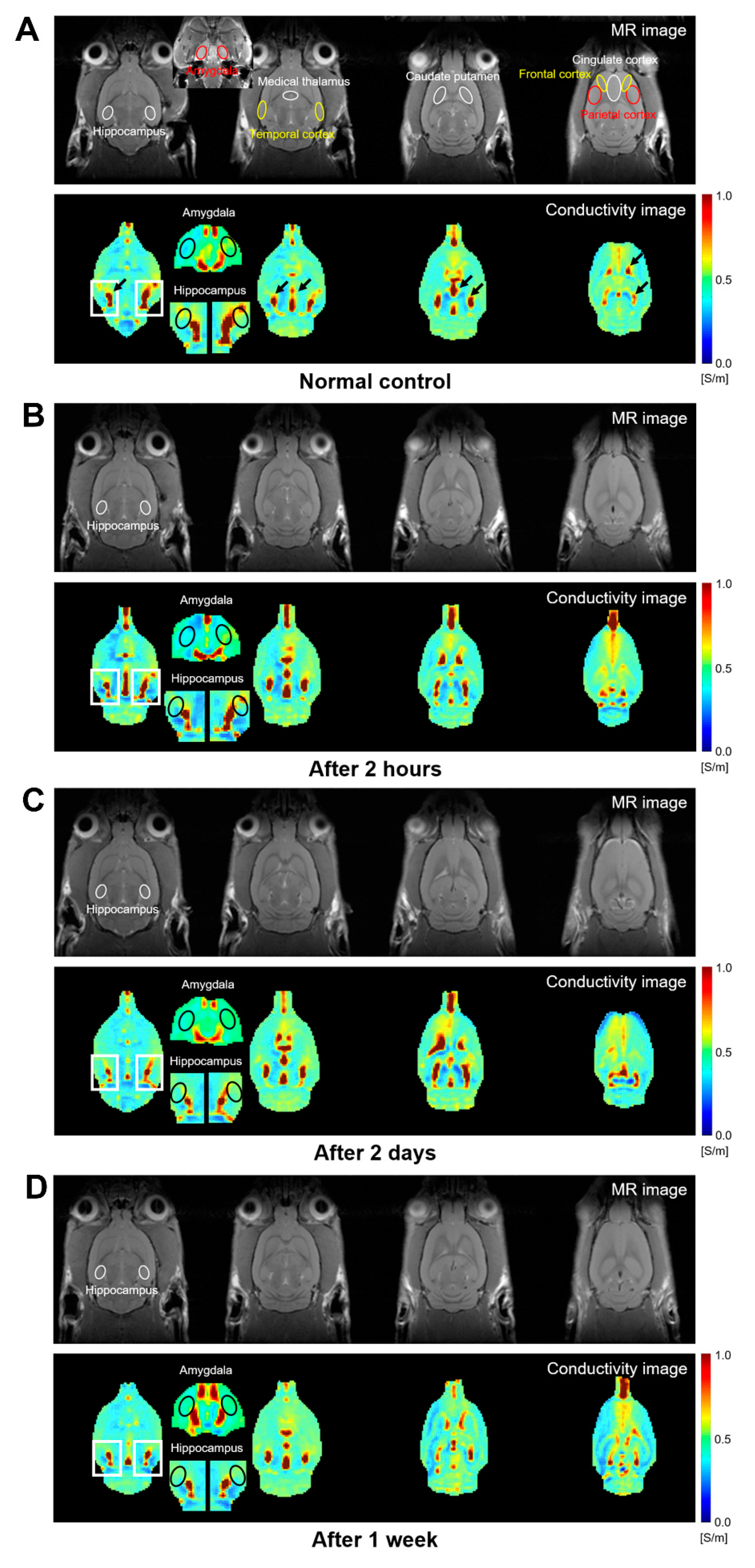
3.2. Change of Conductivity in the Brain of NMDA-Treated Rats
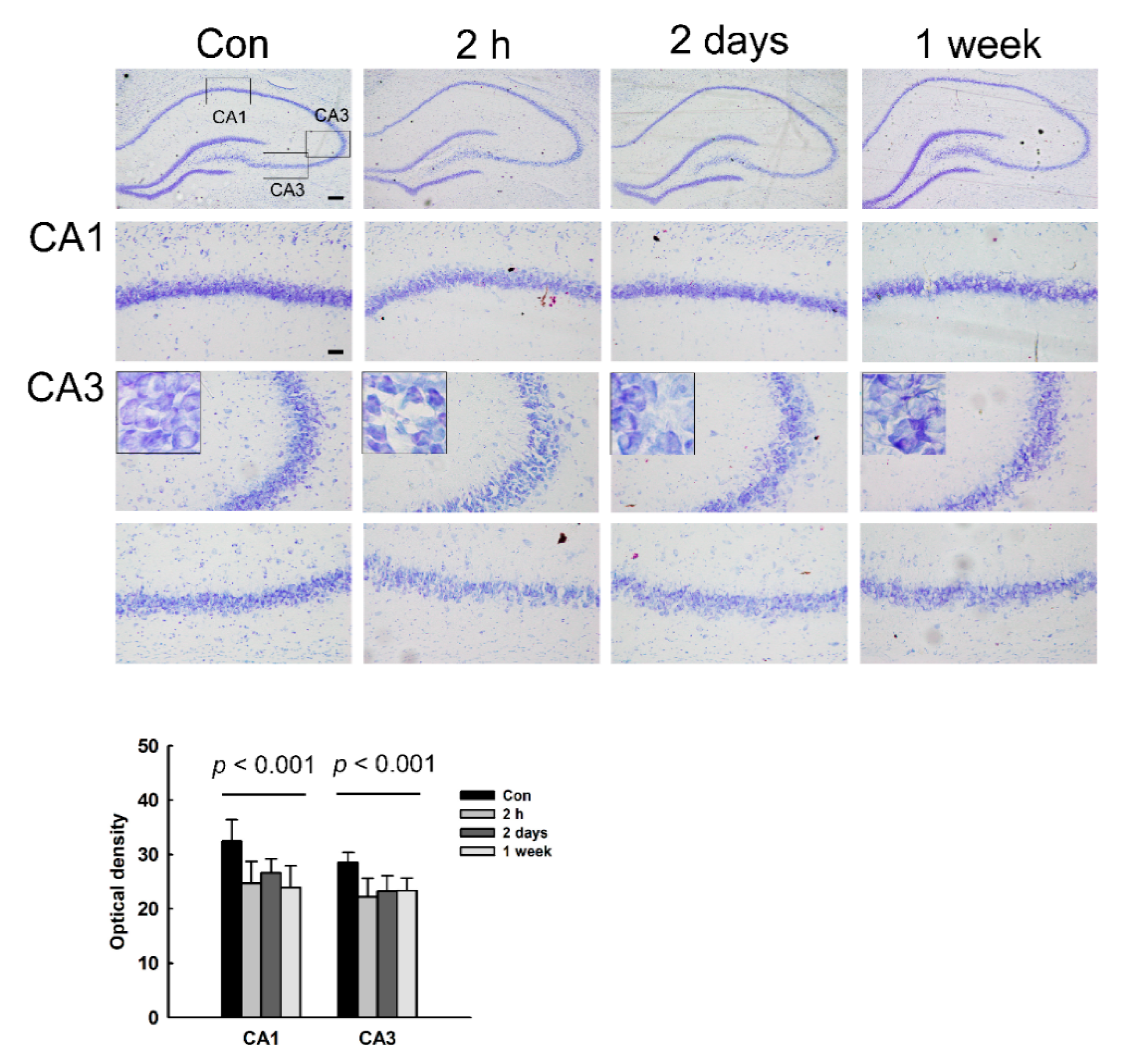
3.3. Morphological Changes and Neuronal Loss in the Hippocampus of NMDA-Treated Rats
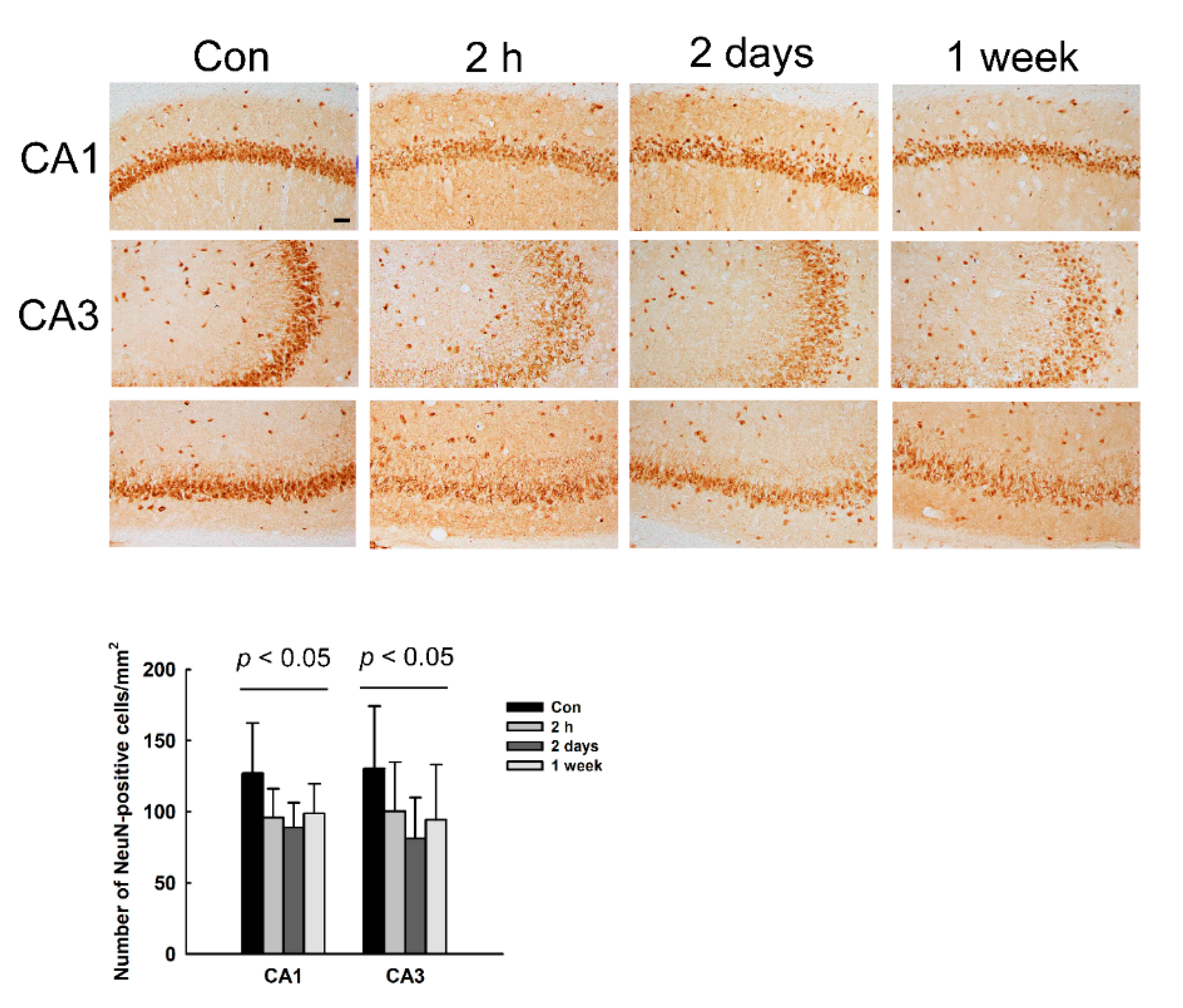
3.4. Decrease of NeuN Immunoreactivity (IR) in the Hippocampus of NMDA-Treated Rats
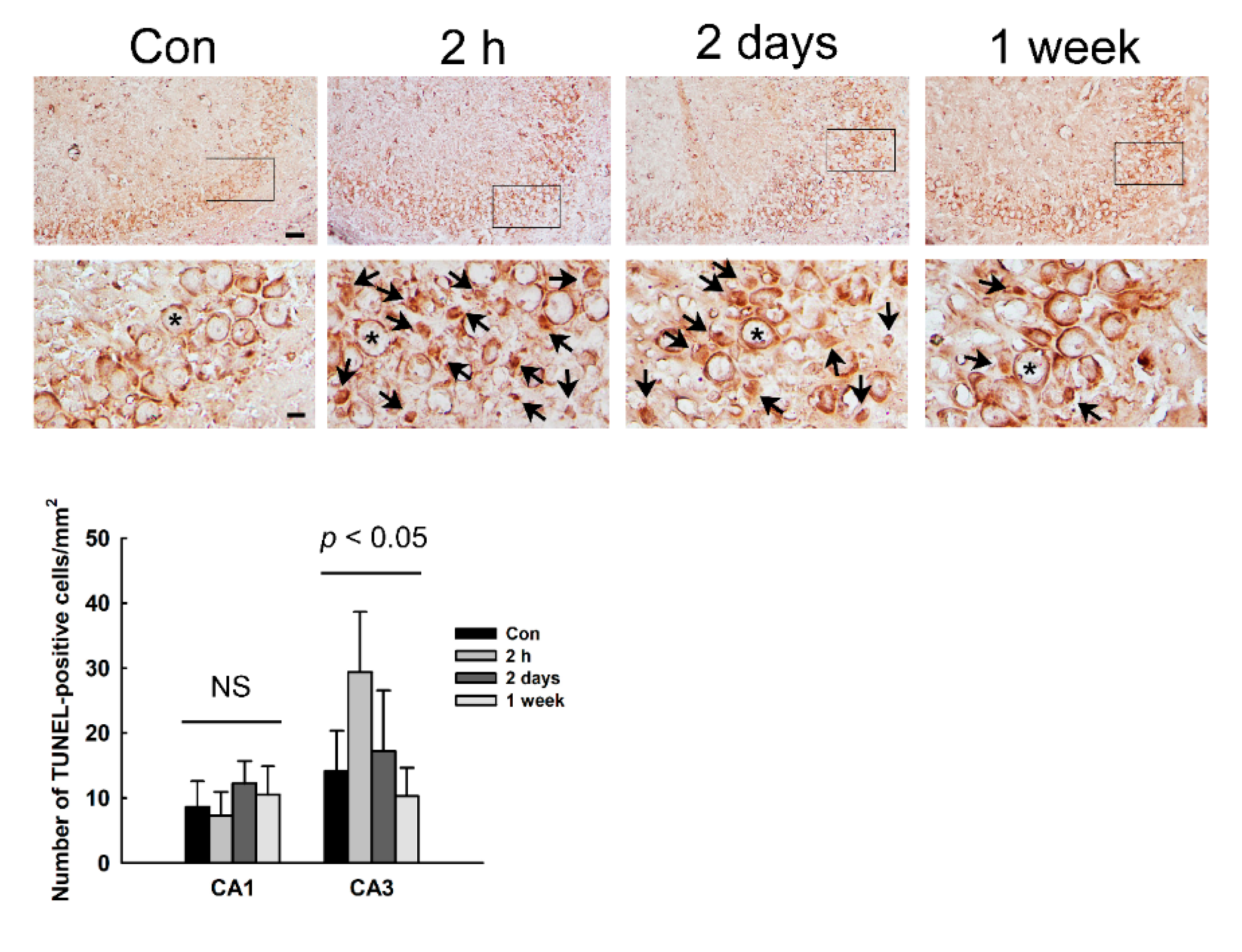
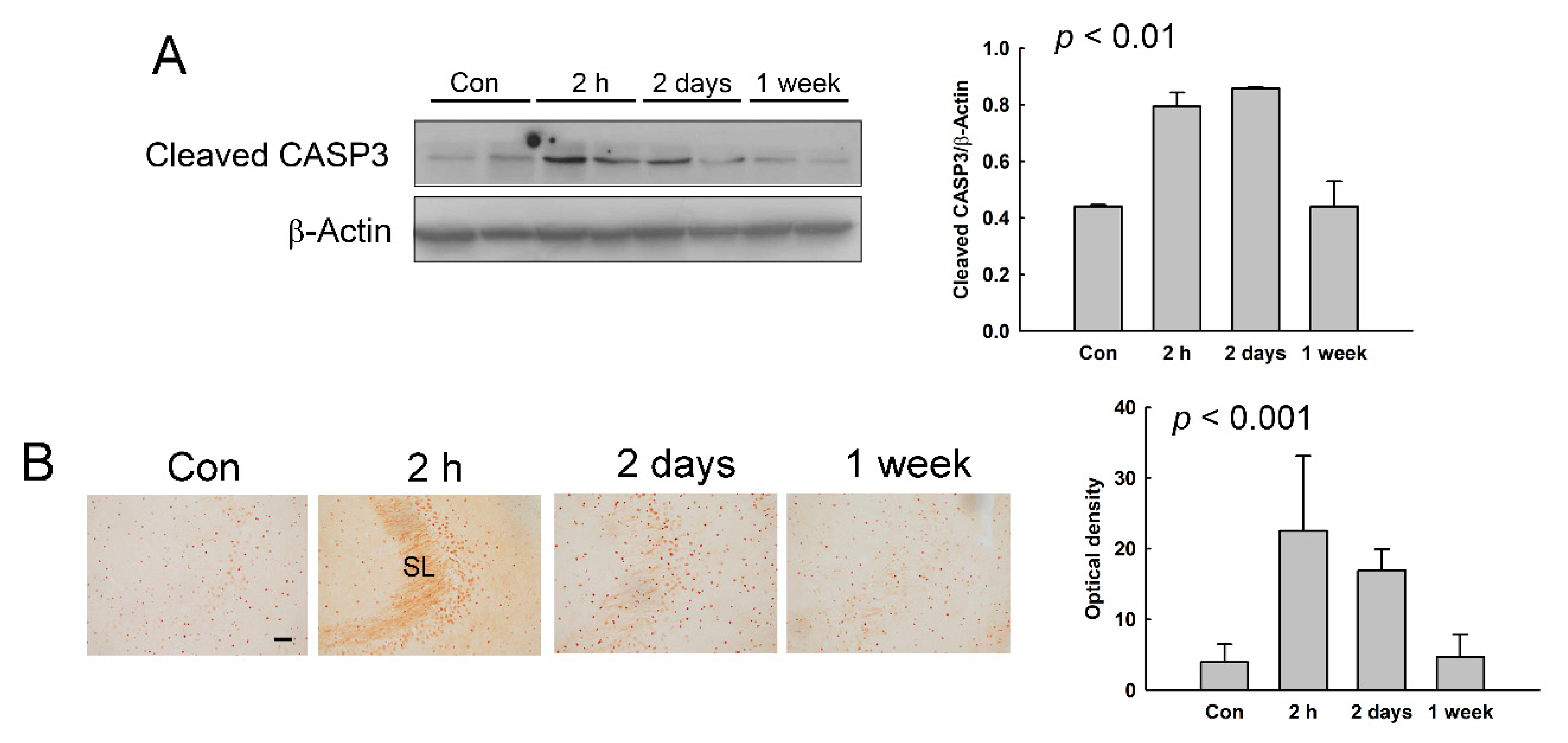
3.5. Induction of Apoptotic Cell Death in the Hippocampus of NMDA-Treated Rats
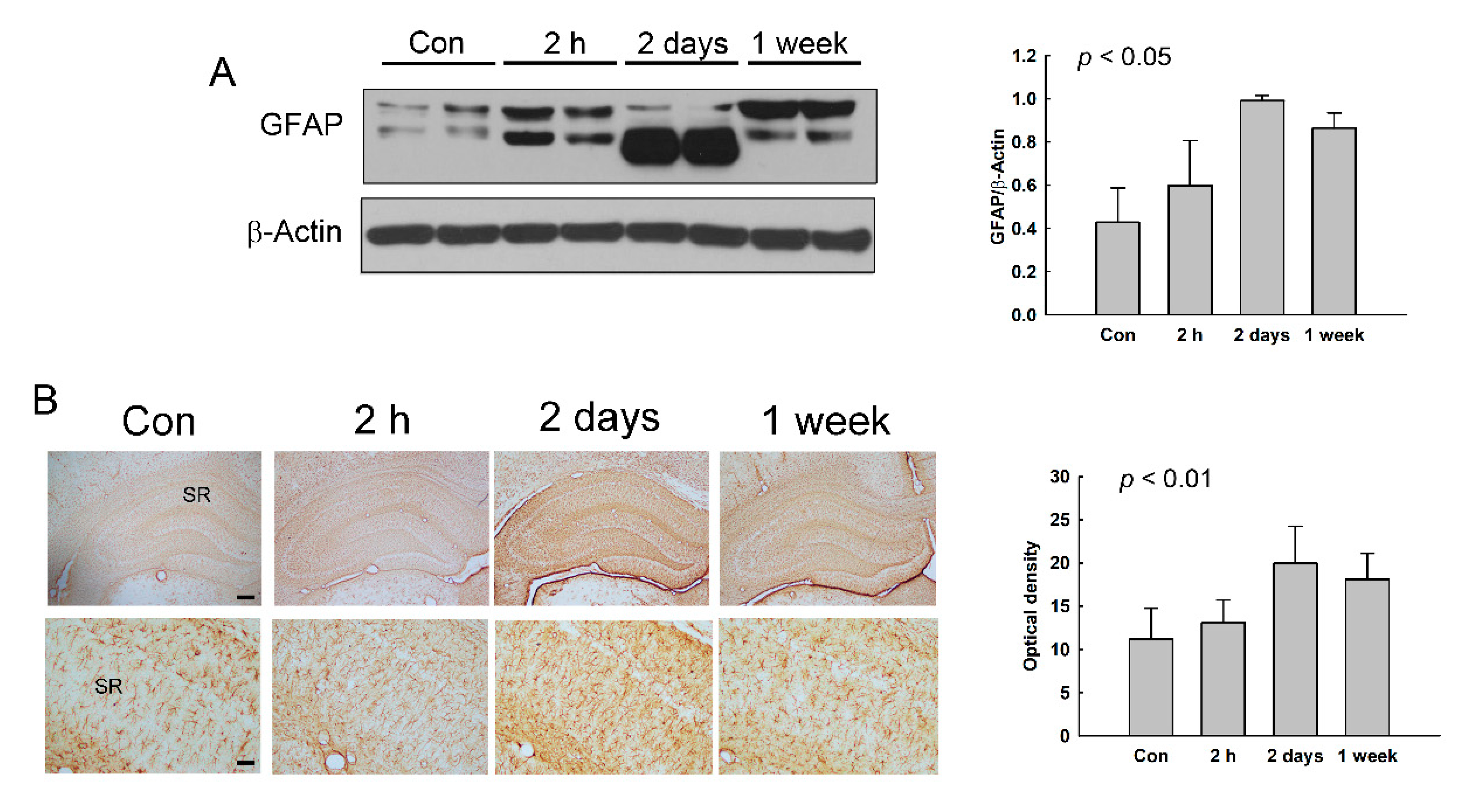
3.6. Increase of Reactive Astrocytes in the Hippocampus of NMDA-Treated Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Stafstrom, C.E.; Carmant, L. Seizures and epilepsy: An overview for neuroscientists. Cold Spring Harb. Perspect. Med. 2015, 5, a022426. [Google Scholar] [CrossRef]
- Chapman, A.G. Glutamate and epilepsy. J. Nutr. 2000, 130, 1043S–1045S. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.C. Pathophysiology of status epilepticus. Neurosci. Lett. 2018, 667, 84–91. [Google Scholar] [CrossRef]
- During, M.J.; Spencer, D.D. Extracellular hippocampal glutamate and spontaneous seizure in the conscious human brain. Lancet 1993, 341, 1607–1610. [Google Scholar] [CrossRef]
- Furness, A.M.; Pal, R.; Michealis, E.K.; Lunte, C.E.; Lunte, S.M. Neurochemical investigation of multiple locally induced seizures using microdialysis sampling: Epilepsy effects on glutamate release. Brain Res. 2019, 1722, 146360. [Google Scholar] [CrossRef]
- Barker-Haliski, M.; White, H.S. Glutamatergic Mechanisms Associated with Seizures and Epilepsy. Cold Spring Harb. Perspect. Med. 2015, 5, a022863. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Puttachary, S.; Thippeswamy, A.; Kanthasamy, A.G.; Thippeswamy, T. Status epilepticus: Behavioral and electroencephalography seizure correlates in kainate experimental models. Front. Neurol. 2018, 9, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, A.; Tymianski, M. Glutamate receptors, neurotoxicity and neurodegeneration. Pflug. Arch. Eur. J. Physiol. 2010, 460, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, H.J.; Park, H.J.; Ra, J.; Zheng, L.T.; Yim, S.V.; Chung, J.H. Protective effect of topiramate on kainic acid-induced cell death in mice hippocampus. Epilepsia 2008, 49, 163–167. [Google Scholar] [CrossRef]
- Cao, J.; Viholainen, J.I.; Dart, C.; Warwick, H.K.; Leyland, M.L.; Courtney, M.J. The PSD95-nNOS interface: A target for inhibition of excitotoxic p38 stress-activated protein kinase activation and cell death. J. Cell Biol. 2005, 168, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Dong, X.X.; Cao, Y.; Liang, Z.Q.; Han, R.; Wu, J.C.; Gu, Z.L.; Qin, Z.H. p53 induction contributes to excitotoxic neuronal death in rat striatum through apoptotic and autophagic mechanisms. Eur. J. Neurosci. 2009, 30, 2258–2270. [Google Scholar] [CrossRef]
- Cianfoni, A.; Caulo, M.; Cerase, A.; Della Marca, G.; Falcone, C.; Di Lella, G.M.; Gaudino, S.; Edwards, J.; Colosimo, C. Seizure-induced brain lesions: A wide spectrum of variably reversible MRI abnormalities. Eur. J. Radiol. 2013, 82, 1964–1972. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Choi, Y.; Joung, H.-Y.; Choi, Y.S.; Kim, H.J.; Joo, Y.; Oh, J.-H.; Hann, H.J.; Cho, Z.-H.; Lee, H.W. Structural and functional alterations at pre-epileptic stage are closely associated with epileptogenesis in pilocarpine-induced epilepsy model. Exp. Neurobiol. 2017, 26, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hübers, A.; Thoma, K.; Schocke, M.; Fauser, S.; Ludolph, A.C.; Kassubek, J.; Pinkhardt, E.H. Acute DWI reductions in patients after single epileptic seizures-more common than assumed. Front. Neurol. 2018, 9, 550. [Google Scholar] [CrossRef] [PubMed]
- Maganti, R.K.; Rutecki, P. EEG and epilepsy monitoring. Continuum 2013, 19, 598–622. [Google Scholar] [CrossRef]
- Hannan, S.; Faulkner, M.; Aristovich, K.; Avery, J.; Holder, D. Frequency-dependent characterisation of impedance changes during epileptiform activity in a rat model of epilepsy. Physiol. Meas. 2018, 39, 085003. [Google Scholar] [CrossRef]
- Kwon, O.I.; Jeong, W.C.; Sajib, S.Z.; Kim, H.J.; Woo, E.J.; Oh, T.I. Reconstruction of dual-frequency conductivity by optimization of phase map in MREIT and MREPT. Biomed. Eng. Online 2014, 13, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Chauhan, M.; Kim, M.O.; Jeong, W.C.; Kim, H.J.; Sersa, I.; Kwon, O.I.; Woo, E.J. Frequency-dependent conductivity contrast for tissue characterization using a dual-frequency range conductivity mapping magnetic resonance method. IEEE Trans. Med. Imaging 2015, 34, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Sajib, S.Z.K.; Kwon, O.I.; Kim, H.J.; Woo, E.J. Electrodeless conductivity tensor imaging (CTI) using MRI: Basic theory and animal experiments. Biomed. Eng. Lett. 2018, 8, 273–282. [Google Scholar] [CrossRef]
- Lüttjohann, A.; Fabene, P.F.; van Luijtelaar, G. A revised Racine’s scale for PTZ-induced seizures in rats. Physiol. Behav. 2009, 98, 579–586. [Google Scholar] [CrossRef]
- Ihara, Y.; Tomonoh, Y.; Deshimaru, M.; Zhang, B.; Uchida, T.; Ishii, A.; Hirose, S. Retigabine, a Kv7. 2/Kv7. 3-channel opener, attenuates drug-induced seizures in knock-in mice harboring Kcnq2 mutations. PLoS ONE 2016, 11, e0150095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bioucas-Dias, J.M.; Valadão, G. Phase unwrapping via graph cuts. IEEE Trans. Image Process. Publ. IEEE Signal Process. Soc. 2007, 16, 698–709. [Google Scholar] [CrossRef] [PubMed]
- Sajib, S.Z.; Katoch, N.; Kim, H.J.; Kwon, O.I.; Woo, E.J. Software toolbox for low-frequency conductivity and current density imaging using MRI. IEEE Trans. Biomed. Eng. 2017, 64, 2505–2514. [Google Scholar] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Meini, A.; Sticozzi, C.; Massai, L.; Palmi, M. A nitric oxide/Ca2+/calmodulin/ERK1/2 mitogen-activated protein kinase pathway is involved in the mitogenic effect of IL-1β in human astrocytoma cells. Br. J. Pharmacol. 2008, 153, 1706–1717. [Google Scholar] [CrossRef] [Green Version]
- Sticozzi, C.; Belmonte, G.; Meini, A.; Carbotti, P.; Grasso, G.; Palmi, M. IL-1β induces GFAP expression in vitro and in vivo and protects neurons from traumatic injury-associated apoptosis in rat brain striatum via NFκB/Ca2+-calmodulin/ERK mitogen-activated protein kinase signaling pathway. Neuroscience 2013, 252, 367–383. [Google Scholar] [CrossRef]
- Kim, D.S.; Kim, J.E.; Kwak, S.E.; Choi, K.C.; Kim, D.W.; Kwon, O.S.; Choi, S.Y.; Kang, T.C. Spatiotemporal characteristics of astroglial death in the rat hippocampo-entorhinal complex following pilocarpine-induced status epilepticus. J. Comp. Neurol. 2008, 511, 581–598. [Google Scholar] [CrossRef]
- Kim, J.-E.; Kang, T.-C. Nucleocytoplasmic p27Kip1 export is required for ERK1/2-mediated reactive astroglial proliferation following status epilepticus. Front. Cell. Neurosci. 2018, 12, 152. [Google Scholar] [CrossRef] [Green Version]
- Voigt, T.; Vaterlein, O.; Stehning, C.; Katscher, U.; Fiehler, J. In vivo glioma characterization using MR conductivity imaging. In Proceedings of the 19th Scientific Meeting of the International Society of Magnetic Resonance in Medicine (ISMRM’11), Montreal, Canada, 7–13 May 2011. [Google Scholar]
- van Lier, A.; Kolk, A.; Brundel, M.; Hendriske, J.; Luijten, J.; Lagendijk, J.; van den Berg, C. Electrical conductivity in ischemic stroke at 7.0 Tesla: A case study. In Proceedings of the 20th Scientific Meeting of the International Society of Magnetic Resonance in Medicine (ISMRM’12), Melbourne, Australia, 5–11 May 2012. [Google Scholar]
- Anderson, C.M.; Swanson, R.A. Astrocyte glutamate transport: Review of properties, regulation, and physiological functions. Glia 2000, 32. [Google Scholar] [CrossRef]
- Simard, M.; Nedergaard, M. The neurobiology of glia in the context of water and ion homeostasis. Neuroscience 2004, 129, 877–896. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Tian, G.-F.; Peng, W.; Lou, N.; Libionka, W.; Han, X.; Nedergaard, M. Astrocyte-mediated control of cerebral blood flow. Nat. Neurosci. 2006, 9, 260–267. [Google Scholar] [CrossRef]
- Ridet, J.; Privat, A.; Malhotra, S.; Gage, F. Reactive astrocytes: Cellular and molecular cues to biological function. Trends Neurosci. 1997, 20, 570–577. [Google Scholar] [CrossRef]
- Horner, P.J.; Gage, F.H. Regenerating the damaged central nervous system. Nature 2000, 407, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.J.; Brady, J.D.; Mohr, C. Astrocyte metabolism and signaling during brain ischemia. Nat. Neurosci. 2007, 10, 1377–1386. [Google Scholar] [CrossRef]
- Panickar, K.S.; Norenberg, M.D. Astrocytes in cerebral ischemic injury: Morphological and general considerations. Glia 2005, 50, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.K.; Woo, E.J. Electrical tissue property imaging at low frequency using MREIT. IEEE Trans. Biomed. Eng. 2014, 61, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Jahng, G.H.; Lee, M.B.; Kim, H.J.; Woo, E.J.; Kwon, O.I. Low-frequency dominant electrical conductivity imaging of in vivo human brain using high-frequency conductivity at Larmor-frequency and spherical mean diffusivity without external injection current. NeuroImage 2021, 225, 117466. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Conductivity (S/m) | p | FDR p | |||
|---|---|---|---|---|---|---|
| Control | 2 h | 2 Days | 1 Week | |||
| Amygdala | 0.434 ± 0.025 | 0.418 ± 0.011 | 0.446 ± 0.025 | 0.480 ± 0.013 | 0.058 | 0.091 |
| Medial thalamus | 0.399 ± 0.016 | 0.392 ± 0.006 | 0.403 ± 0.021 | 0.423 ± 0.010 | 0.131 | 0.234 |
| Hippocampus | 0.501 ± 0.038 | 0.433 ± 0.016 | 0.444 ± 0.013 | 0.442 ± 0.014 | 0.011 * | 0.261 |
| Temporal cortex | 0.441 ± 0.003 | 0.433 ± 0.004 | 0.458 ± 0.015 | 0.466 ± 0.019 | 0.126 | 0.261 |
| Frontal cortex | 0.459 ± 0.013 | 0.455 ± 0.010 | 0.433 ± 0.026 | 0.447 ± 0.015 | 0.516 | 0.510 |
| Parietal cortex | 0.434 ± 0.026 | 0.429 ± 0.005 | 0.415 ± 0.009 | 0.417 ± 0.023 | 0.553 | 0.510 |
| Cingulate cortex | 0.538 ± 0.003 | 0.544 ± 0.050 | 0.536 ± 0.031 | 0.575 ± 0.028 | 0.383 | 0.553 |
| Caudate putamen | 0.420 ± 0.021 | 0.433 ± 0.014 | 0.443 ± 0.040 | 0.465 ± 0.018 | 0.357 | 0.553 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Choi, B.K.; Park, J.A.; Kim, H.J.; Oh, T.I.; Kang, W.S.; Kim, J.W.; Park, H.J. Identification of Brain Damage after Seizures Using an MR-Based Electrical Conductivity Imaging Method. Diagnostics 2021, 11, 569. https://doi.org/10.3390/diagnostics11030569
Kim S, Choi BK, Park JA, Kim HJ, Oh TI, Kang WS, Kim JW, Park HJ. Identification of Brain Damage after Seizures Using an MR-Based Electrical Conductivity Imaging Method. Diagnostics. 2021; 11(3):569. https://doi.org/10.3390/diagnostics11030569
Chicago/Turabian StyleKim, Sanga, Bup Kyung Choi, Ji Ae Park, Hyung Joong Kim, Tong In Oh, Won Sub Kang, Jong Woo Kim, and Hae Jeong Park. 2021. "Identification of Brain Damage after Seizures Using an MR-Based Electrical Conductivity Imaging Method" Diagnostics 11, no. 3: 569. https://doi.org/10.3390/diagnostics11030569