Rehabilitation Improves Mitochondrial Energetics in Progressive Multiple Sclerosis: The Significant Role of Robot-Assisted Gait Training and of the Personalized Intensity


 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Outcome Measures
2.3. Blood Collection and Determination of Mitochondrial Biomarkers
2.4. Endurance Walking Capacity
2.5. Rehabilitation Treatments
2.6. Statistical Analysis
3. Results
3.1. Mitochondrial Function Biomarkers at Baseline
3.2. Mitochondrial Function Biomarkers in Response to Rehabilitation
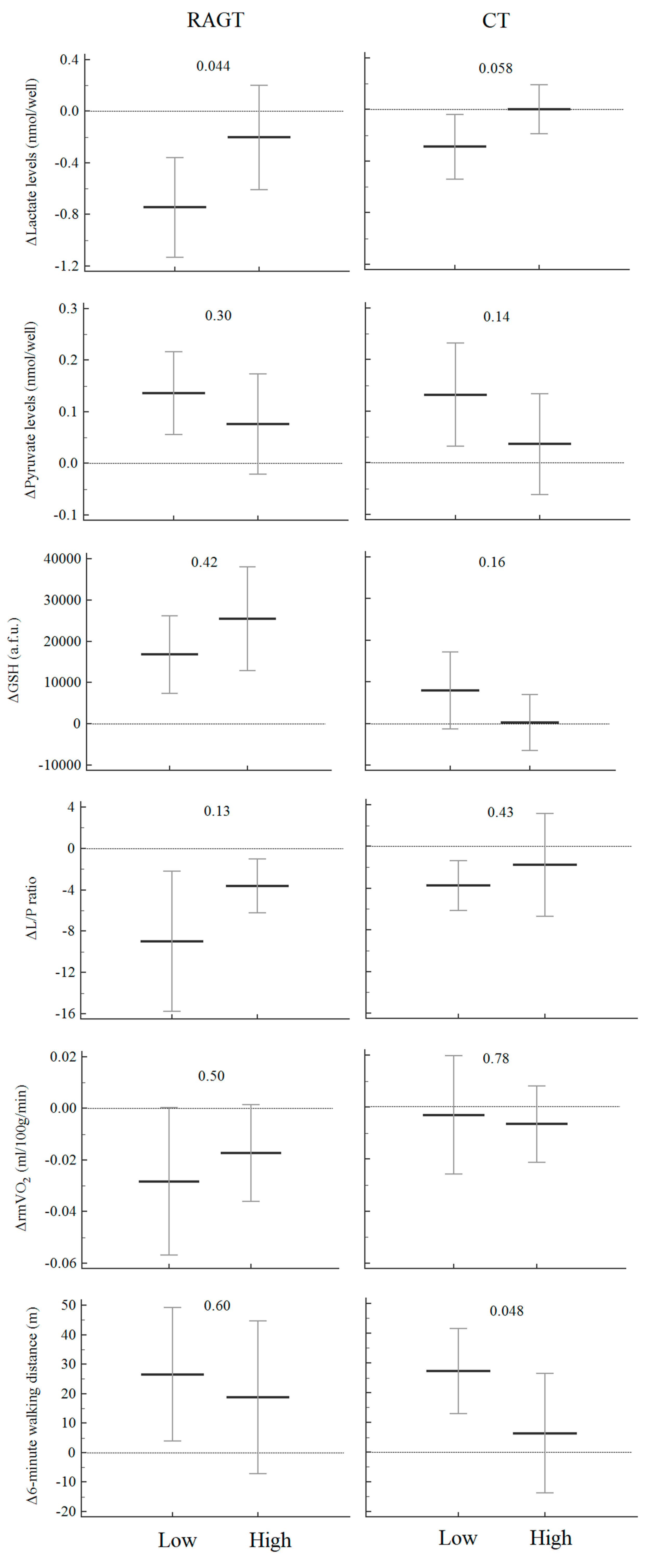
3.3. Mitochondrial Function Biomarker Response to Different Types of Rehabilitation
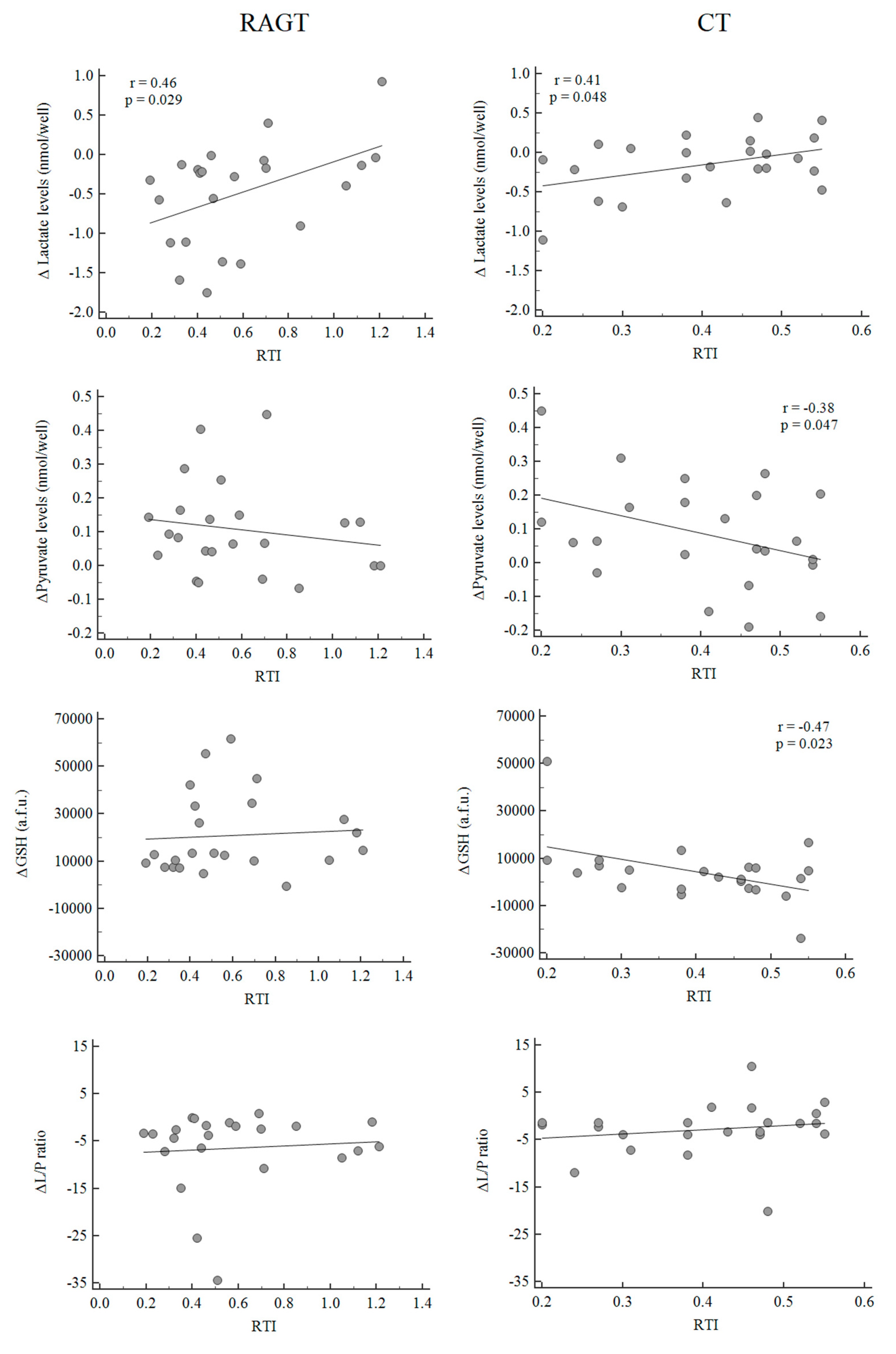
3.4. Mitochondrial Function Biomarkers, Rehabilitative Factors and Endurance Walking Capacity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Patergnani, S.; Fossati, V.; Bonora, M.; Giorgi, C.; Marchi, S.; Missiroli, S.; Rusielewicz, T.; Wieckowski, M.R.; Pinton, P. Mitochondria in Multiple Sclerosis: Molecular Mechanisms of Pathogenesis. Int. Rev. Cell Mol. Biol. 2017, 328, 49–103. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; Marchi, S.; Simoes, I.C.M.; Ren, Z.; Morciano, G.; Perrone, M.; Patalas-Krawczyk, P.; Borchard, S.; Jędrak, P.; Pierzynowska, K.; et al. Mitochondria and Reactive Oxygen Species in Aging and Age-Related Diseases. Int. Rev. Cell Mol. Biol. 2018, 340, 209–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, D.; López-Rodas, G.; Casanova, B.; Marti, M.B. Perturbed glucose metabolism: Insights into multiple sclerosis pathogenesis. Front. Neurol. 2014, 5, 250. [Google Scholar] [CrossRef]
- Adiele, R.C.; Adiele, C.A. Metabolic defects in multiple sclerosis. Mitochondrion 2019, 44, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.R.; Tompkins, S.C.; Taylor, E.B. Regulation of pyruvate metabolism and human disease. Cell. Mol. Life Sci. 2014, 71, 2577–2604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahlin, K. Muscle glucose metabolism during exercise. Ann. Med. 1990, 22, 85–89. [Google Scholar]
- Bonora, M.; Patergnani, S.; Rimessi, A.; De Marchi, E.; Suski, J.M.; Bononi, A.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. ATP synthesis and storage. Purinergic Signal. 2012, 8, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Stallknecht, B.; Vissing, J.; Galbo, H. Lactate production and clearance in exercise. Effects of training. A mini-review. Scand. J. Med. Sci. Sports 1998, 8, 127–131. [Google Scholar] [CrossRef]
- Kaushik, D.K.; Bhattacharya, A.; Mirzaei, R.; Rawji, K.S.; Ahn, Y.; Rho, J.M.; Yong, V.W. Enhanced glycolytic metabolism supports transmigration of brain-infiltrating macrophages in multiple sclerosis. J. Clin. Investig. 2019, 129, 3277–3292. [Google Scholar] [CrossRef]
- Marchi, S.; Giorgi, C.; Suski, J.M.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Missiroli, S.; Patergnani, S.; Poletti, F.; et al. Mitochondria-ros crosstalk in the control of cell death and aging. J. Signal Transduct. 2012, 2012, 329635. [Google Scholar] [CrossRef] [Green Version]
- Beal, M.F. Mitochondria, free radicals, and neurodegeneration. Curr. Opin. Neurobiol. 1996, 6, 661–666. [Google Scholar] [CrossRef]
- Morciano, G.; Bonora, M.; Campo, G.; Aquila, G.; Rizzo, P.; Giorgi, C.; Wieckowski, M.R.; Pinton, P. Mechanistic Role of mPTP in Ischemia-Reperfusion Injury. Adv. Exp. Med. Biol. 2017, 982, 169–189. [Google Scholar] [CrossRef] [PubMed]
- Bonora, M.; Wieckowsk, M.R.; Chinopoulos, C.; Kepp, O.; Kroemer, G.; Galluzzi, L.; Pinton, P. Molecular mechanisms of cell death: Central implication of ATP synthase in mitochondrial permeability transition. Oncogene 2015, 34, 1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilberter, Y.; Gubkina, O.; Ivanov, A.I. A unique array of neuroprotective effects of pyruvate in neuropathology. Front. Neurosci. 2015, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Amorini, A.M.; Nociti, V.; Petzold, A.; Gasperini, C.; Quartuccio, E.; Lazzarino, G.; Di Pietro, V.; Belli, A.; Signoretti, S.; Vagnozzi, R.; et al. Serum lactate as a novel potential biomarker in multiple sclerosis. Biochim Biophys. Acta 2014, 1842, 1137–1143. [Google Scholar] [CrossRef] [Green Version]
- Nijland, P.G.; Molenaar, R.J.; van der Pol, S.M.; Van Der Valk, P.; Van Noorden, C.J.F.; De Vries, H.E.; Van Horssen, J. Differential expression of glucose-metabolizing enzymes in multiple sclerosis lesions. Acta Neuropathol. Commun. 2015, 3, 79. [Google Scholar] [CrossRef] [Green Version]
- Albanese, M.; Zagaglia, S.; Landi, D.; Boffa, L.; Nicoletti, C.G.; Marciani, M.G.; Mandolesi, G.; Marfia, G.A.; Buttari, F.; Mori, F.; et al. Cerebrospinal fluid lactate is associated with multiple sclerosis disease progression. J. Neuroinflamm. 2016, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Castellazzi, M.; Patergnani, S.; Donadio, M.; Giorgi, C.; Bonora, M.; Fainardi, E.; Casetta, I.; Granieri, E.; Pugliatti, M.; Pinton, P. Correlation between auto/mitophagic processes and magnetic resonance imaging activity in multiple sclerosis patients. J. Neuroinflamm. 2019, 16, 131. [Google Scholar] [CrossRef]
- Patergnani, S.; Castellazzi, M.; Bonora, M.; Marchi, S.; Casetta, I.; Pugliatti, M.; Giorgi, C.; Granieri, E.; Pinton, P. Autophagy and mitophagy elements are increased in body fluids of multiple sclerosis-affected individuals. J. Neurol. Neurosurg. Psychiatry 2018, 89, 439–441. [Google Scholar] [CrossRef]
- Kim, H.H.; Jeong, I.H.; Hyun, J.S.; Kong, B.S.; Kim, H.J.; Park, S.J. Metabolomic profiling of CSF in multiple sclerosis and neuromyelitis optica spectrum disorder by nuclear magnetic resonance. PLoS ONE 2017, 12, e0181758. [Google Scholar] [CrossRef]
- Huertas, J.R.; Casuso, R.A.; Agustín, P.H.; Cogliati, S. Stay Fit, Stay Young: Mitochondria in Movement: The Role of Exercise in the New Mitochondrial Paradigm. Oxidative Med. Cell. Longev. 2019, 2019, 7058350. [Google Scholar] [CrossRef] [Green Version]
- Mähler, A.; Steiniger, J.; Bock, M.; Brandt, A.U.; Haas, V.; Boschmann, M.; Paul, F. Is metabolic flexibility altered in multiple sclerosis patients? PLoS ONE 2012, 7, e43675. [Google Scholar] [CrossRef] [PubMed]
- Hortobágyi, T.; Dempsey, L.; Fraser, D.; Zheng, D.; Hamilton, G.; Lambert, J.; Dohm, L. Changes in muscle strength; muscle fibre size and myofibrillar gene expression after immobilization and retraining in humans. J. Physiol. 2000, 524, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Malagoni, A.M.; Felisatti, M.; Lamberti, N.; Basaglia, N.; Manfredini, R.; Salvi, F.; Zamboni, P.; Manfredini, F. Muscle oxygen consumption by NIRS and mobility in multiple sclerosis patients. BMC Neurol. 2013, 13, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greb, E. Patients with multiple sclerosis have altered metabolic flexibility. Neurol. Rev. 2012, 20, 15. [Google Scholar]
- Langeskov-Christensen, M.; Heine, M.; Kwakkel, G.; Dalgas, U. Aerobic capacity in persons with multiple sclerosis: A systematic review and meta-analysis. Sports Med. 2015, 45, 905–923. [Google Scholar] [CrossRef]
- Motl, R.W.; Goldman, M. Physical inactivity; neurological disability; and cardiorespiratory fitness in multiple sclerosis. Acta Neurol. Scand. 2011, 123, 98–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gusdon, A.M.; Callio, J.; Distefano, G.; O’Doherty, R.M.; Goodpaster, B.H.; Coen, P.M.; Chu, C.T. Exercise increases mitochondrial complex I activity and DRP1 expression in the brains of aged mice. Exp. Gerontol. 2017, 90, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, J.L.; Murphy, E.A.; McClellan, J.L.; Carmichael, M.D.; Davis, J.M. Exercise training increases mitochondrial biogenesis in the brain. J. Appl. Physiol. 2011, 111, 1066–1071. [Google Scholar] [CrossRef] [Green Version]
- AAHPERD. Physical Education for Lifelong Fitness: The Physical Best Teacher’s Guide; Human Kinetics: Champaign, IL, USA, 1999; pp. 78–79. [Google Scholar]
- Pedersen, B.K. Physical activity and muscle-brain crosstalk. Nat. Rev. Endocrinol. 2019, 15, 383–392. [Google Scholar] [CrossRef]
- Straudi, S.; Manfredini, F.; Lamberti, N.; Martinuzzi, C.; Maietti, E.; Basaglia, N. Robot-assisted gait training is not superior to intensive overground walking in multiple sclerosis with severe disability (the RAGTIME study): A randomized controlled trial. Mult. Scler. J. 2020, 26, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Straudi, S.; Manfredini, F.; Lamberti, N.; Zamboni, P.; Bernardi, F.; Marchetti, G.; Pinton, P.; Bonora, M.; Secchiero, P.; Tisato, V.; et al. The effectiveness of Robot-Assisted Gait Training versus conventional therapy on mobility in severely disabled progressIve MultiplE sclerosis patients (RAGTIME): Study protocol for a randomized controlled trial. Trials 2017, 18, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, T.E.; Southern, W.M.; Reynolds, M.A.; McCully, K.K. A cross-validation of near-infrared spectroscopy measurements of skeletal muscle oxidative capacity with phosphorus magnetic resonance spectroscopy. J. Appl. Physiol. (1985) 2013, 115, 1757–1766. [Google Scholar] [CrossRef] [Green Version]
- Manfredini, F.; Lamberti, N.; Malagoni, A.M.; Felisatti, M.; Zuccalà, A.; Torino, C.; Tripepi, G.; Catizone, L.; Mallamaci, F.; Zoccali, C.; et al. The role of deconditioning in the end-stage renal disease myopathy: Physical exercise improves altered resting muscle oxygen consumption. Am. J. Nephrol. 2015, 41, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Malagoni, A.M.; Felisatti, M.; Mandini, S.; Mascoli, F.; Manfredini, R.; Basaglia, N.; Zamboni, P.; Manfredini, F. Resting muscle oxygen consumption by near-infrared spectroscopy in peripheral arterial disease: A parameter to be considered in a clinical setting? Angiology 2010, 61, 530–536. [Google Scholar] [CrossRef]
- Cederberg, K.L.J.; Sikes, E.M.; Bartolucci, A.A.; Motl, R.W. Walking endurance in multiple sclerosis: Meta-analysis of six-minute walk test performance. Gait Posture 2019, 73, 147–153. [Google Scholar] [CrossRef]
- Janikiewicz, J.; Szymański, J.; Malinska, D.; Patalas-Krawczyk, P.; Michalska, B.; Duszyński, J.; Giorgi, C.; Bonora, M.; Dobrzyn, A.; Wieckowski, M.R. Mitochondria-associated membranes in aging and senescence: Structure; function; and dynamics. Cell Death Dis. 2018, 9, 332. [Google Scholar] [CrossRef]
- Bonora, M.; De Marchi, E.; Patergnani, S.; Suski, J.M.; Celsi, F.; Bononi, A.; Giorgi, C.; Marchi, S.; Rimessi, A.; Duszynski, J.; et al. Tumor necrosis factor-α impairs oligodendroglial differentiation through a mitochondria-dependent process. Cell Death Differ. 2014, 21, 1198–1208. [Google Scholar] [CrossRef] [Green Version]
- Kestenbaum, B.; Gamboa, J.; Liu, S.; Ali, A.S.; Shankland, E.; Jue, T.; Giulivi, C.; Smith, L.R.; Himmelfarb, J.; De Boer, I.H.; et al. Impaired skeletal muscle mitochondrial bioenergetics and physical performance in chronic kidney disease. JCI Insight. 2020, 5, e133289. [Google Scholar] [CrossRef]
- Rimmer, J.H.; Schiller, W.; Chen, M.D. Effects of disability-associated low energy expenditure deconditioning syndrome. Exerc. Sport Sci. Rev. 2012, 40, 22–29. [Google Scholar] [CrossRef]
- Bogdanis, G.C. Effects of physical activity and inactivity on muscle fatigue. Front. Physiol. 2012, 3, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, H.; Deminice, R.; Yoshihara, T.; Powers, S.K. Mitochondrial dysfunction induces muscle atrophy during prolonged inactivity: A review of the causes and effects. Arch. Biochem. Biophys. 2019, 662, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.L.; Aagaard, P. Myosin heavy chain IIX overshoot in human skeletal muscle. Muscle Nerve. 2000, 23, 1095–1104. [Google Scholar] [CrossRef]
- Stein, T.P.; Wade, C.E. Metabolic consequences of muscle disuse atrophy. J. Nutr. 2005, 135, 1824S–1828S. [Google Scholar] [CrossRef]
- Degens, H.; Always, S.E. Control of muscle size during disuse; disease; and aging. Int. J. Sports Med. 2006, 27, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Hikida, R.S.; Staron, R.S.; Hagerman, F.C.; Walsh, S.; Kaiser, E.; Shell, S.; Hervey, S. Effects of high-intensity resistance training on untrained older men. II. Muscle fiber characteristics and nucleo-cytoplasmic relationships. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2000, 55, B347–B354. [Google Scholar] [CrossRef] [Green Version]
- Häkkinen, K.; Newton, R.U.; Gordon, S.E.; McCormick, M.; Volek, J.S.; Nindl, B.C.; Gotshalk, L.A.; Campbell, W.W.; Evans, W.J.; Häkkinen, A.; et al. Changes in muscle morphology; electromyographic activity; and force production characteristics during progressive strength training in young and older men. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 1998, 53, B415–B423. [Google Scholar] [CrossRef] [Green Version]
- Wens, I.; Dalgas, U.; Vandenabeele, F.; Grevendonk, L.; Verboven, K.; Hansen, D.; Eijnde, B.O. High Intensity Exercise in Multiple Sclerosis: Effects on Muscle Contractile Characteristics and Exercise Capacity, a Randomised Controlled Trial. PLoS ONE 2015, 10, e0133697. [Google Scholar] [CrossRef]
- Lefeber, N.; Swinnen, E.; Kerckhofs, E. The immediate effects of robot-assistance on energy consumption and cardiorespiratory load during walking compared to walking without robot-assistance: A systematic review. Disabil. Rehabil. Assist. Technol. 2017, 12, 657–671. [Google Scholar] [CrossRef]
- Devasahayam, A.J.; Downer, M.B.; Ploughman, M. The Effects of Aerobic Exercise on the Recovery of Walking Ability and Neuroplasticity in People with Multiple Sclerosis: A Systematic Review of Animal and Clinical Studies. Mult. Scler. Int. 2017, 2017, 4815958. [Google Scholar] [CrossRef]
- Xie, X.; Sun, H.; Zeng, Q.; Lu, P.; Zhao, Y.; Fan, T.; Huang, G. Do Patients with Multiple Sclerosis Derive More Benefit from Robot-Assisted Gait Training Compared with Conventional Walking Therapy on Motor Function? A Meta-analysis. Front. Neurol. 2017, 8, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, T.L.; Turner, A.P.; Williams, R.M.; Bowen, J.D.; Hatzakis, M.; Rodriguez, A.; Haselkorn, J.K. Correlates of pain interference in multiple sclerosis. Rehabil. Psychol. 2006, 51, 166–174. [Google Scholar] [CrossRef]
- Rohrig, M. Physical Therapy in Multiple Sclerosis. Clin. Bull. Natl. Mult. Scler. Soc. 2018, 1–16. [Google Scholar]
- Lamberti, N.; Straudi, S.; Donadi, M.; Tanaka, H.; Basaglia, N.; Manfredini, F. Effectiveness of blood flow-restricted slow walking on mobility in severe multiple sclerosis: A pilot randomized trial. Scand. J. Med. Sci. Sports 2020. [Google Scholar] [CrossRef]
- Hansen, D.; Feys, P.; Wens, I.; Eijnde, B.O. Is walking capacity in subjects with multiple sclerosis primarily related to muscle oxidative capacity or maximal muscle strength? A pilot study. Mult. Scler. Int. 2014, 2014, 759030. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| MS (n = 46) | Healthy (n = 10) | p | |
|---|---|---|---|
| Lactate, nmol/well | 1.89 ± 0.52 | 1.24 ± 0.76 | 0.001 |
| Pyruvate, nmol/well | 0.26 ± 0.12 | 0.65 ± 0.18 | <0.001 |
| GSH, a.f.u. | 40,896 ± 21,968 | 84,420 ± 25,181 | <0.001 |
| Lactate/Pyruvate ratio | 9.55 ± 7.12 | 1.75 ± 0.73 | 0.001 |
| Whole Population (n = 46) | RAGT (n = 23) | CT (n = 23) | p | |
|---|---|---|---|---|
| Sessions, n | 11.7 ± 0.7 | 11.8 ± 0.7 | 11.7 ± 0.7 | 0.53 |
| Average walking speed, kmh−1 | 1.3 ± 0.4 | 1.8 ± 0.1 | 0.8 ± 0.6 | <0.001 |
| Total distance walked, m | 8162 ± 3664 | 10217 ± 1109 | 6108 ± 4171 | <0.001 |
| Body weight support, % | 44 ± 8 | 44 ± 8 | n.a. | n.a. |
| Relative Training Intensity *, a.u. | 0.54 ± 0.33 | 0.68 ± 0.41 | 0.40 ± 0.11 | 0.003 |
| T0 | T1 | T2 | |
|---|---|---|---|
| Lactate, nmol/well | 1.89 (1.73–2.04) | 1.75 * (1.61–1.89) | 1.52 * † (1.37–1.68) |
| Pyruvate, nmol/well | 0.26 (0.22–0.30) | 0.34 * (0.30–0.37) | 0.35 * (0.32–0.39) |
| GSH, a.f.u. | 40,896 (34,373–47,420) | 43,713 * (36,747–50,678) | 52,859 * † (46,002–59,716) |
| Lactate/pyruvate ratio | 9.55 (7.44–11.67) | 5.89 * (5.09–6.86) | 4.93 * † (4.11–5.75) |
| rmVO2, mlO2/min/100 g | 0.06 (0.05–0.07) | 0.06 (0.04–0.07) | 0.05 * (0.04–0.06) |
| 6MWD, m | 150 (121–180) | 164 * (134–194) | 170 * (137–204) |
| RAGT (n = 23) | CT (n = 23) | |||||
|---|---|---|---|---|---|---|
| T0 | T1 | T2 | T0 | T1 | T2 | |
| Lactate, nmol/well | 2.00 (1.76–2.23) | 1.71 * (1.52–1.91) | 1.42 * † (1.19–1.66) | 1.77 (1.57–1.98) | 1.79 (1.58–2.00) | 1.63 * † (1.41–1.85) |
| Pyruvate, nmol/well | 0.26 (0.21–0.32) | 0.37 * (0.31–0.42) | 0.38 * (0.33–0.42) | 0.26 (0.21–0.31) | 0.31 * (0.26–0.36) | 0.33 * (0.27–0.38) |
| GSH, a.f.u. | 39,566 (30,487–48,645) | 45,214 (34,746–55,681) | 59,286 * † (50,688–67,884) | 42,227 (32,154–52,300) | 42,212 (32,212–52,212) | 46,432 (35,762–57,102) |
| Lactate/pyruvate ratio | 10.59 (6.87–14.30) | 5.30 * (4.14–6.46) | 4.16 * † (3.33–5.00) | 8.52 (6.25–10.78) | 6.48 * (5.35–7.59) | 5.70 * (4.29–7.11) |
| rmVO2, mlO2/min/100 g | 0.07 (0.06–0.09) | 0.06 (0.04–0.08) | 0.05 * (0.04–0.06) | 0.06 (0.04–0.07) | 0.05 (0.03–0.07) | 0.05 (0.03–0.07) |
| 6MWD, m | 151 (114–187) | 168 * (129–207) | 173 * (127–218) | 150 (100–200) | 160 * (111–210) | 167 * (114–220) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manfredini, F.; Straudi, S.; Lamberti, N.; Patergnani, S.; Tisato, V.; Secchiero, P.; Bernardi, F.; Ziliotto, N.; Marchetti, G.; Basaglia, N.; et al. Rehabilitation Improves Mitochondrial Energetics in Progressive Multiple Sclerosis: The Significant Role of Robot-Assisted Gait Training and of the Personalized Intensity. Diagnostics 2020, 10, 834. https://doi.org/10.3390/diagnostics10100834
Manfredini F, Straudi S, Lamberti N, Patergnani S, Tisato V, Secchiero P, Bernardi F, Ziliotto N, Marchetti G, Basaglia N, et al. Rehabilitation Improves Mitochondrial Energetics in Progressive Multiple Sclerosis: The Significant Role of Robot-Assisted Gait Training and of the Personalized Intensity. Diagnostics. 2020; 10(10):834. https://doi.org/10.3390/diagnostics10100834
Chicago/Turabian StyleManfredini, Fabio, Sofia Straudi, Nicola Lamberti, Simone Patergnani, Veronica Tisato, Paola Secchiero, Francesco Bernardi, Nicole Ziliotto, Giovanna Marchetti, Nino Basaglia, and et al. 2020. "Rehabilitation Improves Mitochondrial Energetics in Progressive Multiple Sclerosis: The Significant Role of Robot-Assisted Gait Training and of the Personalized Intensity" Diagnostics 10, no. 10: 834. https://doi.org/10.3390/diagnostics10100834