Distribution and Ecology of Cyanobacteria in the Rocky Littoral of an English Lake District Water Body, Devoke Water

Abstract
:1. Introduction
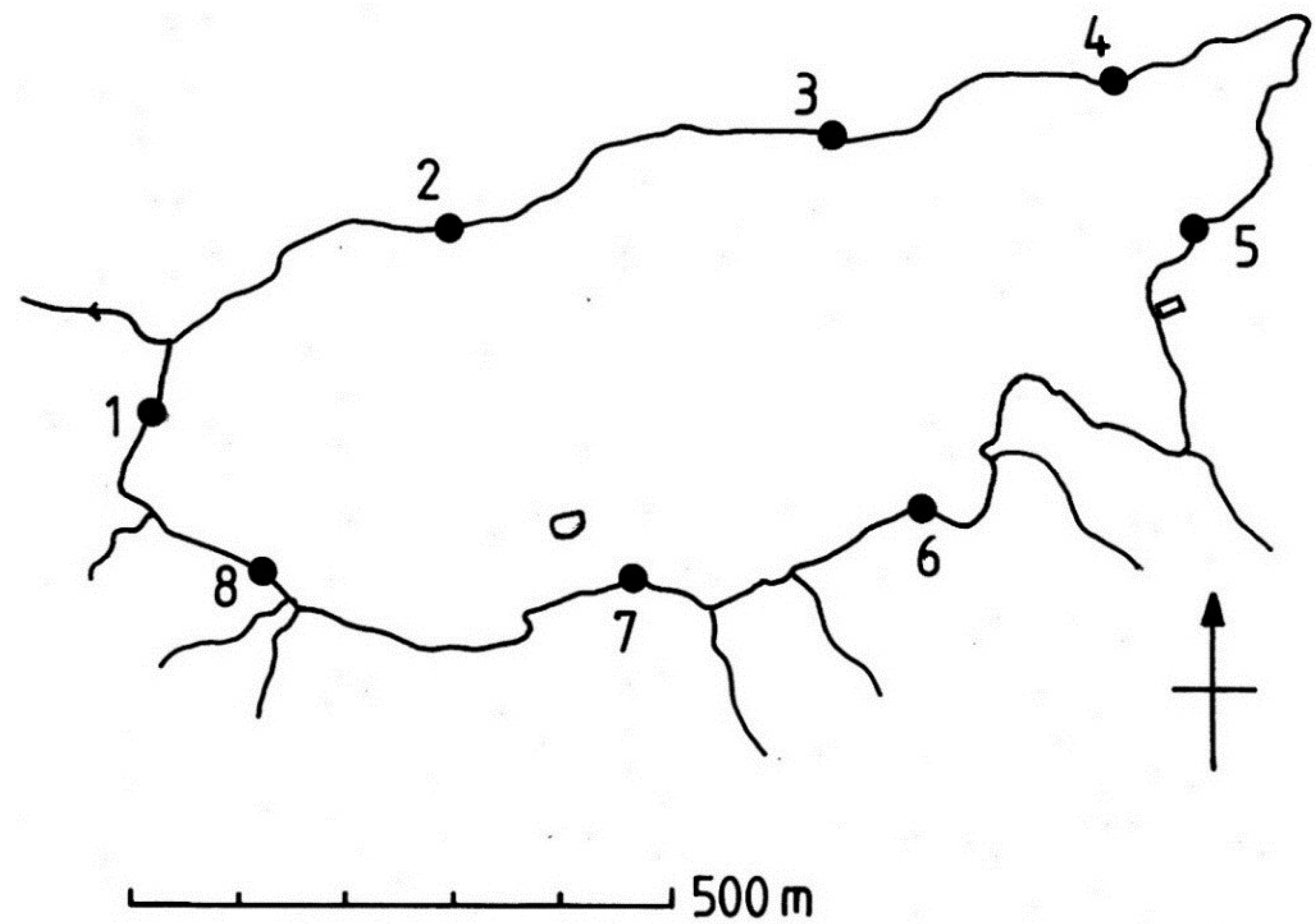
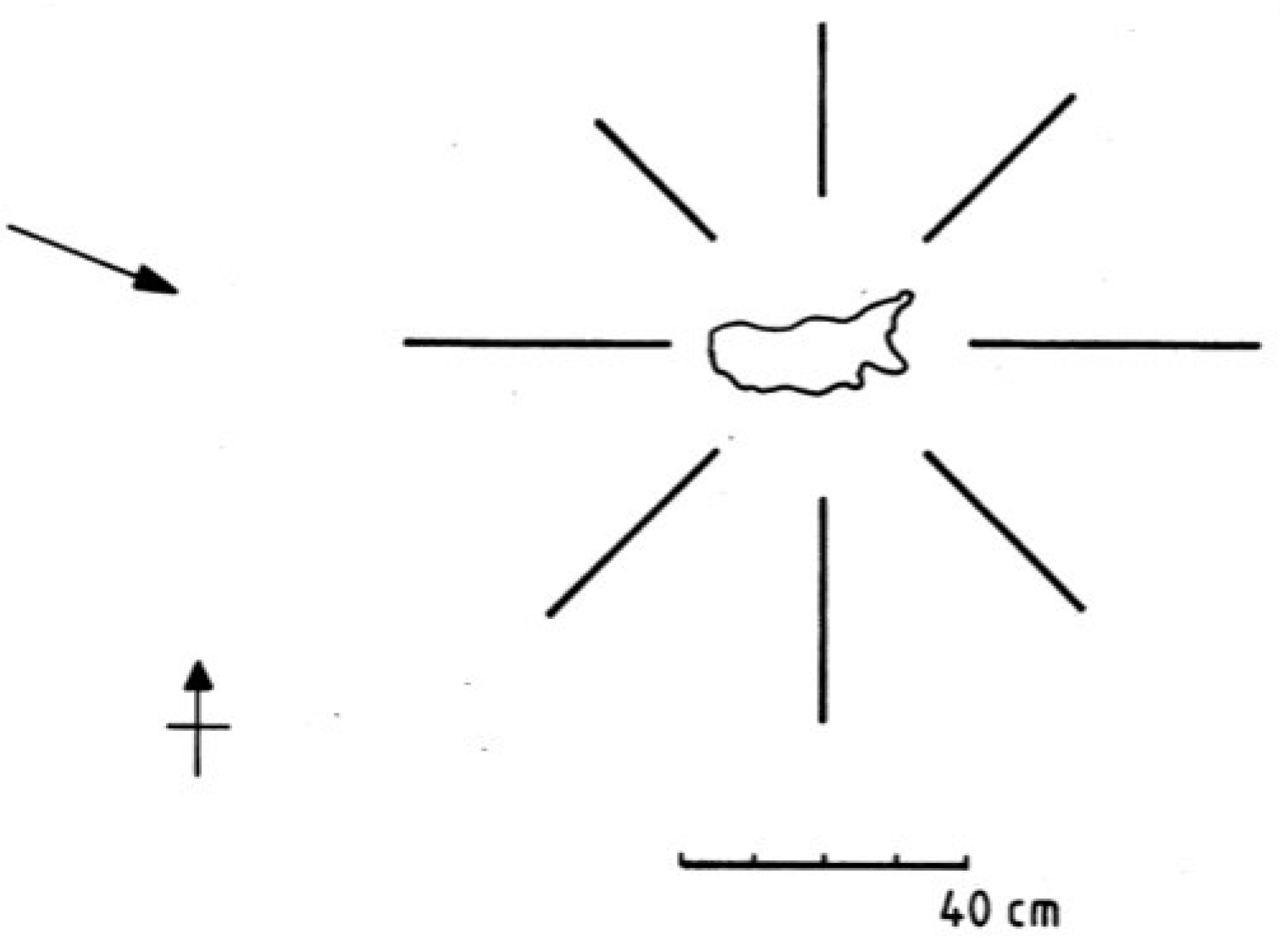
2. Experimental Section


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Value |
|---|---|
| Lat/long | 54°21'N, 3°18'W |
| National grid reference | 34/158969 |
| Altitude | 244 m |
| Surface area | 35 ha |
| Maximum depth | 16 m |
| pH | 6.8 |
| Alkalinity | 66 μeq/L |

3. Results and Discussion
3.1. Results
3.1.1. General Flora
| Order/Genus | Species/Variety | Position Range in Relation to Water Level | Frequency of Occurrence |
|---|---|---|---|
| Chroococcales | |||
| Aphanocapsa | fonticola | 31 | |
| A. | grevillei | −40, +32 | 8 |
| Aphanothece | elabens | −40, −2 | 10 |
| A. | microscopica | −10, −20 | 2 |
| Chroococcus | minutus | −12 | 1 |
| C. | pallidus | −12, −50 | 5 |
| C. | turgidus | −2, −25 | 2 |
| Gloeocapsa | compacta | 46 | |
| G. | granosa | −30 | 1 |
| G. | kuetzingiana | 32 | |
| G. | punctata | 18 | |
| G. | rupicola | 52 | |
| Hydrococcus | rivularis | −10, +10 | 3 |
| Merismopedia | punctata | −30, +18 | 12 |
| Oscillatoriales | |||
| Ammatoidea | normanii | 0, +10 | 3 |
| Homoeothrix | fusca | −4, +32 | 16 |
| Lyngbya | lagerheimii s.l. | 71 | |
| Microcoleus | lacustris | 0, +32 | 3 |
| Schizothrix | calcicola | −12, 0 | 2 |
| S. | fuscescens | 0, +28 | 9 |
| S. | heufleri | −6, +12 | 2 |
| Nostocales | |||
| Calothrix | braunii | −8, −16 | 1 |
| C. | epiphytica | −10 | 1 |
| C. | parietina | 47 | |
| Dichothrix | gypsophila | +22 | 1 |
| D. | orsiniana | 33 | |
| Nostoc | sp. | −16 | 1 |
| Scytonema | mirabile | 0, +30 | 7 |
| S. | myochrous | +10, +16 | 2 |
| Stigonematales | |||
| Stigonema | mamillosum | 31 | |
| S. | ocellatum | 15 |
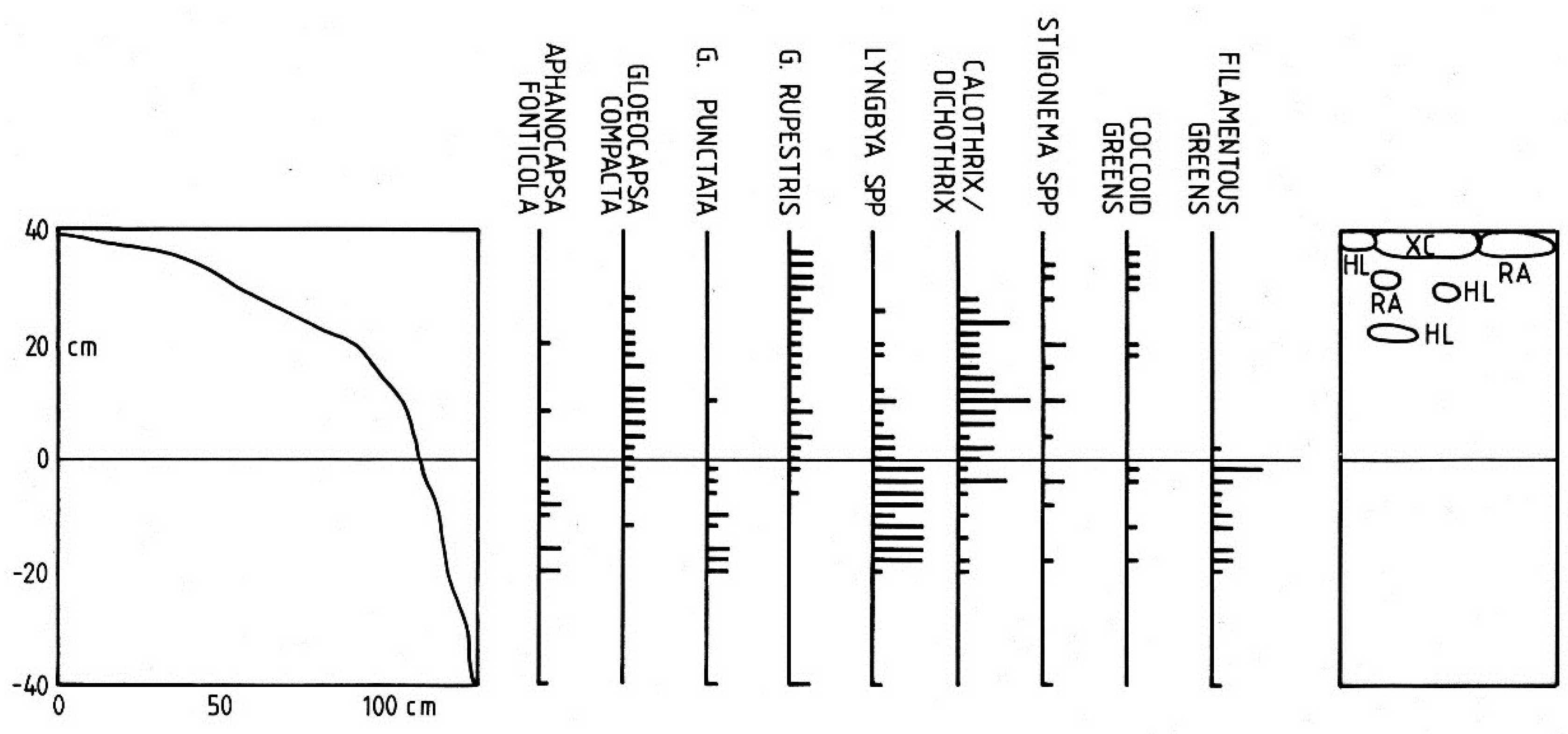
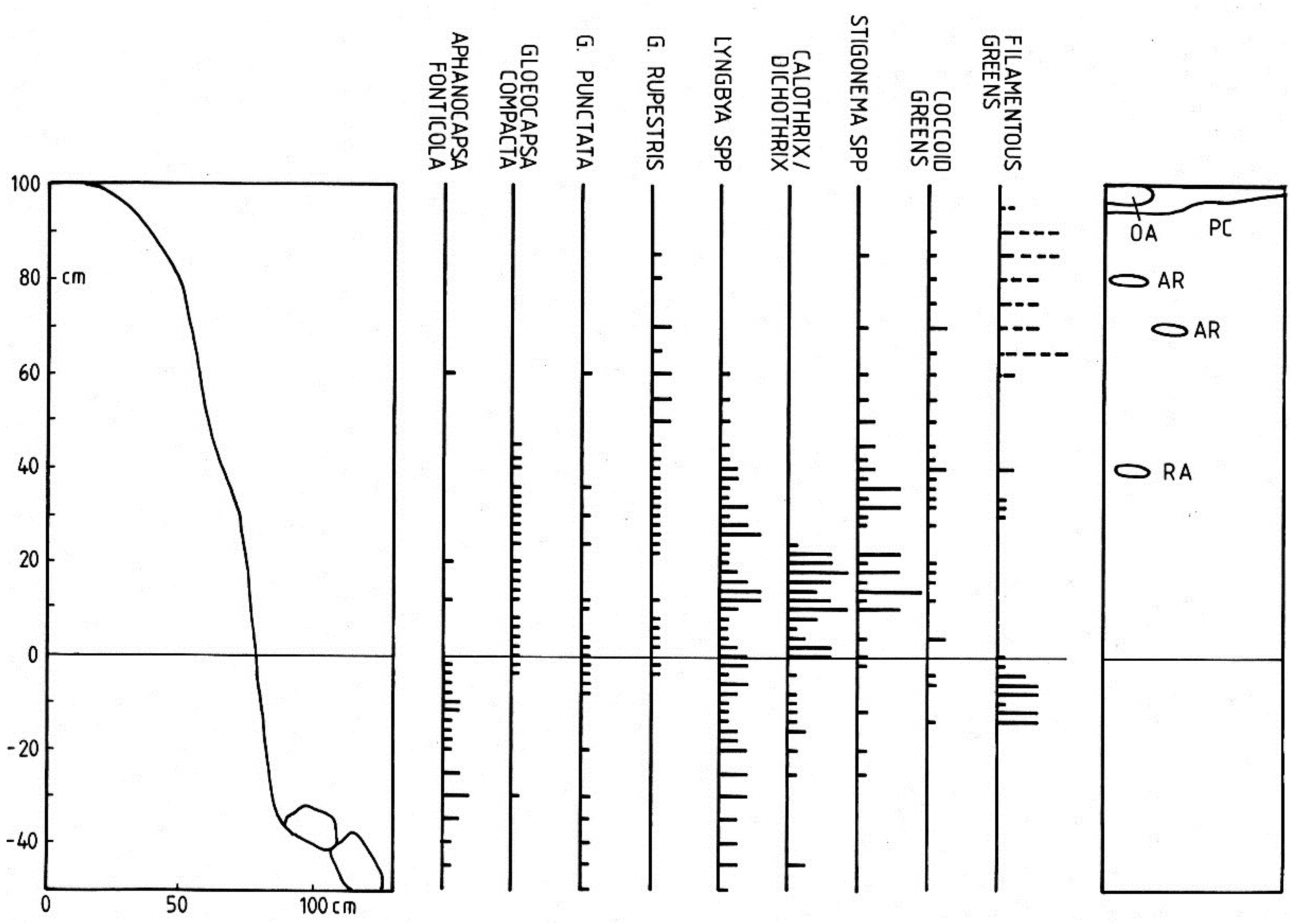
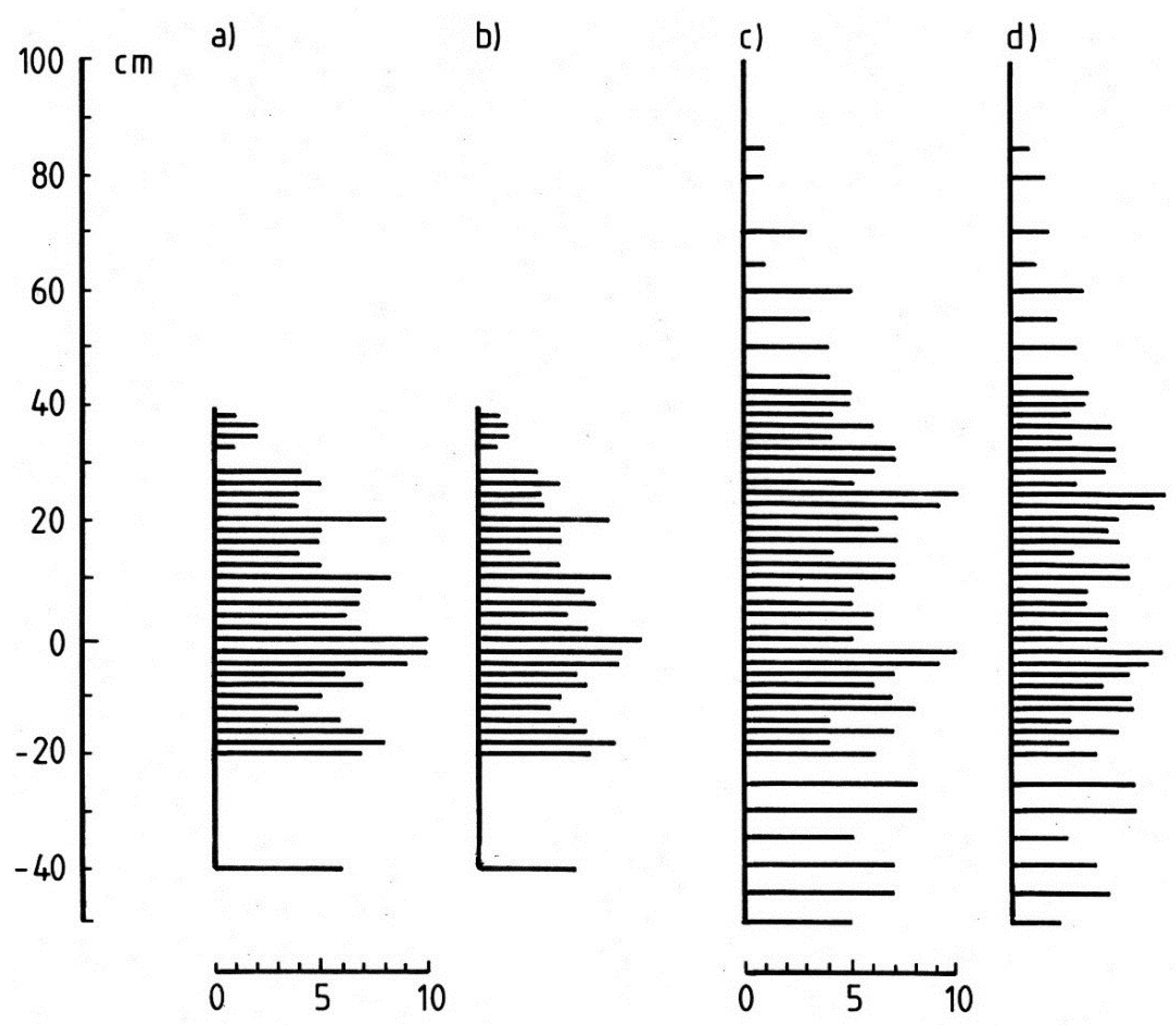
3.1.2. Vertical Zonation



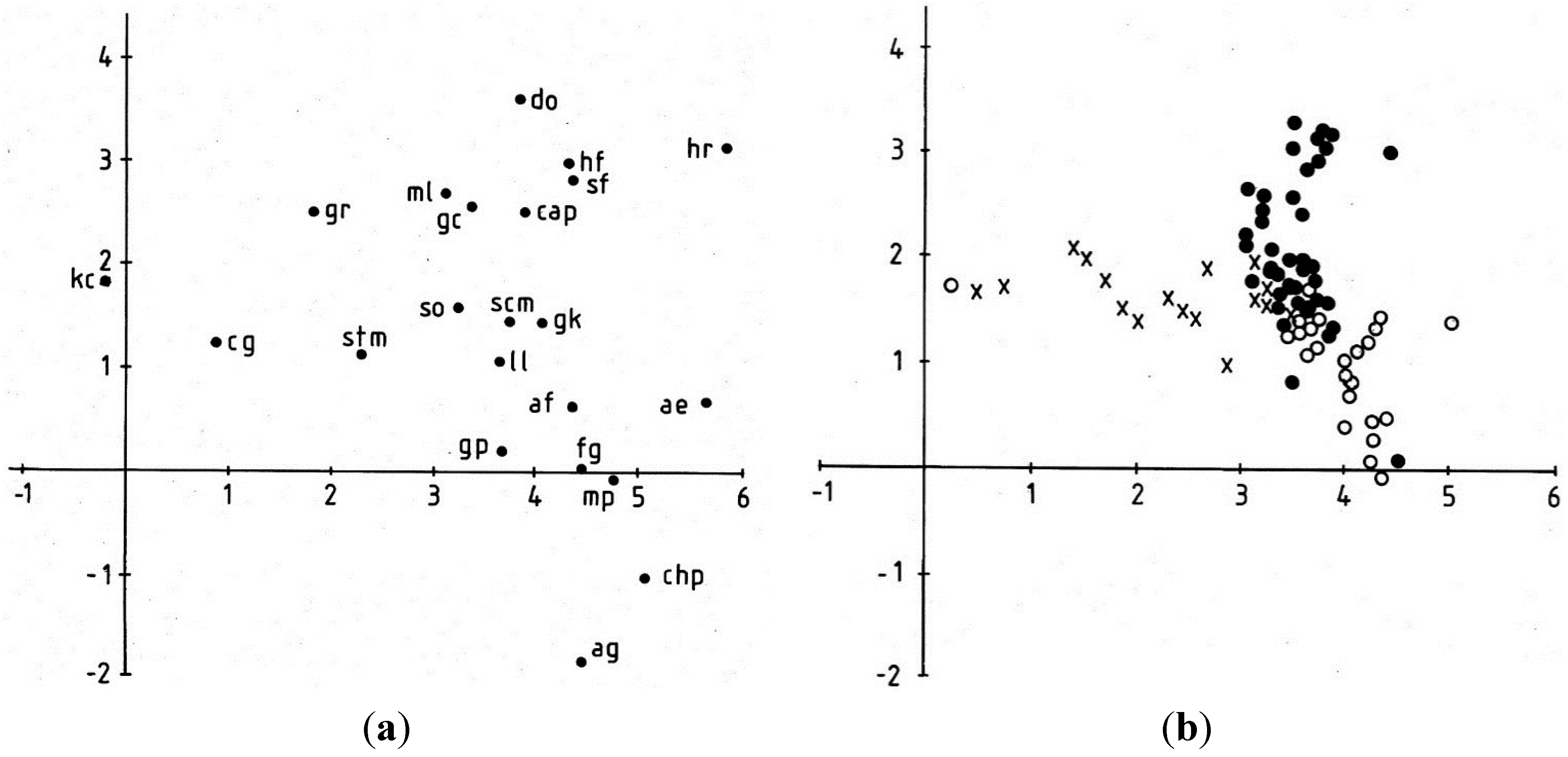
3.1.3. Cyanobacterium Communities

3.1.4. Lateral Variation


3.2. Discussion
Acknowledgments
Conflicts of Interest
References
- Godward, M. An ecological and taxonomic investigation of the littoral algal flora of Lake Windermere. J. Ecol. 1937, 25, 496–568. [Google Scholar]
- Lohscheider, J.N.; Strittmatler, M.; Kuepper, H.; Adamska, I. Vertical distribution of epibenthic freshwater cyanobacterial Synechococcus strains depends on their ability for photoprotection. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Vinebrook, R.D.; Leavitt, P.R. Different responses of littoral communities to ultraviolet radiation in an alpine lake. Ecology 1999, 80, 223–237. [Google Scholar]
- Kann, E. The littoral algal biocoenoses of Lake Erken and its outlet (Uppland, Sweden). Arch. Hydrobiol. Suppl. 1993, 97, 91–112. [Google Scholar]
- Sorokovikova, E.G.; Belykh, O.J.; Gladikh, A.S.; Kotsar, O.V.; Tikhonova, I.V.; Timoshin, O.A.; Parfenova, U.V. Diversity of cyanobacterial species and phylotypes in biofilms from the littoral zone of Lake Baikal. J. Microbiol. 2013, 51, 757–765. [Google Scholar]
- Rott, E.; Pernegger, L. Epilithic cyanophytes from soft-water mountain lakes in the Central Alps (Austria) and in the Pirin Mountains (Bulgaria) with special reference to taxonomy and vertical zonation. Arch. Hydrobiol. Suppl. 1994, 105, 249–264. [Google Scholar]
- Soilscapes. Available online: http://www.landis.org.uk/soilscapes (accessed on 10 December 2014).
- Haworth, E.Y.; de Boer, G.; Evans, I.; Osmaston, H.A.; Pennington, W.; Smith, A.; Storey, P.; Ware, B. Tarns of the Central Lake District; Brathay Exploration Group Trust: Ambleside, UK, 2003. [Google Scholar]
- Talling, J.F.; Parker, J.E. Seasonal dynamics of phytoplankton and phytobenthos and associated chemical interactions in a shallow upland marl lake (Malham Tarn, Northern England). Hydrobiologia 2002, 487, 167–181. [Google Scholar]
- Whitton, B.A. Cyanobacteria. In The Freshwater Algal Flora of the British Isles, 2nd ed.; John, D.M, Whitton, B.A., Brook, A.J., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 31–158. [Google Scholar]
- Ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and Software for Canonical Community Ordination (Version 4); Centre for Biometry: Wageningen, The Netherlands, 1998; p. 251. [Google Scholar]
- Kann, E. Die eulitorale Algenzone in Traunsee (Oberösterreich). Arch. Hydrobiol. 1958, 55, 129–192. [Google Scholar]
- Kann, E. The littoral algal vegetation of the Zellersee (Salzburg, Austria). Arch. Hydrobiol. 1982, 94, 492–501. [Google Scholar]
- Čado, I. Littoral zone of Lake Ohrid and its microphytic vegetation. Verh. Int. Ver. Theor. Angew. Limnol. 1978, 20, 1085–1089. [Google Scholar]
- Pentecost, A.; Whitton, B.A. Subaerial Cyanobacteria. In The Ecology of Cyanobacteria II; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 291–316. [Google Scholar]
- Dell’Uomo, A.; Pedrotti, F. Phytobenthos of the eulittoral zone of Lake Tovel Trentino, northern Italy. Crypt. Algol. 1982, 3, 129–146. [Google Scholar]
- Golubić, S. Die litorale Algenvegetation des Titisees. Verh. Int. Ver. Theor. Angew. Limnol. 1967, 33, 172–205. [Google Scholar]
- Fogg, G.E.; Stewart, W.D.P.; Fay, P.; Walsby, A.E. The Blue-Green Algae; Academic Press: New York, NY, USA, 1973. [Google Scholar]
- Brehm, V.; Ruttner, F. Die Biocoenosen der Lunzer Gewasser. Internationale Revue der gesamten Hydrobiologie und Hydrographie 1926, 16, 281–331. (In German) [Google Scholar]
- Hurter, E. Beobachtungen am Litoralalgen des Vierwaldstattersees. Mitteilung der Naturforschenden Gesellschaft Luzern 1928, 10, 145–398. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pentecost, A. Distribution and Ecology of Cyanobacteria in the Rocky Littoral of an English Lake District Water Body, Devoke Water. Life 2014, 4, 1026-1037. https://doi.org/10.3390/life4041026
Pentecost A. Distribution and Ecology of Cyanobacteria in the Rocky Littoral of an English Lake District Water Body, Devoke Water. Life. 2014; 4(4):1026-1037. https://doi.org/10.3390/life4041026
Chicago/Turabian StylePentecost, Allan. 2014. "Distribution and Ecology of Cyanobacteria in the Rocky Littoral of an English Lake District Water Body, Devoke Water" Life 4, no. 4: 1026-1037. https://doi.org/10.3390/life4041026