
Antimicrobial Peptides in Infectious Diseases and Beyond—A Narrative Review

Abstract
:1. Introduction
2. Search Strategy
3. Definitions and Origins of AMPs
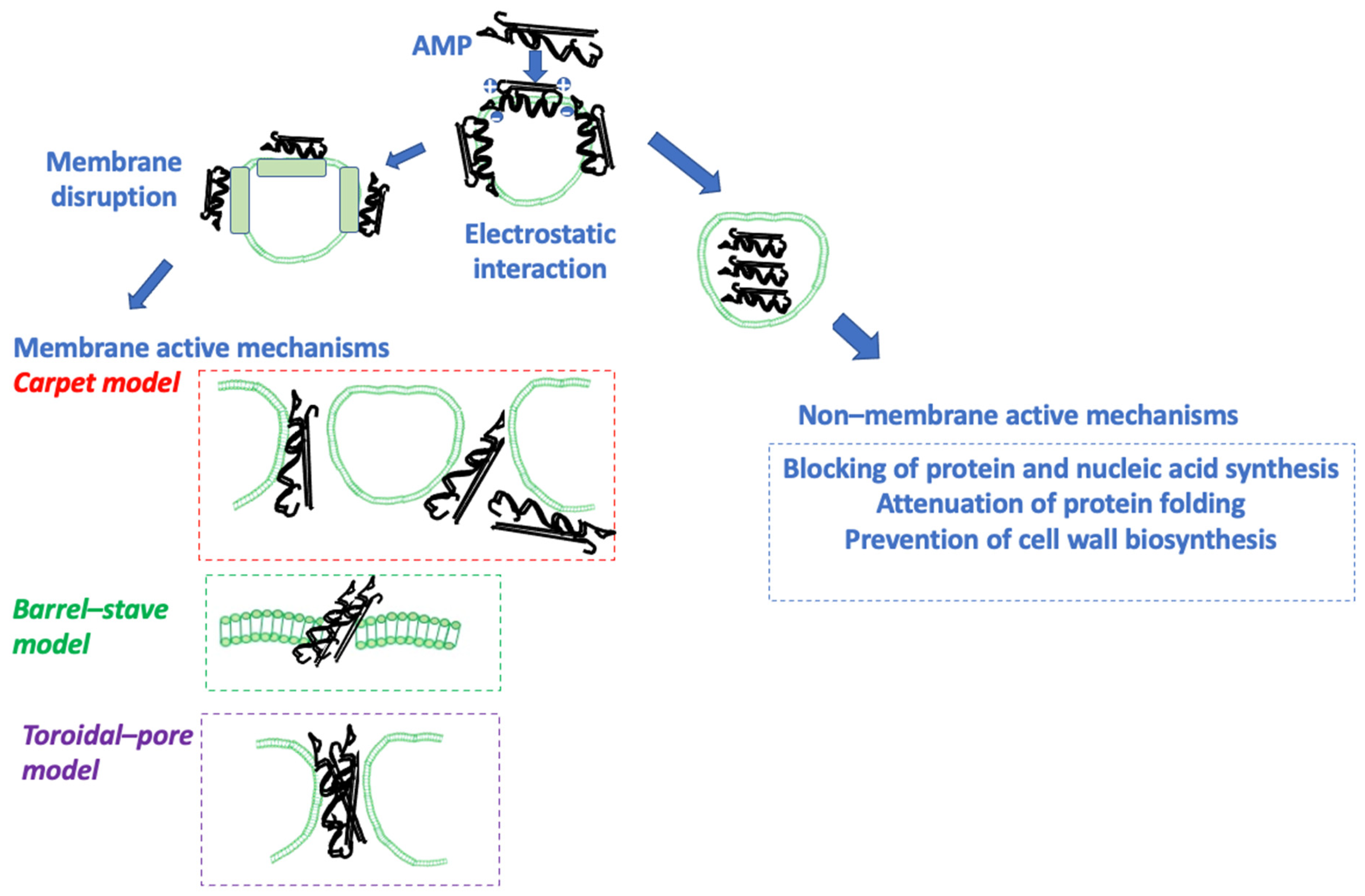
4. Structure and Function of AMPs
5. The Physiological Role of AMPs
6. Applications of AMPs
6.1. Food Industry
6.2. Animal Husbandry
6.3. Plant Protection
6.4. Aquaculture
7. Potential for the Utilization of AMPs in Infectious Diseases
8. Examples of AMPs with Potential Interest in Infectious Diseases
9. Comparison of AMPs with Classic Antibiotics
10. Combination of AMPs with Classic Antibiotics or Other Molecules
11. Examples of AMP Use in Clinical Practice
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Antimicrobial Resistance Collaborators Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Mestrovic, T.; Aguilar, G.R.; Swetschinski, L.R.; Ikuta, K.S.; Gray, A.P.; Weaver, N.D.; Han, C.; E Wool, E.; Hayoon, A.G.; Hay, S.I.; et al. The Burden of Bacterial Antimicrobial Resistance in the WHO European Region in 2019: A Cross-Country Systematic Analysis. Lancet Public Health 2022, 7, e897–e913. [Google Scholar] [CrossRef] [PubMed]
- Tarín-Pelló, A.; Suay-García, B.; Pérez-Gracia, M.-T. Antibiotic Resistant Bacteria: Current Situation and Treatment Options to Accelerate the Development of a New Antimicrobial Arsenal. Expert Rev. Anti Infect. Ther. 2022, 20, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- EClinicalMedicine. Antimicrobial Resistance: A Top Ten Global Public Health Threat. EClinicalMedicine 2021, 41, 101221. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019. [Google Scholar]
- Colomb-Cotinat, M.; Lacoste, J.; Brun-Buisson, C.; Jarlier, V.; Coignard, B.; Vaux, S. Estimating the Morbidity and Mortality Associated with Infections Due to Multidrug-Resistant Bacteria (MDRB), France, 2012. Antimicrob. Resist. Infect. Control 2016, 5, 56. [Google Scholar] [CrossRef] [Green Version]
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [Green Version]
- Pulingam, T.; Parumasivam, T.; Gazzali, A.M.; Sulaiman, A.M.; Chee, J.Y.; Lakshmanan, M.; Chin, C.F.; Sudesh, K. Antimicrobial Resistance: Prevalence, Economic Burden, Mechanisms of Resistance and Strategies to Overcome. Eur. J. Pharm. Sci. 2022, 170, 106103. [Google Scholar] [CrossRef]
- Kaier, K.; Wilson, C.; Chalkley, M.; Davey, P.G.; Suetens, C.; Grundmann, H.; de Kraker, M.; Schumacher, M.; Wolkewitz, M.; Frank, U. Health and Economic Impacts of Antibiotic Resistance in European Hospitals—Outlook on the BURDEN Project. Infection 2008, 36, 492–494. [Google Scholar] [CrossRef]
- Naylor, N.R.; Atun, R.; Zhu, N.; Kulasabanathan, K.; Silva, S.; Chatterjee, A.; Knight, G.M.; Robotham, J.V. Estimating the Burden of Antimicrobial Resistance: A Systematic Literature Review. Antimicrob. Resist. Infect. Control 2018, 7, 58. [Google Scholar] [CrossRef] [Green Version]
- Chokshi, A.; Sifri, Z.; Cennimo, D.; Horng, H. Global Contributors to Antibiotic Resistance. J. Glob. Infect. Dis. 2019, 11, 36–42. [Google Scholar] [CrossRef]
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Yang, L.; Huang, Y.; Jia, J.; Dou, T. Antibiotic Resistance Genes in Bacteria: Occurrence, Spread, and Control. J. Basic Microbiol. 2021, 61, 1049–1070. [Google Scholar] [CrossRef] [PubMed]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal Transfer of Antibiotic Resistance Genes in Clinical Environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular Mechanisms of Antibiotic Resistance Revisited. Nat. Rev. Microbiol. 2022, 21, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Vandenbroucke-Grauls, C.M.J.E.; Kluytmans, J.A.J.W. Tracing the Origins of Antibiotic Resistance. Nat. Med. 2022, 28, 638–640. [Google Scholar] [CrossRef]
- Reygaert, W.C. An Overview of the Antimicrobial Resistance Mechanisms of Bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef]
- Sandoval-Motta, S.; Aldana, M. Adaptive Resistance to Antibiotics in Bacteria: A Systems Biology Perspective. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Blanquart, F.; Lehtinen, S.; Lipsitch, M.; Fraser, C. The Evolution of Antibiotic Resistance in a Structured Host Population. J. R. Soc. Interface 2018, 15, 20180040. [Google Scholar] [CrossRef] [Green Version]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.R.M.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K.; et al. Antibiotic Resistance in Microbes: History, Mechanisms, Therapeutic Strategies and Future Prospects. J. Infect. Public. Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial Resistance in Bacteria: Mechanisms, Evolution, and Persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Flach, C.-F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Karkman, A.; Do, T.T.; Walsh, F.; Virta, M.P.J. Antibiotic-Resistance Genes in Waste Water. Trends Microbiol. 2018, 26, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazda, M.; Kumirska, J.; Stepnowski, P.; Mulkiewicz, E. Antibiotic Resistance Genes Identified in Wastewater Treatment Plant Systems—A Review. Sci. Total Environ. 2019, 697, 134023. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Yuan, T.; Zhou, L.; Cheng, S.; Qu, X.; Lu, P.; Feng, Q. Impact Factors of the Accumulation, Migration and Spread of Antibiotic Resistance in the Environment. Environ. Geochem. Health 2021, 43, 1741–1758. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Xia, Y.; Wei, W.; Ni, B.-J. Accelerated Spread of Antibiotic Resistance Genes (ARGs) Induced by Non-Antibiotic Conditions: Roles and Mechanisms. Water Res. 2022, 224, 119060. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental Factors Influencing the Development and Spread of Antibiotic Resistance. FEMS Microbiol. Rev. 2018, 42, fux053. [Google Scholar] [CrossRef]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Paul, M.; Carrara, E.; Retamar, P.; Tängdén, T.; Bitterman, R.; Bonomo, R.A.; de Waele, J.; Daikos, G.L.; Akova, M.; Harbarth, S.; et al. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) Guidelines for the Treatment of Infections Caused by Multidrug-Resistant Gram-Negative Bacilli (Endorsed by European Society of Intensive Care Medicine). Clin. Microbiol. Infect. 2022, 28, 521–547. [Google Scholar] [CrossRef]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America Guidance on the Treatment of Extended-Spectrum β-Lactamase Producing Enterobacterales (ESBL-E), Carbapenem-Resistant Enterobacterales (CRE), and Pseudomonas Aeruginosa with Difficult-to-Treat Resistance (DTR-P. Aeruginosa). Clin. Infect. Dis. 2021, 72, e169–e183. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakonstantis, S.; Ioannou, P.; Samonis, G.; Kofteridis, D.P. Systematic Review of Antimicrobial Combination Options for Pandrug-Resistant Acinetobacter Baumannii. Antibiotics 2021, 10, 1344. [Google Scholar] [CrossRef] [PubMed]
- Karakonstantis, S.; Kritsotakis, E.I.; Gikas, A. Treatment Options for K. Pneumoniae, P. Aeruginosa and A. Baumannii Co-Resistant to Carbapenems, Aminoglycosides, Polymyxins and Tigecycline: An Approach Based on the Mechanisms of Resistance to Carbapenems. Infection 2020, 48, 835–851. [Google Scholar] [CrossRef] [PubMed]
- Karakonstantis, S.; Gikas, A.; Astrinaki, E.; Kritsotakis, E.I. Excess Mortality Due to Pandrug-Resistant Acinetobacter Baumannii Infections in Hospitalized Patients. J. Hosp. Infect. 2020, 106, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Theuretzbacher, U.; Outterson, K.; Engel, A.; Karlén, A. The Global Preclinical Antibacterial Pipeline. Nat. Rev. Microbiol. 2020, 18, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Butler, M.S.; Gigante, V.; Sati, H.; Paulin, S.; Al-Sulaiman, L.; Rex, J.H.; Fernandes, P.; Arias, C.A.; Paul, M.; Thwaites, G.E.; et al. Analysis of the Clinical Pipeline of Treatments for Drug-Resistant Bacterial Infections: Despite Progress, More Action Is Needed. Antimicrob. Agents Chemother. 2022, 66, e0199121. [Google Scholar] [CrossRef]
- Malik, S.; Kaminski, M.; Landman, D.; Quale, J. Cefiderocol Resistance in Acinetobacter Baumannii: Roles of β-Lactamases, Siderophore Receptors, and Penicillin Binding Protein 3. Antimicrob. Agents Chemother. 2020, 64, e01221-20. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Wang, R.; Cai, Y. Resistance to Ceftazidime-Avibactam and Underlying Mechanisms. J. Glob. Antimicrob. Resist. 2020, 22, 18–27. [Google Scholar] [CrossRef]
- Gaibani, P.; Giani, T.; Bovo, F.; Lombardo, D.; Amadesi, S.; Lazzarotto, T.; Coppi, M.; Rossolini, G.M.; Ambretti, S. Resistance to Ceftazidime/Avibactam, Meropenem/Vaborbactam and Imipenem/Relebactam in Gram-Negative MDR Bacilli: Molecular Mechanisms and Susceptibility Testing. Antibiotics 2022, 11, 628. [Google Scholar] [CrossRef]
- Mitchell, B.G.; Fasugba, O.; Russo, P.L. Where Is the Strength of Evidence? A Review of Infection Prevention and Control Guidelines. J. Hosp. Infect. 2020, 105, 242–251. [Google Scholar] [CrossRef]
- Masterton, R.G.; Mifsud, A.J.; Rao, G.G. Hospital Isolation Precautions Working Group Review of Hospital Isolation and Infection Control Precautions. J. Hosp. Infect. 2003, 54, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Antimicrobial Stewardship and Antimicrobial Resistance. Med. Clin. N. Am. 2018, 102, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Lanckohr, C.; Bracht, H. Antimicrobial Stewardship. Curr. Opin. Crit. Care 2022, 28, 551–556. [Google Scholar] [CrossRef]
- Dyar, O.J.; Huttner, B.; Schouten, J.; Pulcini, C. ESGAP (ESCMID Study Group for Antimicrobial stewardshiP) What Is Antimicrobial Stewardship? Clin. Microbiol. Infect. 2017, 23, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered Bacteriophages for Treatment of a Patient with a Disseminated Drug-Resistant Mycobacterium Abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C. Phage Therapy’s Latest Makeover. Nat. Biotechnol. 2019, 37, 581–586. [Google Scholar] [CrossRef]
- Leuck, A.-M.; Ahmad, A.; Dunny, G.M. Combination Bacteriophage and Antibiotic Treatment for In Vitro Enterococcal Biofilms on Left Ventricular Assist Device Drivelines. Open Forum Infect. Dis. 2016, 3, 2048. [Google Scholar] [CrossRef] [Green Version]
- Ioannou, P.; Baliou, S.; Samonis, G. Bacteriophages in Infectious Diseases and Beyond—A Narrative Review. Antibiotics 2023, 12, 1012. [Google Scholar] [CrossRef]
- Hazam, P.K.; Goyal, R.; Ramakrishnan, V. Peptide Based Antimicrobials: Design Strategies and Therapeutic Potential. Prog. Biophys. Mol. Biol. 2019, 142, 10–22. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial Peptides: Pore Formers or Metabolic Inhibitors in Bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Gueguen, Y.; Bernard, R.; Julie, F.; Paulina, S.; Delphine, D.-G.; Franck, V.; Philippe, B.; Evelyne, B. Oyster Hemocytes Express a Proline-Rich Peptide Displaying Synergistic Antimicrobial Activity with a Defensin. Mol. Immunol. 2009, 46, 516–522. [Google Scholar] [CrossRef] [Green Version]
- Lemaitre, B.; Nicolas, E.; Michaut, L.; Reichhart, J.M.; Hoffmann, J.A. The Dorsoventral Regulatory Gene Cassette Spätzle/Toll/Cactus Controls the Potent Antifungal Response in Drosophila Adults. Cell 1996, 86, 973–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghadam, M.T.; Mojtahedi, A.; Moghaddam, M.M.; Fasihi-Ramandi, M.; Mirnejad, R. Rescuing Humanity by Antimicrobial Peptides against Colistin-Resistant Bacteria. Appl. Microbiol. Biotechnol. 2022, 106, 3879–3893. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Sahl, H.-G. Antimicrobial and Host-Defense Peptides as New Anti-Infective Therapeutic Strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial Peptides as Therapeutic Agents: Opportunities and Challenges. Crit. Rev. Biotechnol. 2020, 40, 978–992. [Google Scholar] [CrossRef]
- Giovati, L.; Ciociola, T.; Magliani, W.; Conti, S. Antimicrobial Peptides with Antiprotozoal Activity: Current State and Future Perspectives. Future Med. Chem. 2018, 10, 2569–2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brice, D.C.; Diamond, G. Antiviral Activities of Human Host Defense Peptides. Curr. Med. Chem. 2020, 27, 1420–1443. [Google Scholar] [CrossRef]
- Fernández de Ullivarri, M.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal Peptides as Therapeutic Agents. Front. Cell Infect. Microbiol. 2020, 10, 105. [Google Scholar] [CrossRef]
- Miyoshi, N.; Isogai, E.; Hiramatsu, K.; Sasaki, T. Activity of Tick Antimicrobial Peptide from Ixodes Persulcatus (Persulcatusin) against Cell Membranes of Drug-Resistant Staphylococcus Aureus. J. Antibiot. 2017, 70, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, N.; Saito, T.; Ohmura, T.; Kuroda, K.; Suita, K.; Ihara, K.; Isogai, E. Functional Structure and Antimicrobial Activity of Persulcatusin, an Antimicrobial Peptide from the Hard Tick Ixodes Persulcatus. Parasites Vectors 2016, 9, 85. [Google Scholar] [CrossRef] [Green Version]
- Landman, D.; Georgescu, C.; Martin, D.A.; Quale, J. Polymyxins Revisited. Clin. Microbiol. Rev. 2008, 21, 449–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velkov, T.; Thompson, P.E.; Nation, R.L.; Li, J. Structure—Activity Relationships of Polymyxin Antibiotics. J. Med. Chem. 2010, 53, 1898–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Kasiakou, S.K. Colistin: The Revival of Polymyxins for the Management of Multidrug-Resistant Gram-Negative Bacterial Infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, A.T.Y.; Gellatly, S.L.; Hancock, R.E.W. Multifunctional Cationic Host Defence Peptides and Their Clinical Applications. Cell Mol. Life Sci. 2011, 68, 2161–2176. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Nijnik, A.; Philpott, D.J. Modulating Immunity as a Therapy for Bacterial Infections. Nat. Rev. Microbiol. 2012, 10, 243–254. [Google Scholar] [CrossRef]
- Chen, C.H.; Lu, T.K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial Peptides: Key Components of the Innate Immune System. Crit. Rev. Biotechnol. 2012, 32, 143–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The Antimicrobial Peptides and Their Potential Clinical Applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.-Y. Antimicrobial Peptides: New Hope in the War against Multidrug Resistance. Zool. Res. 2019, 40, 488–505. [Google Scholar] [CrossRef]
- Chung, P.Y.; Khanum, R. Antimicrobial Peptides as Potential Anti-Biofilm Agents against Multidrug-Resistant Bacteria. J. Microbiol. Immunol. Infect. 2017, 50, 405–410. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The Expanding Scope of Antimicrobial Peptide Structures and Their Modes of Action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Takahashi, D.; Shukla, S.K.; Prakash, O.; Zhang, G. Structural Determinants of Host Defense Peptides for Antimicrobial Activity and Target Cell Selectivity. Biochimie 2010, 92, 1236–1241. [Google Scholar] [CrossRef]
- Travkova, O.G.; Moehwald, H.; Brezesinski, G. The Interaction of Antimicrobial Peptides with Membranes. Adv. Colloid Interface Sci. 2017, 247, 521–532. [Google Scholar] [CrossRef]
- Priyadarshini, D.; Ivica, J.; Separovic, F.; de Planque, M.R.R. Characterisation of Cell Membrane Interaction Mechanisms of Antimicrobial Peptides by Electrical Bilayer Recording. Biophys. Chem. 2022, 281, 106721. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L. The Tall Office Building Artistically Considered. Lippincott’s Mag. 1896, 57, 403–409. [Google Scholar]
- Matsuzaki, K. Why and How Are Peptide-Lipid Interactions Utilized for Self-Defense? Magainins and Tachyplesins as Archetypes. Biochim. Biophys. Acta 1999, 1462, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dathe, M.; Wieprecht, T. Structural Features of Helical Antimicrobial Peptides: Their Potential to Modulate Activity on Model Membranes and Biological Cells. Biochim. Biophys. Acta 1999, 1462, 71–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilhelmelli, F.; Vilela, N.; Albuquerque, P.; da Derengowski, L.S.; Silva-Pereira, I.; Kyaw, C.M. Antibiotic Development Challenges: The Various Mechanisms of Action of Antimicrobial Peptides and of Bacterial Resistance. Front. Microbiol. 2013, 4, 353. [Google Scholar] [CrossRef] [Green Version]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of Antimicrobial Peptide Action and Resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [Green Version]
- Arias, M.; Piga, K.B.; Hyndman, M.E.; Vogel, H.J. Improving the Activity of Trp-Rich Antimicrobial Peptides by Arg/Lys Substitutions and Changing the Length of Cationic Residues. Biomolecules 2018, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Bai, X.; Luan, N.; Yao, H.; Zhang, Z.; Liu, W.; Chen, Y.; Yan, X.; Rong, M.; Lai, R.; et al. A Designed Tryptophan- and Lysine/Arginine-Rich Antimicrobial Peptide with Therapeutic Potential for Clinical Antibiotic-Resistant Candida Albicans Vaginitis. J. Med. Chem. 2016, 59, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.-C.; Strandberg, E.; Grau-Campistany, A.; Wadhwani, P.; Reichert, J.; Bürck, J.; Rabanal, F.; Auger, M.; Paquin, J.-F.; Ulrich, A.S. Influence of the Length and Charge on the Activity of α-Helical Amphipathic Antimicrobial Peptides. Biochemistry 2017, 56, 1680–1695. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Park, T.G.; Lee, K.H. The Effect of Charge Increase on the Specificity and Activity of a Short Antimicrobial Peptide. Peptides 2001, 22, 1669–1674. [Google Scholar] [CrossRef]
- Lyu, Y.; Yang, Y.; Lyu, X.; Dong, N.; Shan, A. Antimicrobial Activity, Improved Cell Selectivity and Mode of Action of Short PMAP-36-Derived Peptides against Bacteria and Candida. Sci. Rep. 2016, 6, 27258. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Mant, C.T.; Farmer, S.W.; Hancock, R.E.W.; Vasil, M.L.; Hodges, R.S. Rational Design of Alpha-Helical Antimicrobial Peptides with Enhanced Activities and Specificity/Therapeutic Index. J. Biol. Chem. 2005, 280, 12316–12329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Vasil, A.I.; Hale, J.D.; Hancock, R.E.W.; Vasil, M.L.; Hodges, R.S. Effects of Net Charge and the Number of Positively Charged Residues on the Biological Activity of Amphipathic Alpha-Helical Cationic Antimicrobial Peptides. Biopolymers 2008, 90, 369–383. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of Peptide Hydrophobicity in the Mechanism of Action of Alpha-Helical Antimicrobial Peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Tachi, T.; Epand, R.F.; Epand, R.M.; Matsuzaki, K. Position-Dependent Hydrophobicity of the Antimicrobial Magainin Peptide Affects the Mode of Peptide-Lipid Interactions and Selective Toxicity. Biochemistry 2002, 41, 10723–10731. [Google Scholar] [CrossRef]
- Hancock, R.E.; Lehrer, R. Cationic Peptides: A New Source of Antibiotics. Trends Biotechnol. 1998, 16, 82–88. [Google Scholar] [CrossRef]
- Ulm, H.; Wilmes, M.; Shai, Y.; Sahl, H.-G. Antimicrobial Host Defensins—Specific Antibiotic Activities and Innate Defense Modulation. Front. Immunol. 2012, 3, 249. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-Helical Cationic Antimicrobial Peptides: Relationships of Structure and Function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falanga, A.; Lombardi, L.; Franci, G.; Vitiello, M.; Iovene, M.R.; Morelli, G.; Galdiero, M.; Galdiero, S. Marine Antimicrobial Peptides: Nature Provides Templates for the Design of Novel Compounds against Pathogenic Bacteria. Int. J. Mol. Sci. 2016, 17, 785. [Google Scholar] [CrossRef] [Green Version]
- Abrunhosa, F.; Faria, S.; Gomes, P.; Tomaz, I.; Pessoa, J.C.; Andreu, D.; Bastos, M. Interaction and Lipid-Induced Conformation of Two Cecropin−Melittin Hybrid Peptides Depend on Peptide and Membrane Composition. J. Phys. Chem. B 2005, 109, 17311–17319. [Google Scholar] [CrossRef] [Green Version]
- Breukink, E.; Wiedemann, I.; van Kraaij, C.; Kuipers, O.P.; Sahl, H.-G.; de Kruijff, B. Use of the Cell Wall Precursor Lipid II by a Pore-Forming Peptide Antibiotic. Science 1999, 286, 2361–2364. [Google Scholar] [CrossRef] [Green Version]
- Shai, Y. Mode of Action of Membrane Active Antimicrobial Peptides. Biopolymers 2002, 66, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Baño, J.; del Toro, M.D.; López-Méndez, J.; Mutters, N.T.; Pascual, A. Minimum Requirements in Infection Control. Clin. Microbiol. Infect. 2015, 21, 1072–1076. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Wang, K.; Dang, W.; Chen, R.; Xie, J.; Zhang, B.; Song, J.; Wang, R. Two Hits Are Better than One: Membrane-Active and DNA Binding-Related Double-Action Mechanism of NK-18, a Novel Antimicrobial Peptide Derived from Mammalian NK-Lysin. Antimicrob. Agents Chemother. 2013, 57, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of Action of the Antimicrobial Peptide Buforin II: Buforin II Kills Microorganisms by Penetrating the Cell Membrane and Inhibiting Cellular Functions. Biochem. Biophys. Res. Commun. 1998, 244, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Hwang, J.S.; Lee, D.G. Antibacterial Action of Lactoferricin B like Peptide against Escherichia Coli: Reactive Oxygen Species-induced Apoptosis-like Death. J. Appl. Microbiol. 2020, 129, 287–295. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Rozek, A. Role of Membranes in the Activities of Antimicrobial Cationic Peptides. FEMS Microbiol. Lett. 2002, 206, 143–149. [Google Scholar] [CrossRef]
- Hilchie, A.L.; Wuerth, K.; Hancock, R.E.W. Immune Modulation by Multifaceted Cationic Host Defense (Antimicrobial) Peptides. Nat. Chem. Biol. 2013, 9, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Afacan, N.J.; Yeung, A.T.Y.; Pena, O.M.; Hancock, R.E.W. Therapeutic Potential of Host Defense Peptides in Antibiotic-Resistant Infections. Curr. Pharm. Des. 2012, 18, 807–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijnik, A.; Hancock, R. Host Defence Peptides: Antimicrobial and Immunomodulatory Activity and Potential Applications for Tackling Antibiotic-Resistant Infections. Emerg. Health Threat. J. 2009, 2, e1. [Google Scholar] [CrossRef]
- Zhang, L.; Gallo, R.L. Antimicrobial Peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Coorens, M.; Scheenstra, M.R.; Veldhuizen, E.J.A.; Haagsman, H.P. Interspecies Cathelicidin Comparison Reveals Divergence in Antimicrobial Activity, TLR Modulation, Chemokine Induction and Regulation of Phagocytosis. Sci. Rep. 2017, 7, 40874. [Google Scholar] [CrossRef] [Green Version]
- Scott, M.G.; Davidson, D.J.; Gold, M.R.; Bowdish, D.; Hancock, R.E.W. The Human Antimicrobial Peptide LL-37 Is a Multifunctional Modulator of Innate Immune Responses. J. Immunol. 2002, 169, 3883–3891. [Google Scholar] [CrossRef] [Green Version]
- Niyonsaba, F.; Iwabuchi, K.; Someya, A.; Hirata, M.; Matsuda, H.; Ogawa, H.; Nagaoka, I. A Cathelicidin Family of Human Antibacterial Peptide LL-37 Induces Mast Cell Chemotaxis. Immunology 2002, 106, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, Y.; Zhang, Y.; Lai, Y.; Chen, W.; Xiao, Z.; Zhang, W.; Jin, M.; Yu, B. LL-37-Induced Human Mast Cell Activation through G Protein-Coupled Receptor MrgX2. Int. Immunopharmacol. 2017, 49, 6–12. [Google Scholar] [CrossRef]
- Le, C.-F.; Fang, C.-M.; Sekaran, S.D. Intracellular Targeting Mechanisms by Antimicrobial Peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Song, Y. Mechanism of Antimicrobial Peptides: Antimicrobial, Anti-Inflammatory and Antibiofilm Activities. Int. J. Mol. Sci. 2021, 22, 11401. [Google Scholar] [CrossRef] [PubMed]
- Rahnamaeian, M.; Cytryńska, M.; Zdybicka-Barabas, A.; Vilcinskas, A. The Functional Interaction between Abaecin and Pore-Forming Peptides Indicates a General Mechanism of Antibacterial Potentiation. Peptides 2016, 78, 17–23. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Ludtke, S.J.; Heller, W.T.; Huang, H.W. Mechanism of Alamethicin Insertion into Lipid Bilayers. Biophys. J. 1996, 71, 2669–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.-F.; Ma, G.-X.; Zhou, X.-X. Apidaecin-Type Peptides: Biodiversity, Structure-Function Relationships and Mode of Action. Peptides 2006, 27, 2350–2359. [Google Scholar] [CrossRef]
- Boman, H.G.; Agerberth, B.; Boman, A. Mechanisms of Action on Escherichia Coli of Cecropin P1 and PR-39, Two Antibacterial Peptides from Pig Intestine. Infect. Immun. 1993, 61, 2978–2984. [Google Scholar] [CrossRef] [Green Version]
- Cudic, M.; Bulet, P.; Hoffmann, R.; Craik, D.J.; Otvos, L. Chemical Synthesis, Antibacterial Activity and Conformation of Diptericin, an 82-Mer Peptide Originally Isolated from Insects. Eur. J. Biochem. 1999, 266, 549–558. [Google Scholar] [CrossRef]
- Chileveru, H.R.; Lim, S.A.; Chairatana, P.; Wommack, A.J.; Chiang, I.-L.; Nolan, E.M. Visualizing Attack of Escherichia Coli by the Antimicrobial Peptide Human Defensin 5. Biochemistry 2015, 54, 1767–1777. [Google Scholar] [CrossRef] [Green Version]
- Puri, S.; Edgerton, M. How Does It Kill?: Understanding the Candidacidal Mechanism of Salivary Histatin 5. Eukaryot. Cell 2014, 13, 958–964. [Google Scholar] [CrossRef] [Green Version]
- Yoneyama, F.; Imura, Y.; Ichimasa, S.; Fujita, K.; Zendo, T.; Nakayama, J.; Matsuzaki, K.; Sonomoto, K. Lacticin Q, a Lactococcal Bacteriocin, Causes High-Level Membrane Permeability in the Absence of Specific Receptors. Appl. Environ. Microbiol. 2009, 75, 538–541. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Chamorro, L.; Puertollano, M.A.; Puertollano, E.; de Cienfuegos, G.A.; de Pablo, M.A. In Vitro Biological Activities of Magainin Alone or in Combination with Nisin. Peptides 2006, 27, 1201–1209. [Google Scholar] [CrossRef]
- Piers, K.L.; Brown, M.H.; Hancock, R.E. Improvement of Outer Membrane-Permeabilizing and Lipopolysaccharide-Binding Activities of an Antimicrobial Cationic Peptide by C-Terminal Modification. Antimicrob. Agents Chemother. 1994, 38, 2311–2316. [Google Scholar] [CrossRef] [Green Version]
- Ruhr, E.; Sahl, H.G. Mode of Action of the Peptide Antibiotic Nisin and Influence on the Membrane Potential of Whole Cells and on Cytoplasmic and Artificial Membrane Vesicles. Antimicrob. Agents Chemother. 1985, 27, 841–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiarto, H.; Yu, P.-L. Mechanisms of Action of Ostrich Beta-Defensins against Escherichia Coli. FEMS Microbiol. Lett. 2007, 270, 195–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsu, T.; Nakao, S.; Iwanaga, S. Mode of Action of an Antimicrobial Peptide, Tachyplesin I, on Biomembranes. Biol. Pharm. Bull. 1993, 16, 178–181. [Google Scholar] [CrossRef] [Green Version]
- Dash, R.; Bhattacharjya, S. Thanatin: An Emerging Host Defense Antimicrobial Peptide with Multiple Modes of Action. Int. J. Mol. Sci. 2021, 22, 1522. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.P.; Yeaman, M.R.; Nast, C.C.; Bayer, A.S. The Cytoplasmic Membrane Is a Primary Target for the Staphylocidal Action of Thrombin-Induced Platelet Microbicidal Protein. Infect. Immun. 1997, 65, 4795–4800. [Google Scholar] [CrossRef]
- Ganz, T. The Role of Antimicrobial Peptides in Innate Immunity. Integr. Comp. Biol. 2003, 43, 300–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harder, J.; Schröder, J.-M.; Gläser, R. The Skin Surface as Antimicrobial Barrier: Present Concepts and Future Outlooks. Exp. Dermatol. 2013, 22, 1–5. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Ma, Y.; Wang, X.; Liang, J.; Zhang, C.; Zhang, K.; Lin, G.; Lai, R. The First Antimicrobial Peptide from Sea Amphibian. Mol. Immunol. 2008, 45, 678–681. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, X.; Wang, J. Recent Advances in Antibacterial and Antiendotoxic Peptides or Proteins from Marine Resources. Mar. Drugs 2018, 16, 57. [Google Scholar] [CrossRef] [Green Version]
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of Antimicrobial Peptides. A Review. Mol. Biol. Rep. 2012, 39, 10957–10970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treffers, C.; Chen, L.; Anderson, R.C.; Yu, P.-L. Isolation and Characterisation of Antimicrobial Peptides from Deer Neutrophils. Int. J. Antimicrob. Agents 2005, 26, 165–169. [Google Scholar] [CrossRef]
- Brogden, K.A.; Ackermann, M.; McCray, P.B.; Tack, B.F. Antimicrobial Peptides in Animals and Their Role in Host Defences. Int. J. Antimicrob. Agents 2003, 22, 465–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zou, A.; Manchu, R.; Zhou, Y.; Wang, S. Purification and Antimicrobial Activity of Antimicrobial Protein from Brown-Spotted Grouper, Epinephelus Fario. Zool. Res. 2009, 29, 627–632. [Google Scholar] [CrossRef]
- Haney, E.F.; Hancock, R.E.W. Peptide Design for Antimicrobial and Immunomodulatory Applications. Biopolymers 2013, 100, 572–583. [Google Scholar] [CrossRef] [Green Version]
- Maróti, G.; Kereszt, A.; Kondorosi, E.; Mergaert, P. Natural Roles of Antimicrobial Peptides in Microbes, Plants and Animals. Res. Microbiol. 2011, 162, 363–374. [Google Scholar] [CrossRef]
- Oppenheim, J.J.; Biragyn, A.; Kwak, L.W.; Yang, D. Roles of Antimicrobial Peptides Such as Defensins in Innate and Adaptive Immunity. Ann. Rheum. Dis. 2003, 62 (Suppl. S2), ii17–ii21. [Google Scholar] [CrossRef]
- Qi, R.-H.; Chen, Y.; Guo, Z.-L.; Zhang, F.; Fang, Z.; Huang, K.; Yu, H.-N.; Wang, Y.-P. Identification and Characterization of Two Novel Cathelicidins from the Frog Odorrana Livida. Zool. Res. 2019, 40, 94–101. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. A Peptidomics Study Reveals the Impressive Antimicrobial Peptide Arsenal of the Wax Moth Galleria Mellonella. Insect Biochem. Mol. Biol. 2009, 39, 792–800. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Brown, K.L.; Mookherjee, N. Host Defence Peptides from Invertebrates—Emerging Antimicrobial Strategies. Immunobiology 2006, 211, 315–322. [Google Scholar] [CrossRef]
- Bulet, P.; Stöcklin, R. Insect Antimicrobial Peptides: Structures, Properties and Gene Regulation. Protein Pept. Lett. 2005, 12, 3–11. [Google Scholar] [CrossRef]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial Peptides from Plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef] [Green Version]
- Stotz, H.U.; Thomson, J.G.; Wang, Y. Plant Defensins: Defense, Development and Application. Plant Signal Behav. 2009, 4, 1010–1012. [Google Scholar] [CrossRef] [Green Version]
- Hammami, R.; Ben Hamida, J.; Vergoten, G.; Fliss, I. PhytAMP: A Database Dedicated to Antimicrobial Plant Peptides. Nucleic Acids Res. 2009, 37, D963–D968. [Google Scholar] [CrossRef] [Green Version]
- Nawrot, R.; Barylski, J.; Nowicki, G.; Broniarczyk, J.; Buchwald, W.; Goździcka-Józefiak, A. Plant Antimicrobial Peptides. Folia Microbiol 2014, 59, 181–196. [Google Scholar] [CrossRef] [Green Version]
- Mylonakis, E.; Podsiadlowski, L.; Muhammed, M.; Vilcinskas, A. Diversity, Evolution and Medical Applications of Insect Antimicrobial Peptides. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozgonyi, F.; Szabo, D.; Kocsis, B.; Ostorházi, E.; Abbadessa, G.; Cassone, M.; Wade, J.D.; Otvos, L. The Antibacterial Effect of a Proline-Rich Antibacterial Peptide A3-APO. Curr. Med. Chem. 2009, 16, 3996–4002. [Google Scholar] [CrossRef]
- Ciumac, D.; Gong, H.; Hu, X.; Lu, J.R. Membrane targeting cationic antimicrobial peptides. J. Colloid. Interface Sci. 2019, 537, 163–185. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Patočka, J.; Kuča, K. Insect Antimicrobial Peptides, a Mini Review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-C.; Lin, C.-H.; Sung, C.T.; Fang, J.-Y. Antibacterial Activities of Bacteriocins: Application in Foods and Pharmaceuticals. Front. Microbiol. 2014, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Mills, S.; Serrano, L.M.; Griffin, C.; O’Connor, P.M.; Schaad, G.; Bruining, C.; Hill, C.; Ross, R.P.; Meijer, W.C. Inhibitory Activity of Lactobacillus Plantarum LMG P-26358 against Listeria Innocua When Used as an Adjunct Starter in the Manufacture of Cheese. Microb. Cell Fact. 2011, 10 (Suppl. S1), S7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef] [PubMed]
- Galanakis, C. Food Quality and Shelf Life; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 978-0-12-817190-5. [Google Scholar]
- Erdem Büyükkiraz, M.; Kesmen, Z. Antimicrobial Peptides (AMPs): A Promising Class of Antimicrobial Compounds. J. Appl. Microbiol. 2022, 132, 1573–1596. [Google Scholar] [CrossRef] [PubMed]
- Elsser-Gravesen, D.; Elsser-Gravesen, A. Biopreservatives. Adv. Biochem. Eng. Biotechnol. 2014, 143, 29–49. [Google Scholar] [CrossRef]
- Thouzeau, C.; Le Maho, Y.; Froget, G.; Sabatier, L.; Le Bohec, C.; Hoffmann, J.A.; Bulet, P. Spheniscins, Avian Beta-Defensins in Preserved Stomach Contents of the King Penguin, Aptenodytes Patagonicus. J. Biol. Chem. 2003, 278, 51053–51058. [Google Scholar] [CrossRef] [Green Version]
- Józefiak, D.; Kierończyk, B.; Juśkiewicz, J.; Zduńczyk, Z.; Rawski, M.; Długosz, J.; Sip, A.; Højberg, O. Dietary Nisin Modulates the Gastrointestinal Microbial Ecology and Enhances Growth Performance of the Broiler Chickens. PLoS ONE 2013, 8, e85347. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.X.; Zhang, S.F.; Wang, T.D.; Guo, X.J.; Hu, R.L. Mammary Gland Expression of Antibacterial Peptide Genes to Inhibit Bacterial Pathogens Causing Mastitis. J. Dairy Sci. 2007, 90, 5218–5225. [Google Scholar] [CrossRef] [Green Version]
- Hao, H.; Cheng, G.; Iqbal, Z.; Ai, X.; Hussain, H.I.; Huang, L.; Dai, M.; Wang, Y.; Liu, Z.; Yuan, Z. Benefits and Risks of Antimicrobial Use in Food-Producing Animals. Front. Microbiol. 2014, 5, 288. [Google Scholar] [CrossRef] [Green Version]
- Cutler, S.A.; Lonergan, S.M.; Cornick, N.; Johnson, A.K.; Stahl, C.H. Dietary Inclusion of Colicin E1 Is Effective in Preventing Postweaning Diarrhea Caused by F18-Positive Escherichia Coli in Pigs. Antimicrob. Agents Chemother. 2007, 51, 3830–3835. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Yin, Y.; Zhang, Y.; Huang, R.; Sun, Z.; Li, T.; Chu, W.; Kong, X.; Li, L.; Geng, M.; et al. Effects of Dietary Supplementation with an Expressed Fusion Peptide Bovine Lactoferricin-Lactoferrampin on Performance, Immune Function and Intestinal Mucosal Morphology in Piglets Weaned at Age 21 d. Br. J. Nutr. 2009, 101, 998–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Zhang, F.; Huang, Z.; Liu, H.; Xie, C.; Zhang, J.; Thacker, P.A.; Qiao, S. Effects of the Antimicrobial Peptide Cecropin AD on Performance and Intestinal Health in Weaned Piglets Challenged with Escherichia Coli. Peptides 2012, 35, 225–230. [Google Scholar] [CrossRef]
- Guo, N.; Zhang, B.; Hu, H.; Ye, S.; Chen, F.; Li, Z.; Chen, P.; Wang, C.; He, Q. Caerin1.1 Suppresses the Growth of Porcine Epidemic Diarrhea Virus In Vitro via Direct Binding to the Virus. Viruses 2018, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Wang, K.; She, R.; Ma, W.; Peng, F.; Jin, H. Swine Intestine Antimicrobial Peptides Inhibit Infectious Bronchitis Virus Infectivity in Chick Embryos. Poult. Sci. 2010, 89, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, X.; Lian, K.; Tian, X.; Zhang, M.; Wang, S.; Chen, C.; Nie, C.; Pan, Y.; Han, F.; et al. Antiviral Effects of Bovine Antimicrobial Peptide against TGEV in Vivo and in Vitro. J. Vet. Sci. 2020, 21, e80. [Google Scholar] [CrossRef]
- Hsieh, I.-N.; Hartshorn, K.L. The Role of Antimicrobial Peptides in Influenza Virus Infection and Their Potential as Antiviral and Immunomodulatory Therapy. Pharmaceuticals 2016, 9, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rekha; Naik, S.N.; Prasad, R. Pesticide Residue in Organic and Conventional Food-Risk Analysis. J. Chem. Health Saf. 2006, 13, 12–19. [Google Scholar] [CrossRef]
- Kamysz, W.; Krolicka, A.; Bogucka, K.; Ossowski, T.; Lukasiak, J.; Lojkowska, E. Antibacterial Activity of Synthetic Peptides Against Plant Pathogenic Pectobacterium Species. J. Phytopathol. 2005, 153, 313–317. [Google Scholar] [CrossRef]
- Gao, A.G.; Hakimi, S.M.; Mittanck, C.A.; Wu, Y.; Woerner, B.M.; Stark, D.M.; Shah, D.M.; Liang, J.; Rommens, C.M. Fungal Pathogen Protection in Potato by Expression of a Plant Defensin Peptide. Nat. Biotechnol. 2000, 18, 1307–1310. [Google Scholar] [CrossRef]
- Zakharchenko, N.S.; Rukavtsova, E.B.; Gudkov, A.T.; Bur’ianov, I.I. [Enhanced resistance to phytopathogenic bacteria in transgenic tobacco plants with synthetic gene of antimicrobial peptide cecropin P1]. Genetika 2005, 41, 1445–1452. [Google Scholar] [CrossRef]
- Alan, A.R.; Blowers, A.; Earle, E.D. Expression of a Magainin-Type Antimicrobial Peptide Gene (MSI-99) in Tomato Enhances Resistance to Bacterial Speck Disease. Plant Cell Rep. 2004, 22, 388–396. [Google Scholar] [CrossRef]
- Vidal, J.R.; Kikkert, J.R.; Malnoy, M.A.; Wallace, P.G.; Barnard, J.; Reisch, B.I. Evaluation of Transgenic “Chardonnay” (Vitis Vinifera) Containing Magainin Genes for Resistance to Crown Gall and Powdery Mildew. Transgenic Res. 2006, 15, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Ganapathi, T.R.; Mukherjee, P.K.; Bapat, V.A. MSI-99, a Magainin Analogue, Imparts Enhanced Disease Resistance in Transgenic Tobacco and Banana. Planta 2003, 216, 587–596. [Google Scholar] [CrossRef]
- Yin, Z.-X.; He, W.; Chen, W.-J.; Yan, J.-H.; Yang, J.-N.; Chan, S.-M.; He, J.-G. Cloning, Expression and Antimicrobial Activity of an Antimicrobial Peptide, Epinecidin-1, from the Orange-Spotted Grouper, Epinephelus Coioides. Aquaculture 2006, 253, 204–211. [Google Scholar] [CrossRef]
- León, R.; Ruiz, M.; Valero, Y.; Cárdenas, C.; Guzman, F.; Vila, M.; Cuesta, A. Exploring Small Cationic Peptides of Different Origin as Potential Antimicrobial Agents in Aquaculture. Fish Shellfish Immunol. 2020, 98, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Di, Y.P. Antimicrobial Peptides: A Potential Therapeutic Option for Surgical Site Infections. Clin. Surg. 2017, 2, 1740. [Google Scholar]
- Loeffler, J.M.; Nelson, D.; Fischetti, V.A. Rapid Killing of Streptococcus Pneumoniae with a Bacteriophage Cell Wall Hydrolase. Science 2001, 294, 2170–2172. [Google Scholar] [CrossRef] [PubMed]
- Van ’t Hof, W.; Veerman, E.C.; Helmerhorst, E.J.; Amerongen, A.V. Antimicrobial Peptides: Properties and Applicability. Biol. Chem. 2001, 382, 597–619. [Google Scholar] [CrossRef]
- Gee, M.L.; Burton, M.; Grevis-James, A.; Hossain, M.A.; McArthur, S.; Palombo, E.A.; Wade, J.D.; Clayton, A.H.A. Imaging the Action of Antimicrobial Peptides on Living Bacterial Cells. Sci. Rep. 2013, 3, 1557. [Google Scholar] [CrossRef] [Green Version]
- Hurdle, J.G.; O’Neill, A.J.; Chopra, I.; Lee, R.E. Targeting Bacterial Membrane Function: An Underexploited Mechanism for Treating Persistent Infections. Nat. Rev. Microbiol. 2011, 9, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.V.R.; Yedery, R.D.; Aranha, C. Antimicrobial Peptides: Premises and Promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef]
- Gennaro, R.; Skerlavaj, B.; Romeo, D. Purification, Composition, and Activity of Two Bactenecins, Antibacterial Peptides of Bovine Neutrophils. Infect. Immun. 1989, 57, 3142–3146. [Google Scholar] [CrossRef] [Green Version]
- Björstad, A.; Askarieh, G.; Brown, K.L.; Christenson, K.; Forsman, H.; Onnheim, K.; Li, H.-N.; Teneberg, S.; Maier, O.; Hoekstra, D.; et al. The Host Defense Peptide LL-37 Selectively Permeabilizes Apoptotic Leukocytes. Antimicrob. Agents Chemother. 2009, 53, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- Dürr, U.H.N.; Sudheendra, U.S.; Ramamoorthy, A. LL-37, the Only Human Member of the Cathelicidin Family of Antimicrobial Peptides. Biochim. Biophys. Acta 2006, 1758, 1408–1425. [Google Scholar] [CrossRef] [Green Version]
- Xhindoli, D.; Pacor, S.; Benincasa, M.; Scocchi, M.; Gennaro, R.; Tossi, A. The Human Cathelicidin LL-37—A Pore-Forming Antibacterial Peptide and Host-Cell Modulator. Biochim. Biophys. Acta 2016, 1858, 546–566. [Google Scholar] [CrossRef]
- De Breij, A.; Riool, M.; Cordfunke, R.A.; Malanovic, N.; de Boer, L.; Koning, R.I.; Ravensbergen, E.; Franken, M.; van der Heijde, T.; Boekema, B.K.; et al. The Antimicrobial Peptide SAAP-148 Combats Drug-Resistant Bacteria and Biofilms. Sci. Transl. Med. 2018, 10, eaan4044. [Google Scholar] [CrossRef] [Green Version]
- Dijkmans, A.C.; Wilms, E.B.; Kamerling, I.M.C.; Birkhoff, W.; Ortiz-Zacarías, N.V.; van Nieuwkoop, C.; Verbrugh, H.A.; Touw, D.J. Colistin: Revival of an Old Polymyxin Antibiotic. Ther. Drug Monit. 2015, 37, 419–427. [Google Scholar] [CrossRef]
- Stevenson, C.L. Advances in Peptide Pharmaceuticals. Curr. Pharm. Biotechnol. 2009, 10, 122–137. [Google Scholar] [CrossRef]
- Sierra, J.M.; Fusté, E.; Rabanal, F.; Vinuesa, T.; Viñas, M. An Overview of Antimicrobial Peptides and the Latest Advances in Their Development. Expert Opin. Biol. Ther. 2017, 17, 663–676. [Google Scholar] [CrossRef]
- Lipsky, B.A.; Holroyd, K.J.; Zasloff, M. Topical versus Systemic Antimicrobial Therapy for Treating Mildly Infected Diabetic Foot Ulcers: A Randomized, Controlled, Double-Blinded, Multicenter Trial of Pexiganan Cream. Clin. Infect. Dis. 2008, 47, 1537–1545. [Google Scholar] [CrossRef] [Green Version]
- Rijsbergen, M.; Rijneveld, R.; Todd, M.; Feiss, G.L.; Kouwenhoven, S.T.P.; Quint, K.D.; van Alewijk, D.C.J.G.; de Koning, M.N.C.; Klaassen, E.S.; Burggraaf, J.; et al. Results of Phase 2 Trials Exploring the Safety and Efficacy of Omiganan in Patients with Human Papillomavirus-Induced Genital Lesions. Br. J. Clin. Pharmacol. 2020, 86, 2133–2143. [Google Scholar] [CrossRef]
- Wang, H.; Ai, L.; Zhang, Y.; Cheng, J.; Yu, H.; Li, C.; Zhang, D.; Pan, Y.; Lin, L. The Effects of Antimicrobial Peptide Nal-P-113 on Inhibiting Periodontal Pathogens and Improving Periodontal Status. Biomed. Res. Int. 2018, 2018, 1805793. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial Peptides and Wound Healing: Biological and Therapeutic Considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Spohn, R.; Daruka, L.; Lázár, V.; Martins, A.; Vidovics, F.; Grézal, G.; Méhi, O.; Kintses, B.; Számel, M.; Jangir, P.K.; et al. Integrated Evolutionary Analysis Reveals Antimicrobial Peptides with Limited Resistance. Nat. Commun. 2019, 10, 4538. [Google Scholar] [CrossRef] [Green Version]
- Volejníková, A.; Melicherčík, P.; Nešuta, O.; Vaňková, E.; Bednárová, L.; Rybáček, J.; Čeřovský, V. Antimicrobial Peptides Prevent Bacterial Biofilm Formation on the Surface of Polymethylmethacrylate Bone Cement. J. Med. Microbiol. 2019, 68, 961–972. [Google Scholar] [CrossRef]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Scott, M.G. The Role of Antimicrobial Peptides in Animal Defenses. Proc. Natl. Acad. Sci. USA 2000, 97, 8856–8861. [Google Scholar] [CrossRef]
- Brandenburg, K.; Heinbockel, L.; Correa, W.; Lohner, K. Peptides with Dual Mode of Action: Killing Bacteria and Preventing Endotoxin-Induced Sepsis. Biochim. Biophys. Acta 2016, 1858, 971–979. [Google Scholar] [CrossRef]
- Brandenburg, K. Lack of New Antiinfective Agents: Passing into the Pre-Antibiotic Age? WJBC 2015, 6, 71. [Google Scholar] [CrossRef]
- Brandenburg, K.; Andrä, J.; Garidel, P.; Gutsmann, T. Peptide-Based Treatment of Sepsis. Appl. Microbiol. Biotechnol. 2011, 90, 799–808. [Google Scholar] [CrossRef]
- Dutta, P.; Das, S. Mammalian Antimicrobial Peptides: Promising Therapeutic Targets Against Infection and Chronic Inflammation. CTMC 2015, 16, 99–129. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hong, T.; Cui, P.; Wang, J.; Xia, J. Antimicrobial peptides towards clinical application: Delivery and formulation. Adv. Drug Deliv. Rev. 2021, 175, 113818. [Google Scholar] [CrossRef] [PubMed]
- Atefyekta, S.; Blomstrand, E.; Rajasekharan, A.K.; Svensson, S.; Trobos, M.; Hong, J.; Webster, T.J.; Thomsen, P.; Andersson, M. Antimicrobial Peptide-Functionalized Mesoporous Hydrogels. ACS Biomater. Sci. Eng. 2021, 7, 1693–1702. [Google Scholar] [CrossRef]
- Boto, A.; Pérez de la Lastra, J.; González, C. The Road from Host-Defense Peptides to a New Generation of Antimicrobial Drugs. Molecules 2018, 23, 311. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Koh, J.-J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane Active Antimicrobial Peptides: Translating Mechanistic Insights to Design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Greco, I.; Molchanova, N.; Holmedal, E.; Jenssen, H.; Hummel, B.D.; Watts, J.L.; Håkansson, J.; Hansen, P.R.; Svenson, J. Correlation between Hemolytic Activity, Cytotoxicity and Systemic in Vivo Toxicity of Synthetic Antimicrobial Peptides. Sci. Rep. 2020, 10, 13206. [Google Scholar] [CrossRef] [PubMed]
- Svendsen, J.S.M.; Grant, T.M.; Rennison, D.; Brimble, M.A.; Svenson, J. Very Short and Stable Lactoferricin-Derived Antimicrobial Peptides: Design Principles and Potential Uses. Acc. Chem. Res. 2019, 52, 749–759. [Google Scholar] [CrossRef]
- Yurkova, M.S.; Zenin, V.A.; Sadykhov, E.G.; Fedorov, A.N. Dimerization of Antimicrobial Peptide Polyphemusin I into One Polypeptide Chain: Theoretical and Practical Consequences. Appl. Biochem. Microbiol. 2020, 56, 893–897. [Google Scholar] [CrossRef]
- Irazazabal, L.N.; Porto, W.F.; Ribeiro, S.M.; Casale, S.; Humblot, V.; Ladram, A.; Franco, O.L. Selective Amino Acid Substitution Reduces Cytotoxicity of the Antimicrobial Peptide Mastoparan. Biochim. Biophys. Acta 2016, 1858, 2699–2708. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, M.; Qiu, S.; Wang, J.; Peng, J.; Zhao, P.; Zhu, R.; Wang, H.; Li, Y.; Wang, K.; et al. Antimicrobial Activity and Stability of the D-Amino Acid Substituted Derivatives of Antimicrobial Peptide Polybia-MPI. AMB Express 2016, 6, 122. [Google Scholar] [CrossRef] [Green Version]
- Kindrachuk, J.; Scruten, E.; Attah-Poku, S.; Bell, K.; Potter, A.; Babiuk, L.A.; Griebel, P.J.; Napper, S. Stability, Toxicity, and Biological Activity of Host Defense Peptide BMAP28 and Its Inversed and Retro-Inversed Isomers. Biopolymers 2011, 96, 14–24. [Google Scholar] [CrossRef]
- Carratalá, J.V.; Serna, N.; Villaverde, A.; Vázquez, E.; Ferrer-Miralles, N. Nanostructured Antimicrobial Peptides: The Last Push towards Clinics. Biotechnol. Adv. 2020, 44, 107603. [Google Scholar] [CrossRef] [PubMed]
- Umerska, A.; Cassisa, V.; Matougui, N.; Joly-Guillou, M.-L.; Eveillard, M.; Saulnier, P. Antibacterial Action of Lipid Nanocapsules Containing Fatty Acids or Monoglycerides as Co-Surfactants. Eur. J. Pharm. Biopharm. 2016, 108, 100–110. [Google Scholar] [CrossRef]
- Radaic, A.; de Jesus, M.B.; Kapila, Y.L. Bacterial Anti-Microbial Peptides and Nano-Sized Drug Delivery Systems: The State of the Art toward Improved Bacteriocins. J. Control. Release 2020, 321, 100–118. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Ma, Q.; Chen, X.; Chen, T.; Ying, Y.; Xi, X.; Wang, L.; Ma, C.; Shaw, C.; Zhou, M. Recent Advances and Challenges in Nanodelivery Systems for Antimicrobial Peptides (AMPs). Antibiotics 2021, 10, 990. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, G.; Kim, E.Y.; Shin, S.Y. LL-37-Derived Membrane-Active FK-13 Analogs Possessing Cell Selectivity, Anti-Biofilm Activity and Synergy with Chloramphenicol and Anti-Inflammatory Activity. Biochim. Biophys. Acta 2017, 1859, 722–733. [Google Scholar] [CrossRef]
- Yu, G.; Baeder, D.Y.; Regoes, R.R.; Rolff, J. Combination Effects of Antimicrobial Peptides. Antimicrob. Agents Chemother. 2016, 60, 1717–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheard, D.E.; O’Brien-Simpson, N.M.; Wade, J.D.; Separovic, F. Combating Bacterial Resistance by Combination of Antibiotics with Antimicrobial Peptides. Pure Appl. Chem. 2019, 91, 199–209. [Google Scholar] [CrossRef]
- Xu, X.; Xu, L.; Yuan, G.; Wang, Y.; Qu, Y.; Zhou, M. Synergistic Combination of Two Antimicrobial Agents Closing Each Other’s Mutant Selection Windows to Prevent Antimicrobial Resistance. Sci. Rep. 2018, 8, 7237. [Google Scholar] [CrossRef]
- Jangra, M.; Raka, V.; Nandanwar, H. In Vitro Evaluation of Antimicrobial Peptide Tridecaptin M in Combination with Other Antibiotics against Multidrug Resistant Acinetobacter Baumannii. Molecules 2020, 25, 3255. [Google Scholar] [CrossRef]
- Oo, T.Z.; Cole, N.; Garthwaite, L.; Willcox, M.D.P.; Zhu, H. Evaluation of Synergistic Activity of Bovine Lactoferricin with Antibiotics in Corneal Infection. J. Antimicrob. Chemother. 2010, 65, 1243–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuding, S.; Frasch, T.; Schaller, M.; Stange, E.F.; Zabel, L.T. Synergistic Effects of Antimicrobial Peptides and Antibiotics against Clostridium Difficile. Antimicrob. Agents Chemother. 2014, 58, 5719–5725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahangiri, A.; Neshani, A.; Mirhosseini, S.A.; Ghazvini, K.; Zare, H.; Sedighian, H. Synergistic Effect of Two Antimicrobial Peptides, Nisin and P10 with Conventional Antibiotics against Extensively Drug-Resistant Acinetobacter Baumannii and Colistin-Resistant Pseudomonas Aeruginosa Isolates. Microb. Pathog. 2021, 150, 104700. [Google Scholar] [CrossRef] [PubMed]
- Portelinha, J.; Angeles-Boza, A.M. The Antimicrobial Peptide Gad-1 Clears Pseudomonas Aeruginosa Biofilms under Cystic Fibrosis Conditions. ChemBioChem 2021, 22, 1646–1655. [Google Scholar] [CrossRef]
- Dosler, S.; Mataraci, E. In Vitro Pharmacokinetics of Antimicrobial Cationic Peptides Alone and in Combination with Antibiotics against Methicillin Resistant Staphylococcus Aureus Biofilms. Peptides 2013, 49, 53–58. [Google Scholar] [CrossRef]
- Mataraci, E.; Dosler, S. In Vitro Activities of Antibiotics and Antimicrobial Cationic Peptides Alone and in Combination against Methicillin-Resistant Staphylococcus Aureus Biofilms. Antimicrob. Agents Chemother. 2012, 56, 6366–6371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wu, J.; Cheng, H.; Dai, Y.; Wang, Y.; Yang, H.; Xiong, F.; Xu, W.; Wei, L. Anti-Infective Effects of a Fish-Derived Antimicrobial Peptide Against Drug-Resistant Bacteria and Its Synergistic Effects with Antibiotic. Front. Microbiol. 2020, 11, 602412. [Google Scholar] [CrossRef]
- Song, M.; Liu, Y.; Huang, X.; Ding, S.; Wang, Y.; Shen, J.; Zhu, K. A Broad-Spectrum Antibiotic Adjuvant Reverses Multidrug-Resistant Gram-Negative Pathogens. Nat. Microbiol. 2020, 5, 1040–1050. [Google Scholar] [CrossRef]
- Zarkotou, O.; Pournaras, S.; Tselioti, P.; Dragoumanos, V.; Pitiriga, V.; Ranellou, K.; Prekates, A.; Themeli-Digalaki, K.; Tsakris, A. Predictors of Mortality in Patients with Bloodstream Infections Caused by KPC-Producing Klebsiella Pneumoniae and Impact of Appropriate Antimicrobial Treatment. Clin. Microbiol. Infect. 2011, 17, 1798–1803. [Google Scholar] [CrossRef] [Green Version]
- Bassetti, M.; Righi, E. New Antibiotics and Antimicrobial Combination Therapy for the Treatment of Gram-Negative Bacterial Infections. Curr. Opin. Crit. Care 2015, 21, 402–411. [Google Scholar] [CrossRef]
- Antonoplis, A.; Zang, X.; Huttner, M.A.; Chong, K.K.L.; Lee, Y.B.; Co, J.Y.; Amieva, M.R.; Kline, K.A.; Wender, P.A.; Cegelski, L. A Dual-Function Antibiotic-Transporter Conjugate Exhibits Superior Activity in Sterilizing MRSA Biofilms and Killing Persister Cells. J. Am. Chem. Soc. 2018, 140, 16140–16151. [Google Scholar] [CrossRef]
- Reinhardt, A.; Neundorf, I. Design and Application of Antimicrobial Peptide Conjugates. IJMS 2016, 17, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Chen, F.; Wang, X.; Peng, H.; Zhang, H.; Wang, K.-J. The Synergistic Effect of Mud Crab Antimicrobial Peptides Sphistin and Sph12−38 with Antibiotics Azithromycin and Rifampicin Enhances Bactericidal Activity Against Pseudomonas Aeruginosa. Front. Cell. Infect. Microbiol. 2020, 10, 572849. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, Z.; Li, X.; Tian, Y.; Fan, Y.; Yu, C.; Zhou, B.; Liu, Y.; Xiang, R.; Yang, L. Synergistic Effects of Antimicrobial Peptide DP7 Combined with Antibiotics against Multidrug-Resistant Bacteria. DDDT 2017, 11, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Akbari, R.; Hakemi-Vala, M.; Pashaie, F.; Bevalian, P.; Hashemi, A.; Pooshang Bagheri, K. Highly Synergistic Effects of Melittin with Conventional Antibiotics Against Multidrug-Resistant Isolates of Acinetobacter Baumannii and Pseudomonas Aeruginosa. Microb. Drug Resist. 2019, 25, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Naghmouchi, K.; Le Lay, C.; Baah, J.; Drider, D. Antibiotic and Antimicrobial Peptide Combinations: Synergistic Inhibition of Pseudomonas Fluorescens and Antibiotic-Resistant Variants. Res. Microbiol. 2012, 163, 101–108. [Google Scholar] [CrossRef]
- Kampshoff, F.; Willcox, M.D.P.; Dutta, D. A Pilot Study of the Synergy between Two Antimicrobial Peptides and Two Common Antibiotics. Antibiotics 2019, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Ruden, S.; Rieder, A.; Chis Ster, I.; Schwartz, T.; Mikut, R.; Hilpert, K. Synergy Pattern of Short Cationic Antimicrobial Peptides Against Multidrug-Resistant Pseudomonas Aeruginosa. Front. Microbiol. 2019, 10, 2740. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Lee, D.G. Synergistic Effect of Antimicrobial Peptide Arenicin-1 in Combination with Antibiotics against Pathogenic Bacteria. Res. Microbiol. 2012, 163, 479–486. [Google Scholar] [CrossRef]
- Lee, H.; Lee, D.G. Novel Approaches for Efficient Antifungal Drug Action. J. Microbiol. Biotechnol. 2018, 28, 1771–1781. [Google Scholar] [CrossRef]
- Hanson, M.A.; Dostálová, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaitre, B. Synergy and Remarkable Specificity of Antimicrobial Peptides in Vivo Using a Systematic Knockout Approach. eLife 2019, 8, e44341. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.J.; Purves, J.; Kamysz, W.; Rolff, J. Comparing Selection on S. Aureus between Antimicrobial Peptides and Common Antibiotics. PLoS ONE 2013, 8, e76521. [Google Scholar] [CrossRef] [Green Version]
- Rahnamaeian, M.; Cytryńska, M.; Zdybicka-Barabas, A.; Dobslaff, K.; Wiesner, J.; Twyman, R.M.; Zuchner, T.; Sadd, B.M.; Regoes, R.R.; Schmid-Hempel, P.; et al. Insect Antimicrobial Peptides Show Potentiating Functional Interactions against Gram-Negative Bacteria. Proc. R. Soc. B. 2015, 282, 20150293. [Google Scholar] [CrossRef] [PubMed]
- Casciaro, B.; Loffredo, M.R.; Luca, V.; Verrusio, W.; Cacciafesta, M.; Mangoni, M.L. Esculentin-1a Derived Antipseudomonal Peptides: Limited Induction of Resistance and Synergy with Aztreonam. PPL 2019, 25, 1155–1162. [Google Scholar] [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The Value of Antimicrobial Peptides in the Age of Resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Glattard, E.; Salnikov, E.S.; Aisenbrey, C.; Bechinger, B. Investigations of the Synergistic Enhancement of Antimicrobial Activity in Mixtures of Magainin 2 and PGLa. Biophys. Chem. 2016, 210, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Zerweck, J.; Strandberg, E.; Kukharenko, O.; Reichert, J.; Bürck, J.; Wadhwani, P.; Ulrich, A.S. Molecular Mechanism of Synergy between the Antimicrobial Peptides PGLa and Magainin 2. Sci. Rep. 2017, 7, 13153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilyas, H.; Kim, J.; Lee, D.; Malmsten, M.; Bhunia, A. Structural Insights into the Combinatorial Effects of Antimicrobial Peptides Reveal a Role of Aromatic–Aromatic Interactions in Antibacterial Synergism. J. Biol. Chem. 2019, 294, 14615–14633. [Google Scholar] [CrossRef]
- Najjar, M.B.; Kashtanov, D.; Chikindas, M.L. Epsilon-Poly-l-Lysine and Nisin A Act Synergistically against Gram-Positive Food-Borne Pathogens Bacillus Cereus and Listeria Monocytogenes. Lett. Appl. Microbiol. 2007, 45, 13–18. [Google Scholar] [CrossRef]
- Field, D.; Seisling, N.; Cotter, P.D.; Ross, R.P.; Hill, C. Synergistic Nisin-Polymyxin Combinations for the Control of Pseudomonas Biofilm Formation. Front. Microbiol. 2016, 7, 1713. [Google Scholar] [CrossRef] [Green Version]
- Westerhoff, H.V.; Zasloff, M.; Rosner, J.L.; Hendler, R.W.; Waal, A.; Gomes, A.V.; Jongsma, A.P.M.; Riethorst, A.; Juretic, D. Functional Synergism of the Magainins PGLa and Magainin-2 in Escherichia Coli, Tumor Cells and Liposomes. Eur. J. Biochem. 1995, 228, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Hancock, R.E.W. Synergistic Interactions between Mammalian Antimicrobial Defense Peptides. Antimicrob. Agents Chemother. 2001, 45, 1558–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zdybicka-Barabas, A.; Mak, P.; Klys, A.; Skrzypiec, K.; Mendyk, E.; Fiołka, M.J.; Cytryńska, M. Synergistic Action of Galleria Mellonella Anionic Peptide 2 and Lysozyme against Gram-Negative Bacteria. Biochim. Biophys. Acta 2012, 1818, 2623–2635. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Niyonsaba, F.; Ushio, H.; Okuda, D.; Nagaoka, I.; Ikeda, S.; Okumura, K.; Ogawa, H. Synergistic Effect of Antibacterial Agents Human Beta-Defensins, Cathelicidin LL-37 and Lysozyme against Staphylococcus Aureus and Escherichia Coli. J. Dermatol. Sci. 2005, 40, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhen, Z.; Wang, X.; Guo, N. Synergy of a Combination of Nisin and Citric Acid against Staphylococcus Aureus and Listeria Monocytogenes. Food Addit. Contam. Part. A 2017, 34, 2058–2068. [Google Scholar] [CrossRef]
- Ruden, S.; Hilpert, K.; Berditsch, M.; Wadhwani, P.; Ulrich, A.S. Synergistic Interaction between Silver Nanoparticles and Membrane-Permeabilizing Antimicrobial Peptides. Antimicrob. Agents Chemother. 2009, 53, 3538–3540. [Google Scholar] [CrossRef] [Green Version]
- Doolin, T.; Amir, H.M.; Duong, L.; Rosenzweig, R.; Urban, L.A.; Bosch, M.; Pol, A.; Gross, S.P.; Siryaporn, A. Mammalian Histones Facilitate Antimicrobial Synergy by Disrupting the Bacterial Proton Gradient and Chromosome Organization. Nat. Commun. 2020, 11, 3888. [Google Scholar] [CrossRef]
- Bennett, J.E.; Dolin, E.; Blaser, M.J. Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 9th ed.; Elsevier: Philadelphia, PA, USA, 2019. [Google Scholar]
- Hobbs, J.K.; Miller, K.; O’Neill, A.J.; Chopra, I. Consequences of Daptomycin-Mediated Membrane Damage in Staphylococcus Aureus. J. Antimicrob. Chemother. 2008, 62, 1003–1008. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.; Wenzel, M.; Strahl, H.; Grein, F.; Saaki, T.N.V.; Kohl, B.; Siersma, T.; Bandow, J.E.; Sahl, H.-G.; Schneider, T.; et al. Daptomycin Inhibits Cell Envelope Synthesis by Interfering with Fluid Membrane Microdomains. Proc. Natl. Acad. Sci. USA 2016, 113, E7077–E7086. [Google Scholar] [CrossRef]
- Pogliano, J.; Pogliano, N.; Silverman, J.A. Daptomycin-Mediated Reorganization of Membrane Architecture Causes Mislocalization of Essential Cell Division Proteins. J. Bacteriol. 2012, 194, 4494–4504. [Google Scholar] [CrossRef]
- Konychev, A.; Heep, M.; Moritz, R.K.C.; Kreuter, A.; Shulutko, A.; Fierlbeck, G.; Bouylout, K.; Pathan, R.; Trostmann, U.; Chaves, R.L. Safety and Efficacy of Daptomycin as First-Line Treatment for Complicated Skin and Soft Tissue Infections in Elderly Patients: An Open-Label, Multicentre, Randomized Phase IIIb Trial. Drugs Aging 2013, 30, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, N.; Kusachi, S.; Mikamo, H.; Takesue, Y.; Watanabe, S.; Tanaka, Y.; Morita, A.; Tsumori, K.; Kato, Y.; Yoshinari, T. Efficacy and Safety of Intravenous Daptomycin in Japanese Patients with Skin and Soft Tissue Infections. J. Infect. Chemother. 2013, 19, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telles, J.P.; Cieslinski, J.; Tuon, F.F. Daptomycin to Bone and Joint Infections and Prosthesis Joint Infections: A Systematic Review. Braz. J. Infect. Dis. 2019, 23, 191–196. [Google Scholar] [CrossRef]
- Jaber, R.H.; Beahm, N.P. Daptomycin for the Treatment of Acute Bacterial Meningitis: A Narrative Review. Int. J. Antimicrob. Agents 2023, 61, 106770. [Google Scholar] [CrossRef]
- Rosanova, M.T.; Bes, D.; Serrano-Aguilar, P.; Sberna, N.; Herrera-Ramos, E.; Lede, R.L. Efficacy and Safety of Daptomycin: Systematic Review and Meta-Analysis. Ther. Adv. Infect. Dis. 2019, 6, 2049936119886465. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The Re-Emerging Antibiotic for Multidrug-Resistant Gram-Negative Bacterial Infections. Lancet Infect. Dis. 2006, 6, 589–601. [Google Scholar] [CrossRef]
- From, A.H.; Fong, J.S.; Good, R.A. Polymyxin B Sulfate Modification of Bacterial Endotoxin: Effects on the Development of Endotoxin Shock in Dogs. Infect. Immun. 1979, 23, 660–664. [Google Scholar] [CrossRef]
- Kofteridis, D.P.; Andrianaki, A.M.; Maraki, S.; Mathioudaki, A.; Plataki, M.; Alexopoulou, C.; Ioannou, P.; Samonis, G.; Valachis, A. Treatment Pattern, Prognostic Factors, and Outcome in Patients with Infection Due to Pan-Drug-Resistant Gram-Negative Bacteria. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 965–970. [Google Scholar] [CrossRef]
- Jung, S.Y.; Lee, S.H.; Lee, S.Y.; Yang, S.; Noh, H.; Chung, E.K.; Lee, J.I. Antimicrobials for the Treatment of Drug-Resistant Acinetobacter Baumannii Pneumonia in Critically Ill Patients: A Systemic Review and Bayesian Network Meta-Analysis. Crit. Care 2017, 21, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Shu, Y.; Zhu, F.; Feng, B.; Zhang, Z.; Liu, L.; Wang, G. Comparative Efficacy and Safety of Combination Therapy with High-Dose Sulbactam or Colistin with Additional Antibacterial Agents for Multiple Drug-Resistant and Extensively Drug-Resistant Acinetobacter Baumannii Infections: A Systematic Review and Network Meta-Analysis. J. Glob. Antimicrob. Resist. 2021, 24, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Betrosian, A.P.; Frantzeskaki, F.; Xanthaki, A.; Douzinas, E.E. Efficacy and Safety of High-Dose Ampicillin/Sulbactam vs. Colistin as Monotherapy for the Treatment of Multidrug Resistant Acinetobacter Baumannii Ventilator-Associated Pneumonia. J. Infect. 2008, 56, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Fowler, V.G.; Boucher, H.W.; Corey, G.R.; Abrutyn, E.; Karchmer, A.W.; Rupp, M.E.; Levine, D.P.; Chambers, H.F.; Tally, F.P.; Vigliani, G.A.; et al. Daptomycin versus Standard Therapy for Bacteremia and Endocarditis Caused by Staphylococcus Aureus. N. Engl. J. Med. 2006, 355, 653–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.L.; Slatkin, M.H.; Johnson, B.A. Local Use of Bacitracin. J. Investig. Dermatol. 1948, 10, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Grönberg, A.; Mahlapuu, M.; Ståhle, M.; Whately-Smith, C.; Rollman, O. Treatment with LL-37 Is Safe and Effective in Enhancing Healing of Hard-to-Heal Venous Leg Ulcers: A Randomized, Placebo-Controlled Clinical Trial. Wound Repair. Regen. 2014, 22, 613–621. [Google Scholar] [CrossRef]
- Daley, P.; Louie, T.; Lutz, J.E.; Khanna, S.; Stoutenburgh, U.; Jin, M.; Adedoyin, A.; Chesnel, L.; Guris, D.; Larson, K.B.; et al. Surotomycin versus Vancomycin in Adults with Clostridium Difficile Infection: Primary Clinical Outcomes from the Second Pivotal, Randomized, Double-Blind, Phase 3 Trial. J. Antimicrob. Chemother. 2017, 72, 3462–3470. [Google Scholar] [CrossRef]
- Niemeyer-van der Kolk, T.; van der Wall, H.; Hogendoorn, G.K.; Rijneveld, R.; Luijten, S.; van Alewijk, D.C.J.G.; van den Munckhof, E.H.A.; de Kam, M.L.; Feiss, G.L.; Prens, E.P.; et al. Pharmacodynamic Effects of Topical Omiganan in Patients with Mild to Moderate Atopic Dermatitis in a Randomized, Placebo-Controlled, Phase II Trial. Clin. Transl. Sci. 2020, 13, 994–1003. [Google Scholar] [CrossRef]
- Peek, N.F.A.W.; Nell, M.J.; Brand, R.; Jansen-Werkhoven, T.; van Hoogdalem, E.J.; Verrijk, R.; Vonk, M.J.; Wafelman, A.R.; Valentijn, A.R.P.M.; Frijns, J.H.M.; et al. Ototopical Drops Containing a Novel Antibacterial Synthetic Peptide: Safety and Efficacy in Adults with Chronic Suppurative Otitis Media. PLoS ONE 2020, 15, e0231573. [Google Scholar] [CrossRef] [Green Version]
- Mercer, D.K.; Robertson, J.C.; Miller, L.; Stewart, C.S.; O’Neil, D.A. NP213 (Novexatin®): A Unique Therapy Candidate for Onychomycosis with a Differentiated Safety and Efficacy Profile. Med. Mycol. 2020, 58, 1064–1072. [Google Scholar] [CrossRef] [Green Version]
- Mullane, K.; Lee, C.; Bressler, A.; Buitrago, M.; Weiss, K.; Dabovic, K.; Praestgaard, J.; Leeds, J.A.; Blais, J.; Pertel, P. Multicenter, Randomized Clinical Trial to Compare the Safety and Efficacy of LFF571 and Vancomycin for Clostridium Difficile Infections. Antimicrob. Agents Chemother. 2015, 59, 1435–1440. [Google Scholar] [CrossRef] [Green Version]
- Corey, R.; Naderer, O.J.; O’Riordan, W.D.; Dumont, E.; Jones, L.S.; Kurtinecz, M.; Zhu, J.Z. Safety, Tolerability, and Efficacy of GSK1322322 in the Treatment of Acute Bacterial Skin and Skin Structure Infections. Antimicrob. Agents Chemother. 2014, 58, 6518–6527. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Name | Source | Mechanism of Action | Ref |
|---|---|---|---|
| Abaecin | Bumblebee | Inhibits DnaK | [113] |
| Alamethicin | Fungus Trichoderma viride | Barrel-stave transmembrane pore model | [114] |
| Apidaecin Hb1a | Honeybee | Inhibits protein biosynthesis by targeting ribosomes, inhibits DnaK and GroEL, inhibits ABC transport system, binds LPS | [115] |
| Buforin II | Enzymatic cleavage of buforin I (from Asian toad Bufo bufo gargarizans) | Inhibits DNA, inhibits RNA | [100] |
| Cecropin P1 | Pig intestine | Carpet model/Detergent-like mode non-membrane pore model | [116] |
| Diptericin | Hemolymph of injured Sarcophaga peregrine larva | Inhibits septation | [117] |
| HD5 (oxidized) | Human small intestine | Inhibits cell division | [118] |
| Histatin-5 | Human saliva | Inhibits proteases, inhibits MMP-2 and MMP-9, inhibits generation of ROS | [119] |
| Lacticin Q | Lactococcus lactis QU 5 | Toroidal transmembrane pore model | [120] |
| Magainin 1 | African clawed frog Xenopus laevis | Inhibits energy metabolism proteins, inhibits amino-acid metabolism | [121] |
| MBI-27 | Derived from part of silk moth cecropin and bee melittin peptides | Inhibits LPS | [122] |
| Nisin | Lactococcus lactis | Inhibits lipid II in peptidoglycan biosynthesis | [123] |
| Ostricacin-1 | Ostrich defensin | Inhibits DNA | [124] |
| Tachyplesin | Horseshoe crab hemocytes | Binds DNA minor groove | [125] |
| Thanatin | Podisus maculiventris | Agglutination non-membrane pore model | [126] |
| tPMP-1 | Platelet granules | Activation of autolytic enzyme | [127] |
| AMP | Antibiotic | Target | Ref |
|---|---|---|---|
| Tridecaptin M | Rifampicin, vancomycin, ceftazidime | A. baumannii | [222] |
| Lactoferricin | Ciprofloxacin, ceftazidime | P. aeruginosa | [223] |
| LL-37, HBD3 | Tigecycline, moxifloxacin, piperacillin/tazobactam, meropenem | C. difficile | [224] |
| P10 | Ceftazidime, doripenem | A. baumannii and P. aeruginosa | [225] |
| Gad-1 | Kanamycin, ciprofloxacin | P. aeruginosa | [226] |
| Nisin | Penicillin, chloramphenicol, ciprofloxacin, indolicidin, or azithromycin | S. aureus | [227,228] |
| SAAP-148 | Demeclocycline hydrochloride (DMCT) | P. aeruginosa | [229] |
| (SLAP)-S25 | Cefepime, colistin, ofloxacin, rifampicin, tetracycline, and vancomycin | multidrug-resistant Gram-negative pathogens | [230] |
| Colistin | tigecycline, carbapenem, gentamicin | Klebsiella KPC | [231,232] |
| Octaarginine | Vancomycin | biofilms of S. aureus | [233,234] |
| Sphistin, Sph12−38 | Rifampicin, azithromycin | P. aeruginosa | [235] |
| DP7 | Azithromycin, vancomycin | S. aureus, P. aeruginosa, A. baumannii, E. coli | [236] |
| P10 | Ceftazidime, doripenem | A. baumannii, colistin-resistant P. aeruginosa | [225] |
| Melittin | Doripenem and ceftazidime | A. baumannii and P. aeruginosa | [237] |
| LL 17–29 | Chloramphenicol | S. aureus, P. aeruginosa | [218] |
| Nisin Z, pediocin, or colistin | Penicillin, ampicillin, or rifampicin | P. fluorescens | [238] |
| Melamine | Ciprofloxacin, fluoroquinolone | P. aeruginosa | [239] |
| Indolicidin, polymyxin B | Tobramycin, gentamycin, and amikacin | P. aeruginosa | [240] |
| Arenicin-1 | Ampicillin, erythromycin, and chloramphenicol | S. aureus, S. epidermis, P. aeruginosa, and E. coli | [241] |
| AMP | Synergistic AMP Molecule | Target | Ref |
|---|---|---|---|
| HsAFP1 | RsAFP2 and RsAFP1 | Biofilm cells | [242] |
| Diptericins | Attacins | P. burhodogranariea | [243] |
| Pexiganan | Melittin | S. aureus | [244] |
| Abaecin | Hymenoptaecin | Gram-negative bacteria | [245] |
| Esculentin-1a | Aztreonam | P. aeruginosa | [246] |
| PGLa | Magainin 2 | E. coli | [247,248,249] |
| VG16KRKP | KYE28 | plant pathogens | [250] |
| Nisin A | Epsilon-poly-L-lysine | Bacillus cereus and L. monocytogenes | [251] |
| Nisin | Colistin | Biofilms of P. aeruginosa | [225,252] |
| Nisin | Colistin | A. baumannii, colistin-resistant P. aeruginosa | [225] |
| Magainin-2 | Peptidyl-glycylleucine-carboxyamide | E. coli | [253] |
| Indolicidin, LL-37 | Bactenecin | P. aeruginosa and E. coli | [254] |
| Indolicidin | Bactenecin | P. aeruginosa and E. coli | [254] |
| Protegrin 1 | LL-37 | E. faecalis | [254] |
| Bactenecin | LL-37 | E. faecalis | [254] |
| Protegrin1 | Bactenecin | E. faecalis | [254] |
| Apidaecin, pexiganan | LL 19–27 | E. coli | [219] |
| Galleria mellonella anionic peptide 2 | Lysozyme | Gram-negative bacteria | [255] |
| B-defensin, LL-37 | Lysozyme | S. aureus, E. coli | [256] |
| AMP | Synergistic Compound | Target | Ref |
|---|---|---|---|
| Nisin | Citric acid | S. aureus and L. monocytogenes | [257] |
| Nisin A | Epsilon-poly-L-lysine | Gram-positive food-borne pathogens Bacillus cereus and L. monocytogenes | [251] |
| Polymyxin B, Gramicidin S | Silver nitrate, silver nanoparticles | Gram-negative bacteria | [258] |
| AMP | Histone | Target | Ref |
|---|---|---|---|
| LL-37 and magainin-2 | H2A and H3 | Gram-positive and Gram-negative bacteria | [259] |
| Polymyxin B | H2A | E. coli | [259] |
| Study | Population | Intervention | Comparator | Outcome |
|---|---|---|---|---|
| Fowler et al., 2006 [275]—Phase III RCT | Patients with S. aureus bacteremia with or without endocarditis | IV treatment with the AMP daptomycin | IV treatment with low-dose gentamicin plus either an antistaphylococcal penicillin or vancomycin | Daptomycin was non-inferior to standard-of-care. Treatment success rates were similar in subgroups of patients with complicated bacteremia, right-sided endocarditis, and methicillin-resistant S. aureus |
| Miller et al., 1948 [276]—non-randomized study | 130 patients with superficial infections of the skin and 35 patients with secondary skin infections | Bacitracin applied locally | None | Cure rate higher than 50% in superficial skin infections and 100% in secondary skin infections |
| NCT05340790—Phase I RCT | Healthy female volunteers | Dose 1 to 5 of AMP PL-18 vaginal suppositories | Placebo doses 1 to 5 of vaginal suppositories | Safety assessment (recruiting) |
| Gronberg et al., 2014 [277]—RCT | Adult patients with hard-to-heal venous leg ulcers | Repeated doses of LL-37 applied locally | Repeated doses of placebo applied locally | Safe and well tolerated. Significant early healing of ulcers |
| Daley et al., 2017 [278]—Phase III RCT | Adult patients with Clostridioides difficile infection | AMP surotomycin orally | Vancomycin orally | Non-inferior but non-superior to vancomycin for clinical response |
| Lipsky et al., 2008 [192]—Phase III RCT | Adult diabetic patients with infected wounds at the lower extremities | AMP pexiganan locally and oral placebo | Local placebo and oral ofloxacin | Pexiganan is comparable to oral ofloxacin for mildly infected diabetic ulcers |
| Niemeyer-van der Kolk et al., 2020 [279]—Phase II RCT | Patients 18–65 years old with mild-to-moderate atopic dermatitis | AMP Omiganan locally | Vehicle gel locally (placebo) | Treatment with the AMP improved dysbiosis in mild to moderate atopic dermatitis patients Small improvements in clinical scores were detected |
| Peek et al., 2020 [280]—Phase II RCT | Adults with chronic suppurative otitis media resistant to antibiotic therapy | AMP P60.4Ac locally with ear drops | Vehicle locally with ear drops (placebo) | Safe and well-tolerated treatment. Significantly higher treatment success than placebo |
| NCT00231153, Phase III RCT | Patients with central venous catheters | AMP Omiganan 1% gel local application at the catheter insertion site | Povidone-Iodine 10% local application at the catheter insertion site | Failed to show adequate efficacy in catheter-associated infections |
| NCT04767321, Phase I/II RCT | Adults 18–65 years old with persistent carriage of S. aureus | Nasal application of the AMP LTX-109 | Nasal application of placebo | Safety, tolerability, and microbial eradication—recruitment completed |
| Mercer et al., 2020 [281]—Phase I and Phase II RCTs | 12, 48, and 47 patients with onychomycosis of the toenail | Local application of NP213 | Local application of placebo | NP213 clinical safety profile. Positive patient-reported outcomes |
| Mullane et al., 2015 [282]—Phase II RCT | 72 patients with Clostridioides difficile infection | LFF571 orally | Vancomycin orally | The rate of clinical cure was non-inferior to that of vancomycin. Similar 30-day sustained cure rates. More adverse events for LFF571. |
| Corey et al., 2014 [283]—Phase IIa RCT | 84 adult patients with ABSSSI | GSK1322322 orally | Linezolid orally | Clinical success in the ITT population and the per-protocol population were 67 and 91% in the GSK1322322-treated group and 89 and 100% in the linezolid-treated group |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ioannou, P.; Baliou, S.; Kofteridis, D.P. Antimicrobial Peptides in Infectious Diseases and Beyond—A Narrative Review. Life 2023, 13, 1651. https://doi.org/10.3390/life13081651
Ioannou P, Baliou S, Kofteridis DP. Antimicrobial Peptides in Infectious Diseases and Beyond—A Narrative Review. Life. 2023; 13(8):1651. https://doi.org/10.3390/life13081651
Chicago/Turabian StyleIoannou, Petros, Stella Baliou, and Diamantis P. Kofteridis. 2023. "Antimicrobial Peptides in Infectious Diseases and Beyond—A Narrative Review" Life 13, no. 8: 1651. https://doi.org/10.3390/life13081651