

New Species of Neocosmospora (Ascomycota) from China as Evidenced by Morphological and Molecular Data

Abstract
:1. Introduction
2. Materials and Methods
3. Results
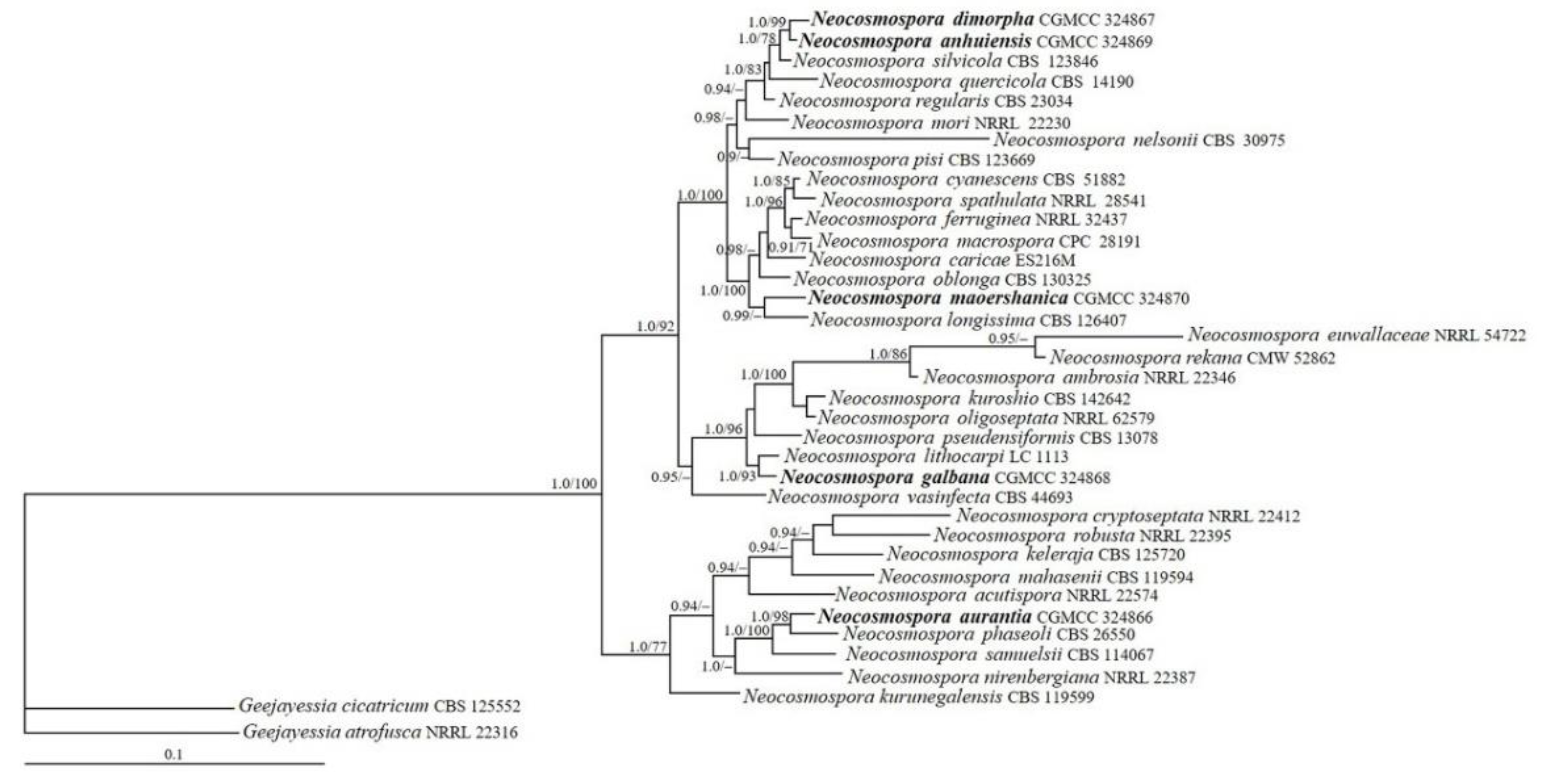
3.1. Phylogenetic Analyses
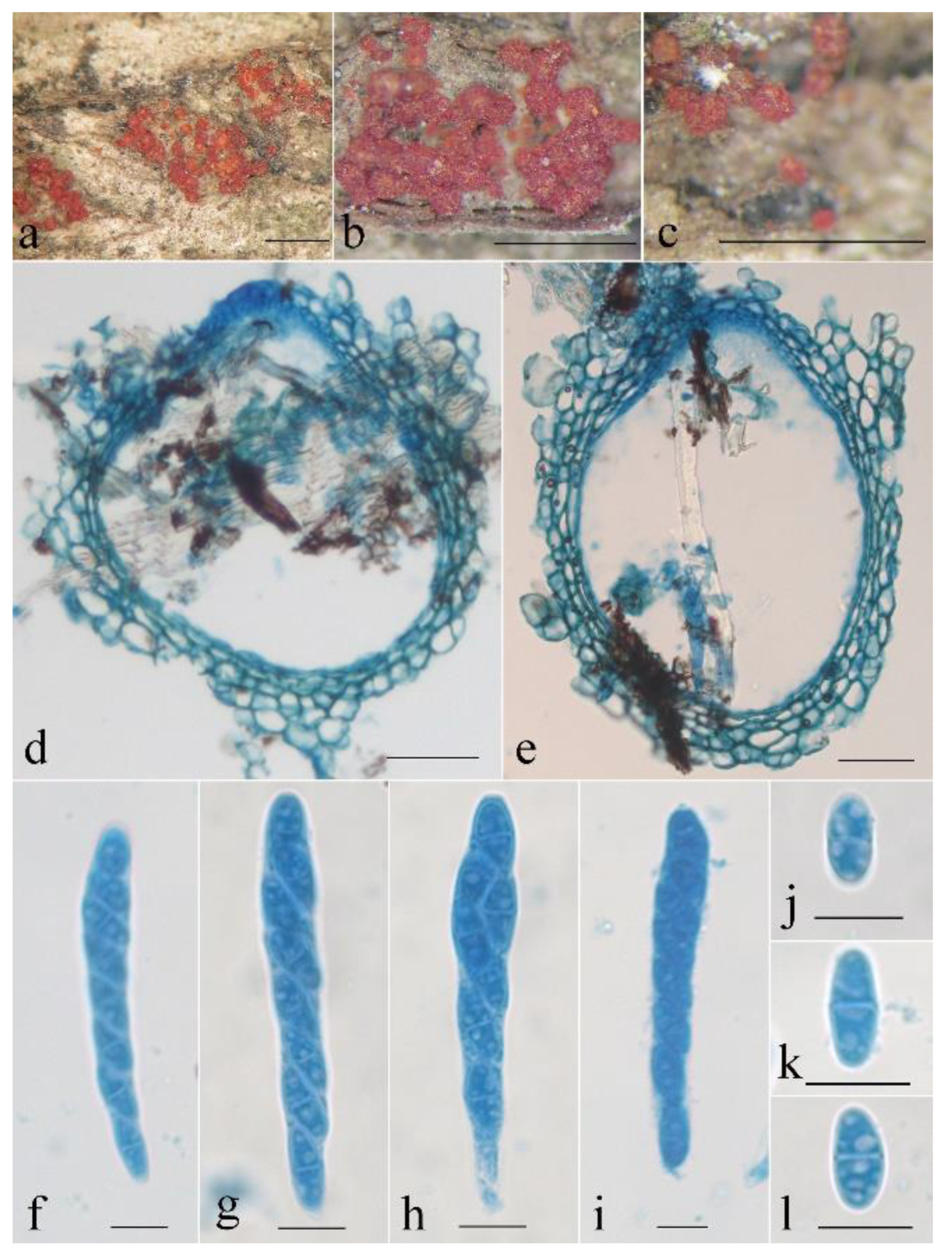
3.2. Taxonomy
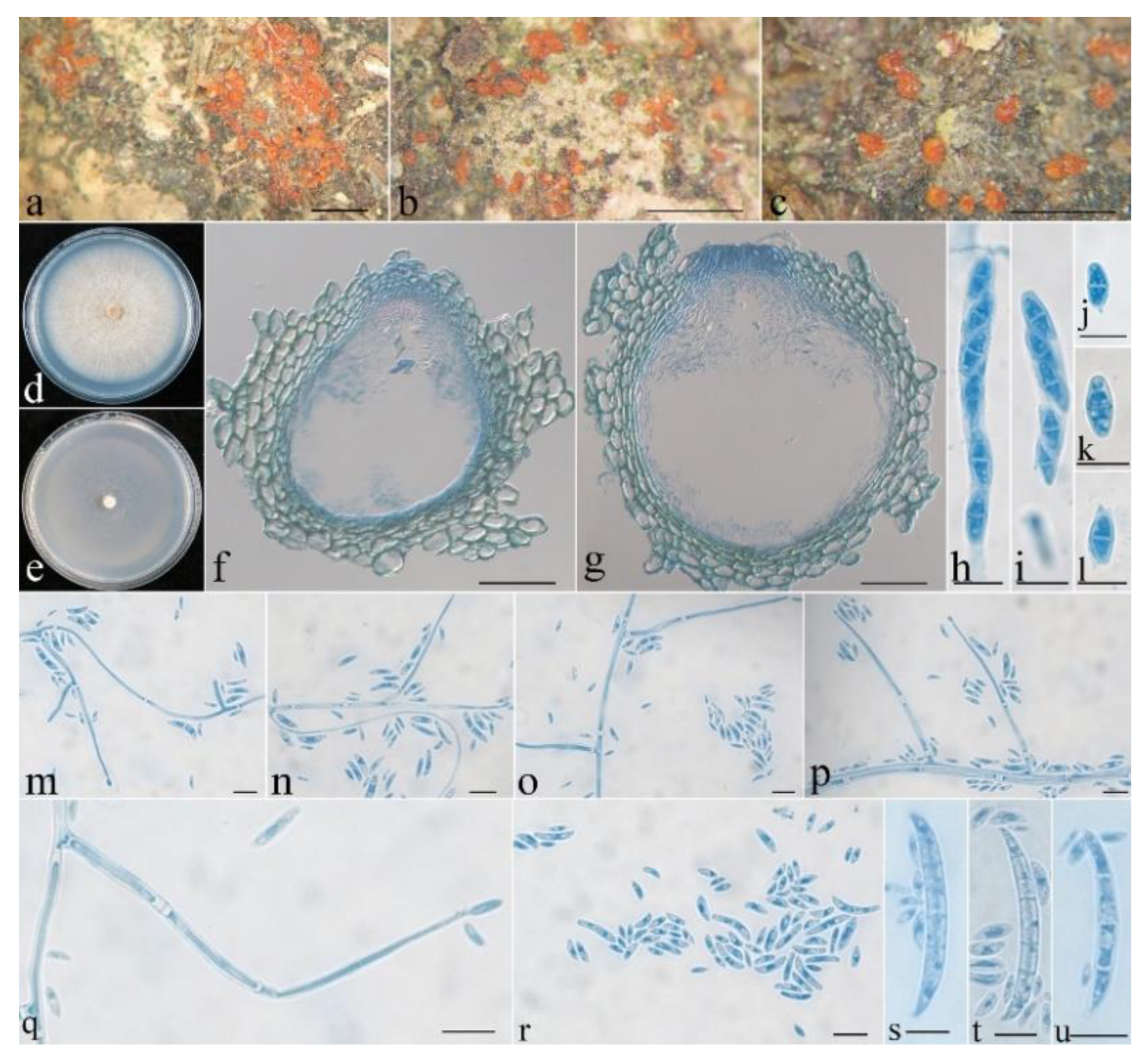
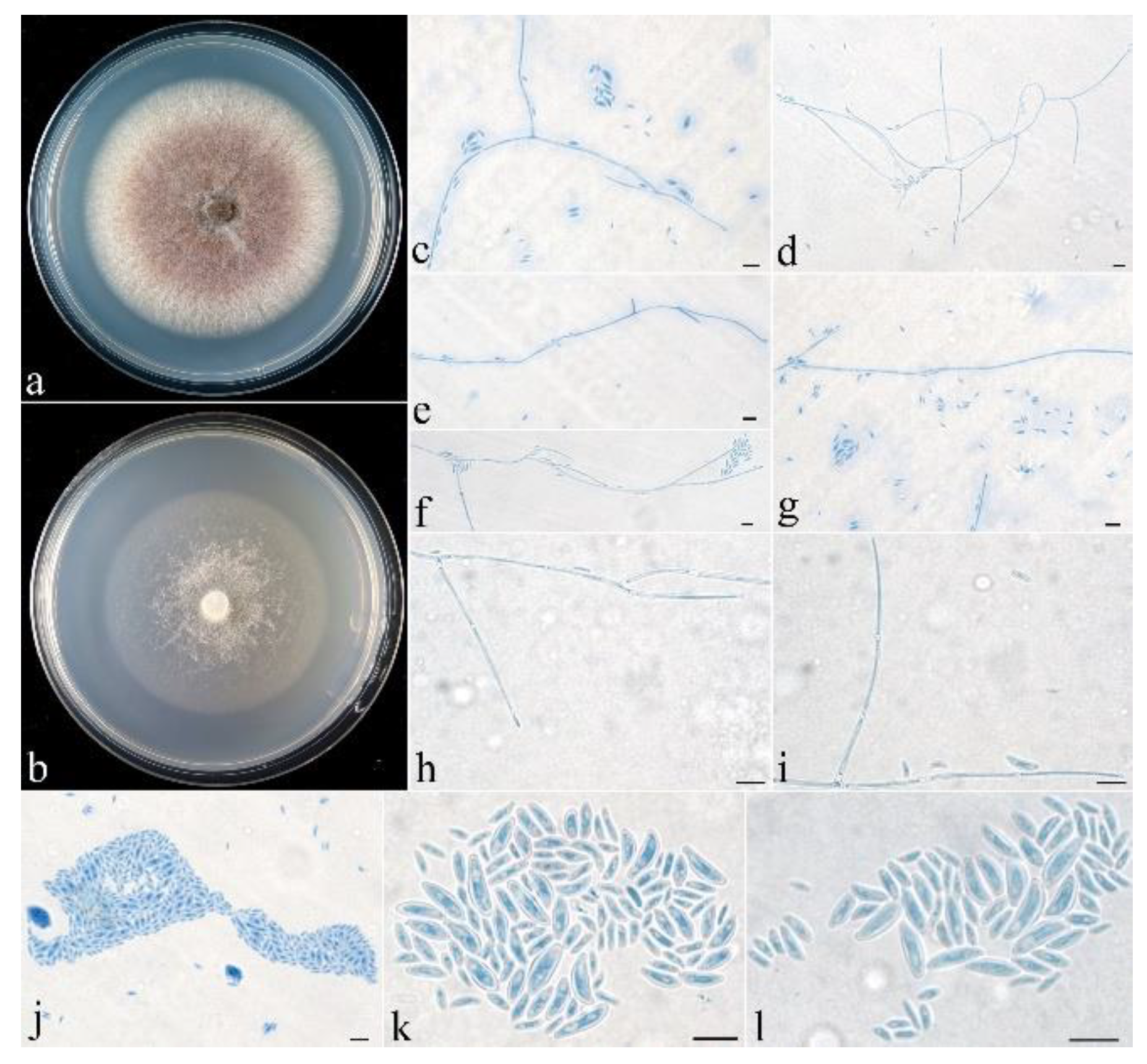
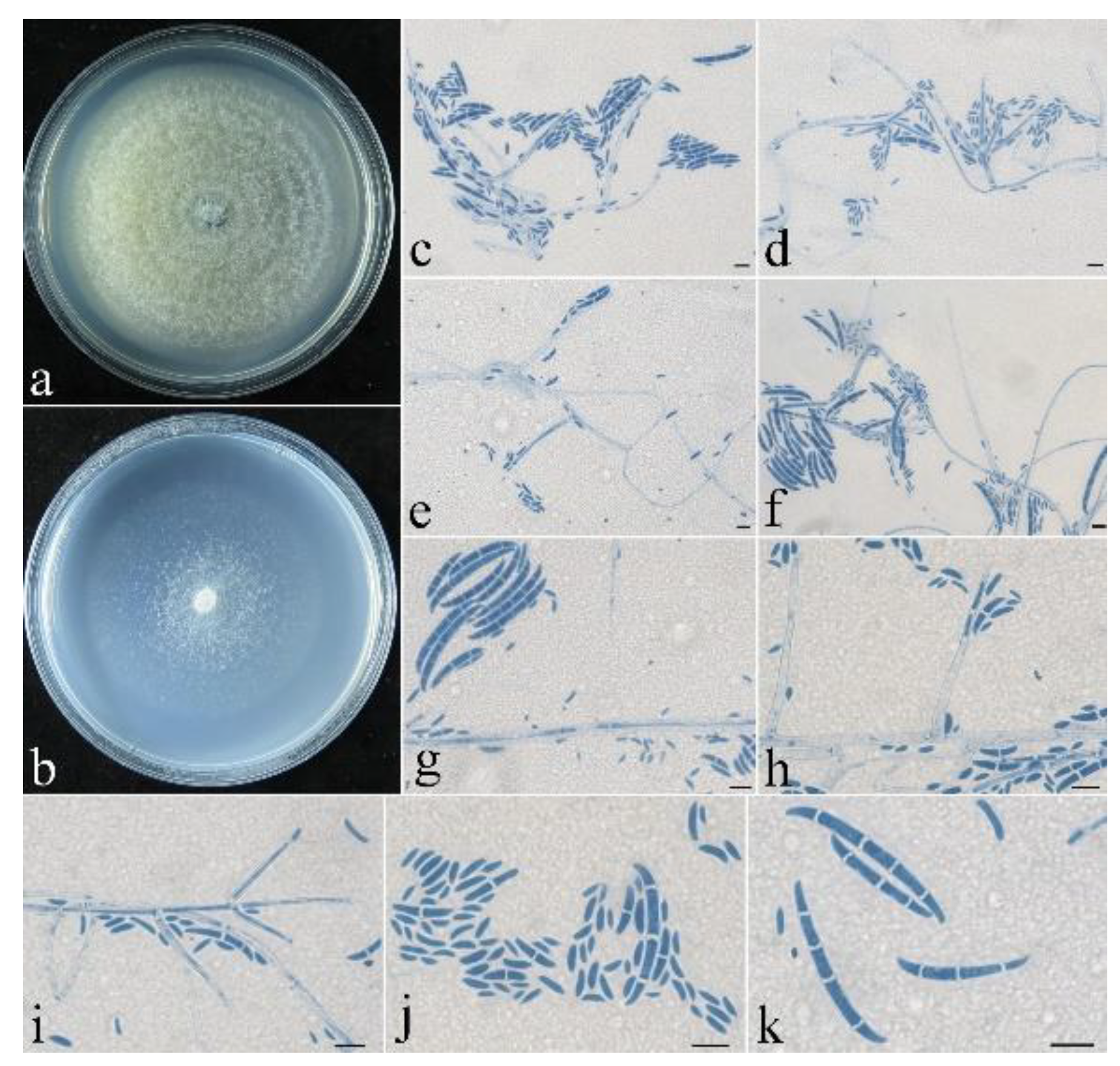
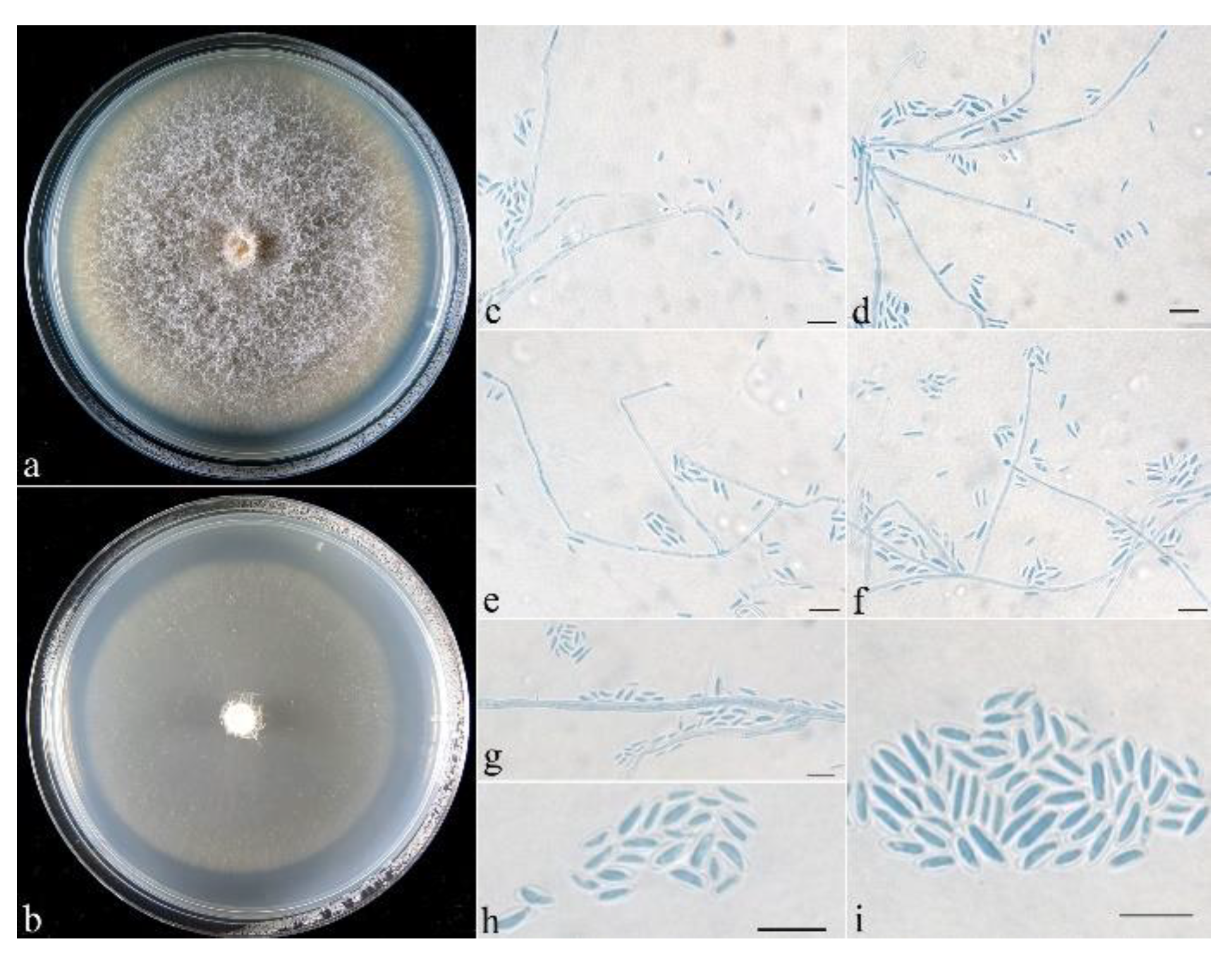
- Neocosmospora anhuiensis Z.Q. Zeng & W.Y. Zhuang, sp. nov. Figure 2.
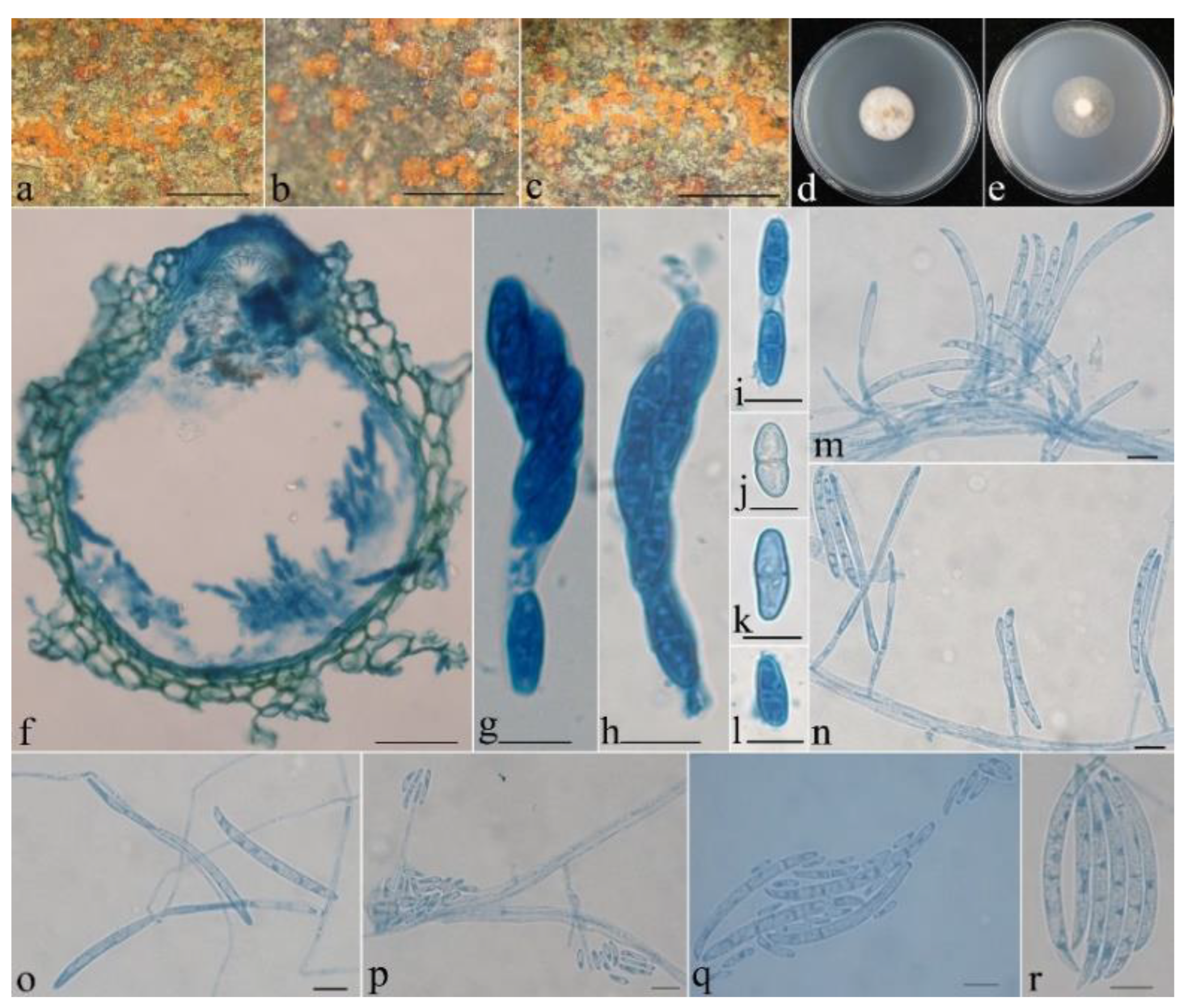
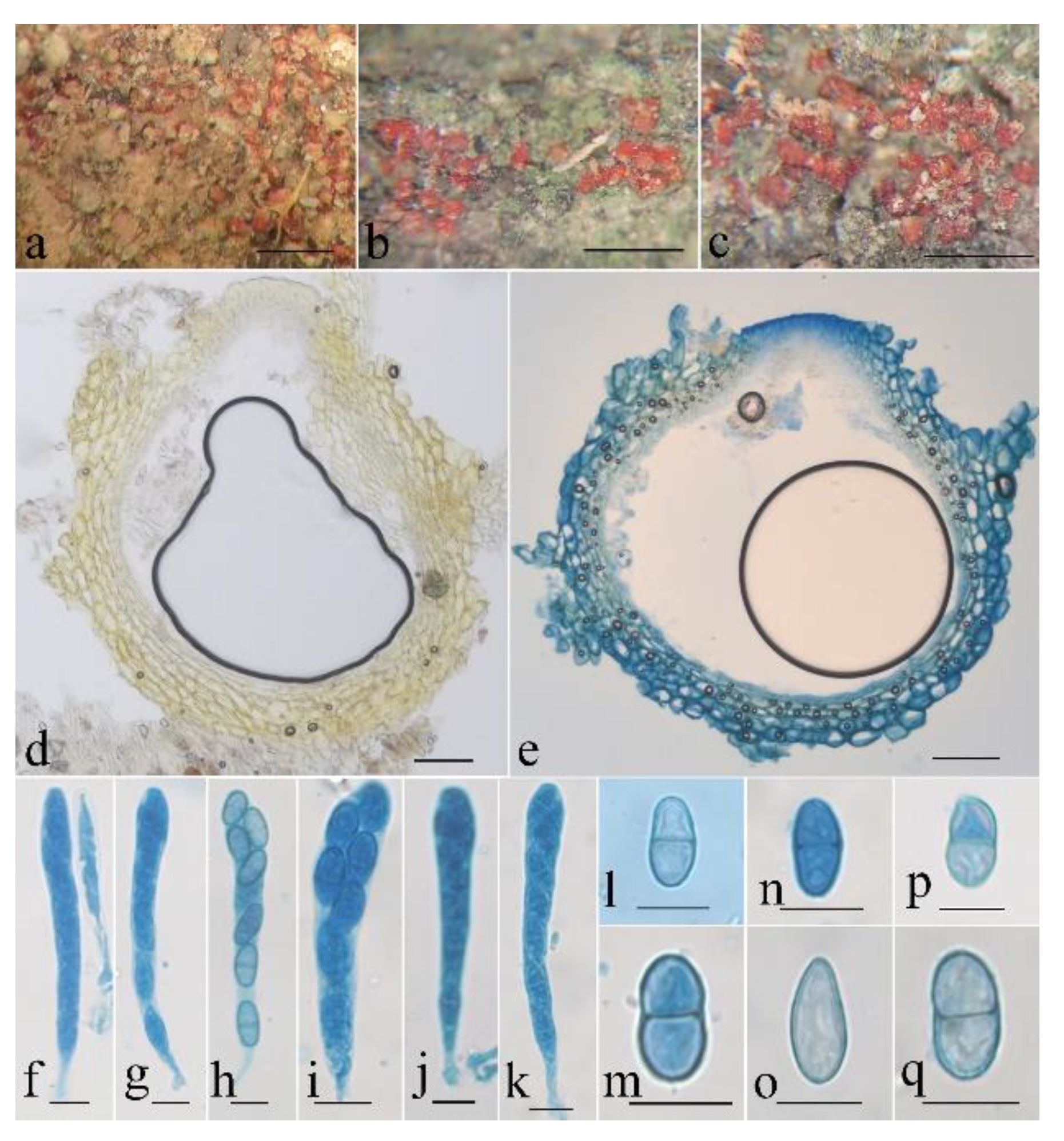
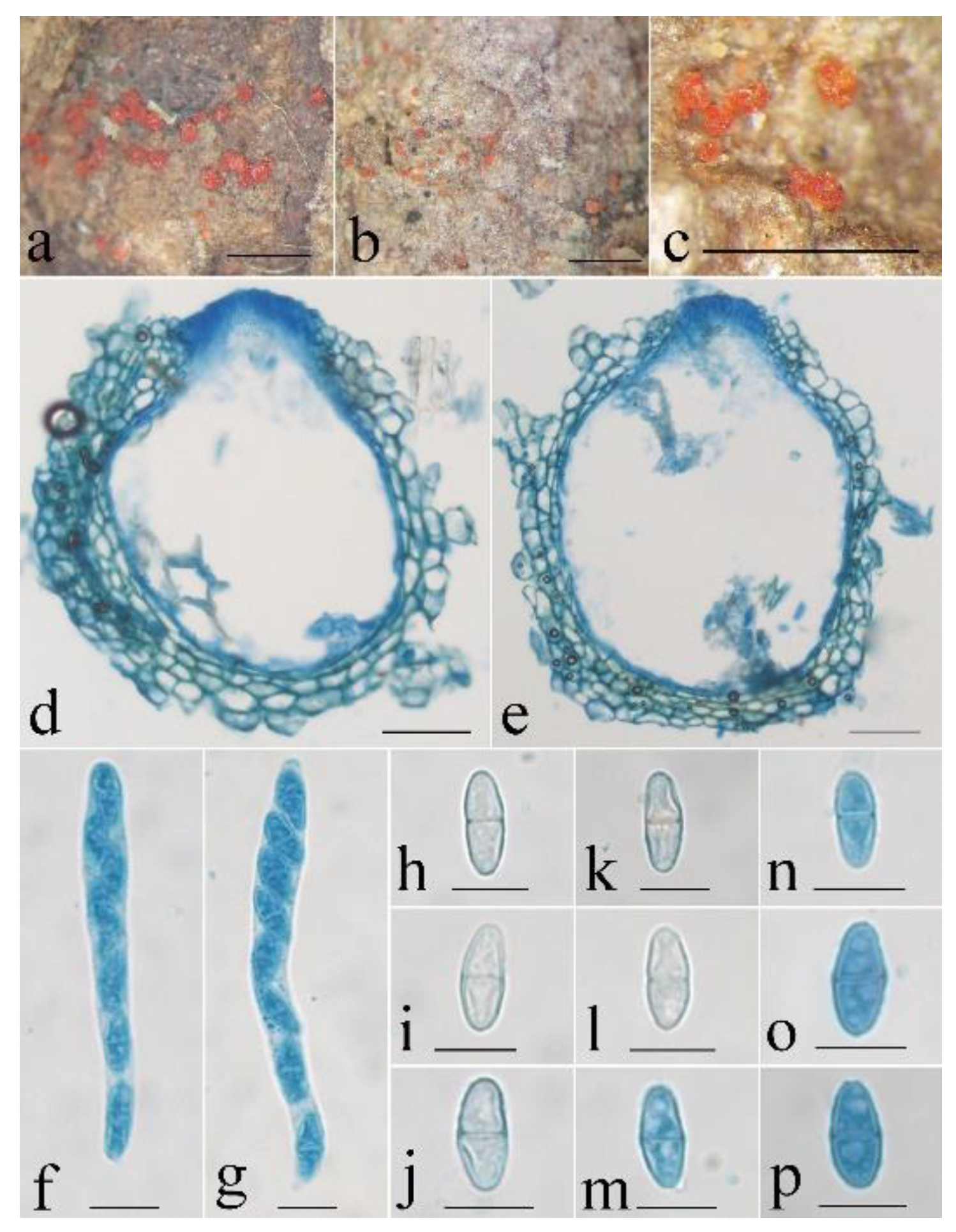
- Neocosmospora aurantia Z.Q. Zeng & W.Y. Zhuang, sp. nov. Figure 3.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rossman, A.Y.; Samuels, G.J.; Rogerson, C.T.; Lowen, R. Genera of Bionectriaceae, Hypocreaceae and Nectriaceae (Hypocreales, Ascomycetes). Stud. Mycol. 1999, 42, 1–248. [Google Scholar]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [Green Version]
- Karunarathna, S.C.; Dong, Y.; Karasaki, S.; Tibpromma, S.; Hyde, K.D.; Lumyong, S.; Xu, J.C.; Sheng, J.; Mortimer, P.E. Discovery of novel fungal species and pathogens on bat carcasses in a cave in Yunnan Province, China. Emerg. Microbes Infec. 2020, 9, 1554–1566. [Google Scholar] [CrossRef]
- Perera, R.H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; McKenzie, E.H.C.; Stadler, M.; Lee, H.B.; Samarakoon, M.C.; Ekanayaka, A.H.; Camporesi, E.; et al. Fungi on wild seeds and fruits. Mycosphere 2020, 11, 2108–2480. [Google Scholar] [CrossRef]
- Perera, R.H.; Hyde, K.D.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Bundhun, D.; Camporesi, E.; Akulov, A.; Liu, J.K.; Liu, Z.Y. Profile of Bionectriaceae, Calcarisporiaceae, Hypocreaceae, Nectriaceae, Tilachlidiaceae, Ijuhyaceae fam. nov., Stromatonectriaceae fam. nov. and Xanthonectriaceae fam. nov. Fungal Divers. 2023, 118, 95–271. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.J.; Chaverri, P.; Gene, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 1–184. [Google Scholar] [CrossRef]
- Crous, P.W.; Sandoval-Denis, M.; Costa, M.M.; Groenewald, J.Z.; van Iperen, A.L.; Starink-Willemse, M.; Hernández-Restrepo, M.; Kandemir, H.; Ulaszewski, B.; de Boer, W.; et al. Fusarium and allied fusarioid taxa (FUSA). 1. Fungal Syst. Evol. 2022, 9, 161–200. [Google Scholar] [CrossRef]
- Guarnaccia, V.; van Niekerk, J.; Crous, P.W.; Sandoval-Denis, M. Neocosmospora spp. associated with dry root rot of citrus in South Africa. Phytopathol. Mediterr. 2021, 60, 79–100. [Google Scholar] [CrossRef]
- Wang, M.M.; Crous, P.W.; Sandoval-Denis, M.; Han, S.L.; Liu, F.; Liang, J.M.; Duan, W.J.; Cai, L. Fusarium and allied genera from China: Species diversity and distribution. Persoonia 2022, 48, 1–53. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Lombard, L.; Crous, P.W. Back to the roots: A reappraisal of Neocosmospora. Persoonia 2019, 43, 90–185. [Google Scholar] [CrossRef] [PubMed]
- Merlin, J.N.; Christudas, I.V.S.N.; Kumar, P.P.; Agastian, P. Optimization of growth and bioactive metabolite production: Fusarium solani. Asian Journal. Pharm. Clin. Res. 2013, 6, 98–103. [Google Scholar]
- Wu, Y.R.; Nian, D.L. Production optimization and molecular structure characterization of a newly isolated novel laccase from Fusarium solani MAS2, an anthracene-degrading fungus. Int. Biodeter. Biodegr. 2014, 86, 382–389. [Google Scholar] [CrossRef]
- Jallouli, R.; Othman, H.; Amara, S.; Parsiegla, G.; Carriere, F.; Srairi-abid, N.; Gargouri, Y.; Bezzine, S. The galactolipase activity of Fusarium solani (phospho) lipase. BBA-Mol. Cell. Biol. Lipids 2015, 1851, 282–289. [Google Scholar] [CrossRef]
- Baard, S.W.; van Wyk, P.S. Neocosmospora vasinfecta pathogenic to groundnuts in South Africa. Phytophylactica 1985, 17, 49–50. [Google Scholar]
- Huang, J.W.; Chen, S.S.; Chung, W.C. Neocosmospora foot rot of peanut in Taiwan. Plant Pathol. Bull. 1992, 1, 203–205. [Google Scholar]
- Fuhlbohm, M.F.; Tatnell, J.R.; Ryley, M.J. Neocosmospora vasinfecta is pathogenic on peanut in Queensland. Australas. Plant Dis. Notes 2007, 2, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S.; Sharon, M.; Maymon, M.; Mendel, Z.; Protasov, A.; Aoki, T.; Eskalen, A.; O’Donnell, K. Fusarium euwallaceae sp. nov.—A symbiotic fungus of Euwallacea sp., an invasive ambrosia beetle in Israel and California. Mycologia 2013, 105, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- Guarnaccia, V.; Sandoval-Denis, M.; Aiello, D.; Polizzi, G.; Crous, P.W. Neocosmospora perseae sp. nov., causing trunk cankers on avocado in Italy. Fungal Syst. Evol. 2018, 1, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Guarnaccia, V.; Martino, I.; Brondino, L.; Gullino, M.L. Paraconiothyrium fuckelii, Diaporthe eres and Neocosmospora parceramosa causing cane blight of red raspberry in Northern Italy. J. Plant Pathol. 2022, 104, 683–698. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Guarnaccia, V.; Polizzi, G.; Crous, P.W. Symptomatic Citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Persoonia 2018, 40, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, F.; Xu, G.; Zheng, F.Q.; Ding, X.F.; Xie, C.P. Neocosmospora rubicola causing stem rot of pitaya (Hylocereus costaricensis) in China. Plant Dis. 2018, 102, 2653. [Google Scholar] [CrossRef]
- Crespo, M.; Lawrence, D.P.; Nouri, M.T.; Doll, D.A.; Trouillas, F.P. Characterization of Fusarium and Neocosmospora species associated with crown rot and stem canker of pistachio rootstocks in California. Plant Dis. 2019, 103, 1931–1939. [Google Scholar] [CrossRef]
- Polat, Z.; Gültekin, M.A.; Palacıoğlu, G.; Bayraktar, H.; Özer, N.; Yılmaz, S. First report of Neocosmospora solani causing stem canker on Juglans regia in Turkey. J. Plant Pathol. 2020, 102, 1289. [Google Scholar] [CrossRef]
- Riaz, M.; Akhtar, N.; Khan, S.N.; Shakeel, M.; Tahir, A. Neocosmospora rubicola: An unrecorded pathogen from Pakistan causing potato stem rot. Sarhad J. Agric. 2020, 36, 906–912. [Google Scholar] [CrossRef]
- Bolboli, Z.; Mostowfizadeh-Ghalamfarsa, R.; Sandoval-Denis, M.; Jafari, M.; Crous, P.W. Neocosmospora caricae sp. nov. and N. metavorans, two new stem and trunk canker pathogens on Ficus carica in Iran. Mycol. Prog. 2022, 21, 89. [Google Scholar] [CrossRef]
- Luo, J.; Zhuang, W.Y. Three new species of Neonectria (Nectriaceae, Hypocreales) with notes on their phylogenetic positions. Mycologia 2010, 102, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Nirenberg, H.I. Untersuchungen über die morphologische und biologische Differenzierung in der Fusarium-Sektion Liseola. Mitt. Biol. Bundesanst. Land. Forstw. 1976, 169, 1–117. [Google Scholar]
- Wang, L.; Zhuang, W.Y. Designing primer sets for amplification of partial calmodulin genes from penicillia. Mycosystema 2004, 23, 466–473. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Tacke, B.K.; Casper, H.H. Gene genealogies reveal global phylogeographic structure and reproductive isolation among lineages of Fusarium graminearum, the fungus causing wheat scab. Proc. Natl. Acad. Sci. USA 2000, 95, 7905–7910. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgin, D.G. The ClustalX windows interface: Flexible strategies for multiple sequences alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Mol. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest, v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comp. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [PubMed] [Green Version]
- Nalim, F.A.; Samuels, G.J.; Wijesundera, R.L.; Geiser, D.M. New species from the Fusarium solani species complex derived from perithecia and soil in the old world tropics. Mycologia 2011, 103, 1302–1330. [Google Scholar] [CrossRef]
- Aoki, T.; O’Donnell, K.; Homma, Y.; Lattanzi, A.R. Sudden-death syndrome of soybean is caused by two morphologically and phylogenetically distinct species within the Fusarium solani species complex-F. virguliforme in North America and F. tucumaniae in South America. Mycologia 2003, 95, 660–684. [Google Scholar] [CrossRef]
- Smith, E.F. Wilt disease of cotton, watermelon and cowpea (Neocosmospora nov. gen.). US. Dep. Agric. Bull. 1899, 17, 1–53. [Google Scholar]
- Mahoney, D.P. A new Neocosmospora from Galapagos Island soil. Mycologia 1976, 68, 1111–1116. [Google Scholar] [CrossRef]
- Cannon, P.F.; Hawksworth, L. A revision of the genus Neocosmospora (Hypocreales). Trans. Br. Mycol. Soc. 1984, 82, 673–688. [Google Scholar] [CrossRef]
- Udagawa, S.I.; Horie, Y.; Cannon, P.F. Two new species of Neocosmospora from Japan, with a key to the currently accepted species. Sydowia 1989, 41, 349–359. [Google Scholar]
- Pfenning, L. A new species of Neocosmospora from Brazil. Sydowia 1995, 47, 65–69. [Google Scholar]
- Gräfenhan, T.; Schroers, H.J.; Nirenberg, H.I.; Seifert, K.A. An overview of the taxonomy, phylogeny, and typification of nectriaceous fungi in Cosmospora, Acremonium, Fusarium, Stilbella, and Volutella. Stud. Mycol. 2011, 68, 79–113. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.T.; Fu, X.H.; Chen, Y.D. Two petroliphilous taxa of Fusarium. Acta Mycol. Sin. Suppl. 1987, 1, 328–333. [Google Scholar]
- Luo, J.; Zhuang, W.Y. Four new species and a new Chinese record of the nectrioid fungi. Sci. China Life Sci. 2010, 53, 909–915. [Google Scholar] [CrossRef]
- Zeng, Z.Q.; Zhuang, W.Y. Two new species of Neocosmospora from China. Phytotaxa 2017, 319, 175–183. [Google Scholar] [CrossRef]
- Zeng, Z.Q.; Zhuang, W.Y. New species of Nectriaceae (Hypocreales) from China. J. Fungi 2022, 8, 1075. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain/Herbarium Numbers | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| CAM | ITS | RPB2 | TEF1 | ||
| Geejayessia atrofusca | NRRL 22316 | - | AF178423 | JX171609 | AF178361 |
| Geejayessia cicatricum | CBS 125552 | - | MH863560 | HQ728153 | HM626644 |
| Neocosmospora acutispora | NRRL 22574 | MW834122 | NR169884 | EU329514 | AF178345 |
| Neocosmospora ambrosia | NRRL 22346 | - | EU329669 | EU329503 | FJ240350 |
| Neocosmospora anhuiensis | CGMCC 3.24869 | OR014310 a | OQ842733 | OQ866525 | OQ866530 |
| Neocosmospora aurantia | CGMCC 3.24866 | OR014308 | OQ842731 | OQ866523 | OQ866528 |
| Neocosmospora caricae | ES216M | - | OK422518 | OK415859 | OK539518 |
| Neocosmospora cryptoseptata | NRRL 22412 | MW834126 | NR172368 | EU329510 | AF178351 |
| Neocosmospora cyanescens | CBS 51882 | MW218064 | AB190389 | LR583826 | LR583605 |
| Neocosmospora dimorpha | CGMCC 3.24867 | OR014309 | OQ842732 | OQ866524 | OQ866529 |
| Neocosmospora euwallaceae | NRRL 54722 | KU171422 | JQ038014 | JQ038028 | JQ038007 |
| Neocosmospora ferruginea | NRRL 32437 | MW834132 | DQ094446 | EU329581 | DQ246979 |
| Neocosmospora galbana | CGMCC 3.24868 | OR014307 | OQ842730 | OQ866527 | OQ866532 |
| Neocosmospora keleraja | CBS 125720 | MW834138 | LR583720 | LR583834 | LR583612 |
| Neocosmospora kuroshio | CBS 142642 | MW834140 | LR583723 | LR583837 | KX262216 |
| Neocosmospora kurunegalensis | CBS 119599 | MW834141 | JF433036 | LR583838 | DQ247511 |
| Neocosmospora lithocarpi | LC 1113 | - | MW016711 | MW474697 | MW620172 |
| Neocosmospora longissima | CBS 126407 | MW834144 | NR178144 | LR583846 | LR583621 |
| Neocosmospora macrospora | CPC 28191 | MW218078 | NR163291 | LT746331 | LT746218 |
| Neocosmospora mahasenii | CBS 119594 | MW834145 | JF433045 | LT960563 | DQ247513 |
| Neocosmospora maoershanica | CGMCC 3.24870 | OR014311 | OQ842734 | OQ866526 | OQ866531 |
| Neocosmospora mori | NRRL 22230 | MW834149 | AF178420 | EU329499 | AF178358 |
| Neocosmospora nelsonii | CBS 30975 | MW834152 | MW827630 | MW847904 | MW847907 |
| Neocosmospora nirenbergiana | NRRL 22387 | MW834153 | NR169883 | EU329505 | AF178339 |
| Neocosmospora oblonga | CBS 130325 | MW834154 | LR583746 | LR583853 | LR583631 |
| Neocosmospora oligoseptata | NRRL 62579 | MW834155 | KC691566 | LR583854 | KC691538 |
| Neocosmospora phaseoli | CBS 26550 | KM231380 | MH856617 | KM232375 | HE647964 |
| Neocosmospora pisi | CBS 123669 | MW834159 | KM231796 | KM232364 | KM231925 |
| Neocosmospora pseudensiformis | CBS 130.78 | MW834162 | LR583759 | LR583868 | DQ247635 |
| Neocosmospora quercicola | CBS 14190 | MW834164 | NR178125 | LR583869 | DQ247634 |
| Neocosmospora regularis | CBS 23034 | MW834168 | LR583763 | LR583873 | LR583643 |
| Neocosmospora rekana | CMW 52862 | - | MN249094 | MN249137 | MN249151 |
| Neocosmospora robusta | NRRL 22395 | MW834169 | NR172367 | EU329507 | AF178341 |
| Neocosmospora samuelsii | CBS 114067 | MW834170 | NR178127 | LR583874 | LR583644 |
| Neocosmospora silvicola | CBS 123846 | MW834172 | LR583766 | LR583876 | LR583646 |
| Neocosmospora spathulata | NRRL 28541 | MW218091 | EU329674 | EU329542 | DQ246882 |
| Neocosmospora vasinfecta | CBS 446.93 | MW834175 | LR583791 | LR583898 | LR583670 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Z.-Q.; Zhuang, W.-Y. New Species of Neocosmospora (Ascomycota) from China as Evidenced by Morphological and Molecular Data. Life 2023, 13, 1515. https://doi.org/10.3390/life13071515
Zeng Z-Q, Zhuang W-Y. New Species of Neocosmospora (Ascomycota) from China as Evidenced by Morphological and Molecular Data. Life. 2023; 13(7):1515. https://doi.org/10.3390/life13071515
Chicago/Turabian StyleZeng, Zhao-Qing, and Wen-Ying Zhuang. 2023. "New Species of Neocosmospora (Ascomycota) from China as Evidenced by Morphological and Molecular Data" Life 13, no. 7: 1515. https://doi.org/10.3390/life13071515