
In Vitro Antioxidant and Fibroblast Migration Activities of Fractions Eluded from Dichloromethane Leaf Extract of Marantodes pumilum

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Preparation of Plant Material
2.3. Column Chromatography Purification
2.4. Antioxidant Studies
2.4.1. Total Antioxidant Capacity (TAC)
2.4.2. Total Proanthocyanidins (TPA)
2.5. Free Radical Scavenging Studies
2.5.1. 2,2-Diphenylpicrylhydrazyl (DPPH) Assay
2.5.2. Nitric Oxide (NO) Assay
2.5.3. Hydroxyl Radical Inhibition Assay
2.5.4. Superoxide Radical Inhibition Assay
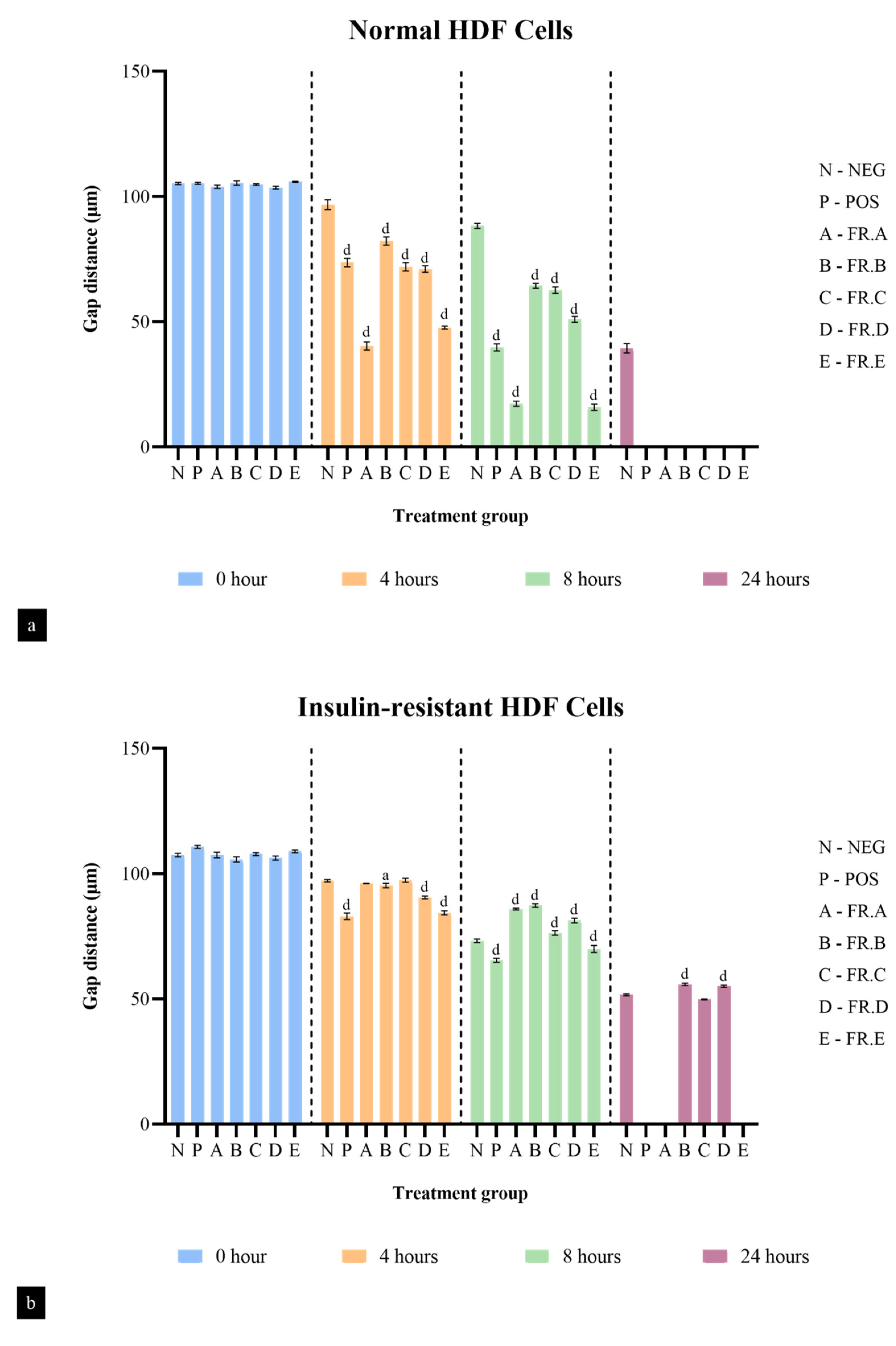
2.6. Fibroblast Cell Migration Studies
2.6.1. HDF Cell Culture and Differentiation
2.6.2. Induction of Insulin Resistance
2.6.3. Scratch Wound Assay
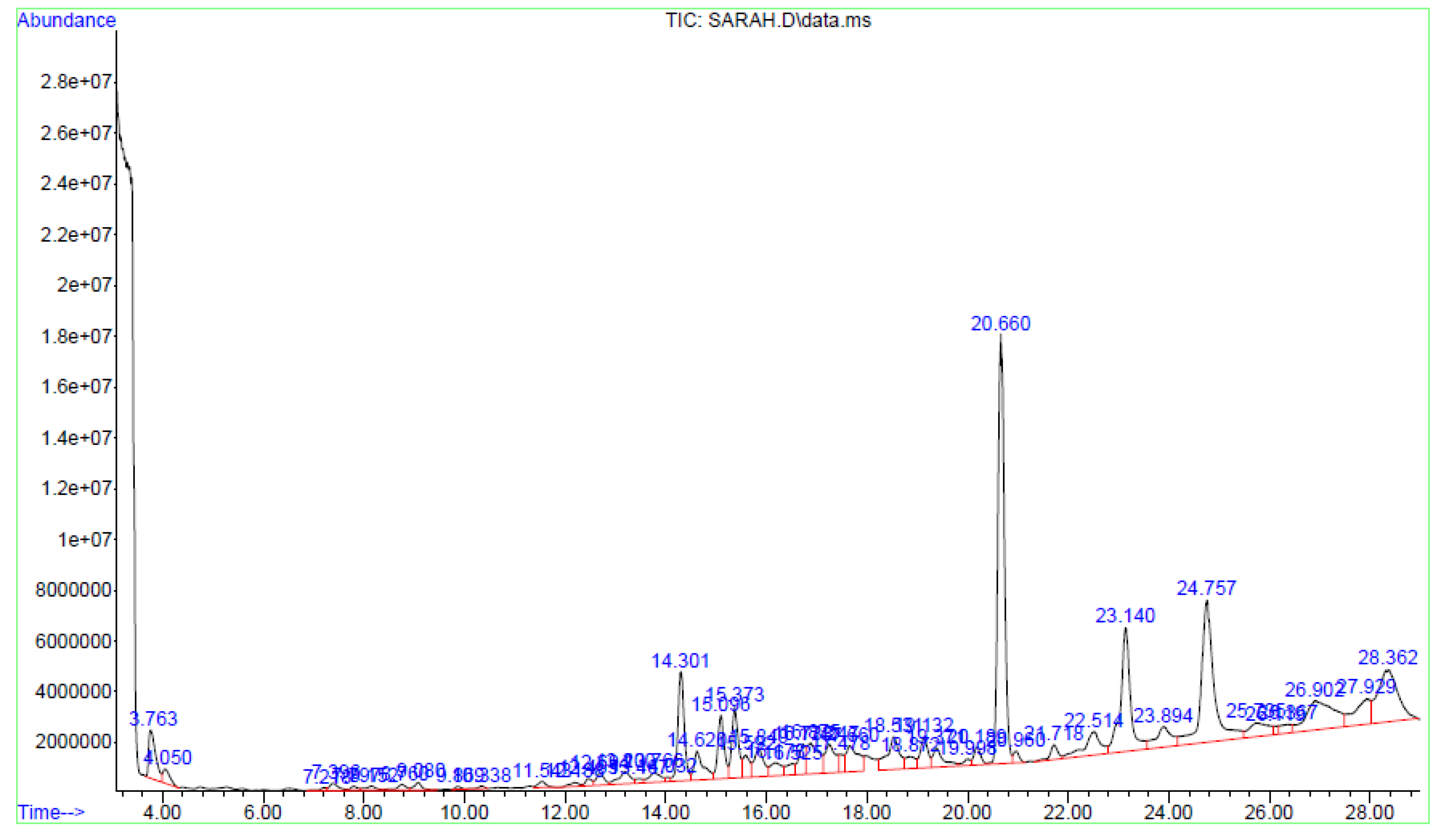
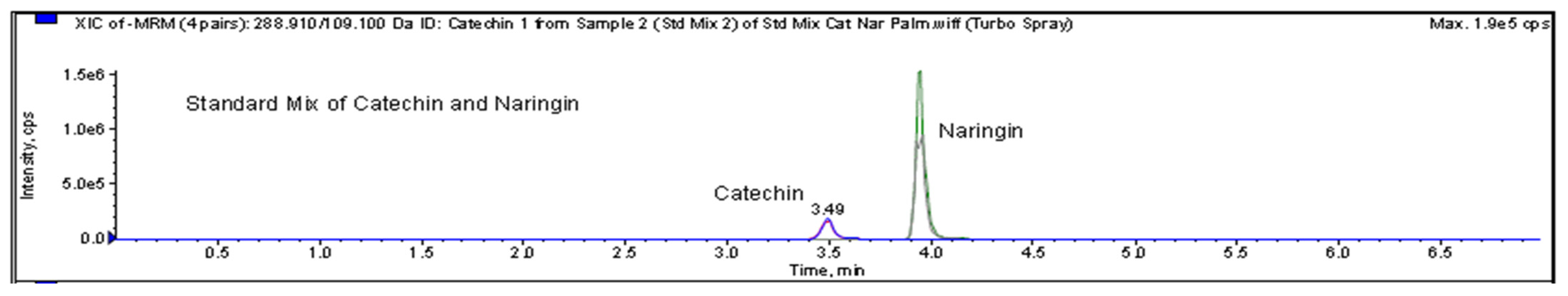
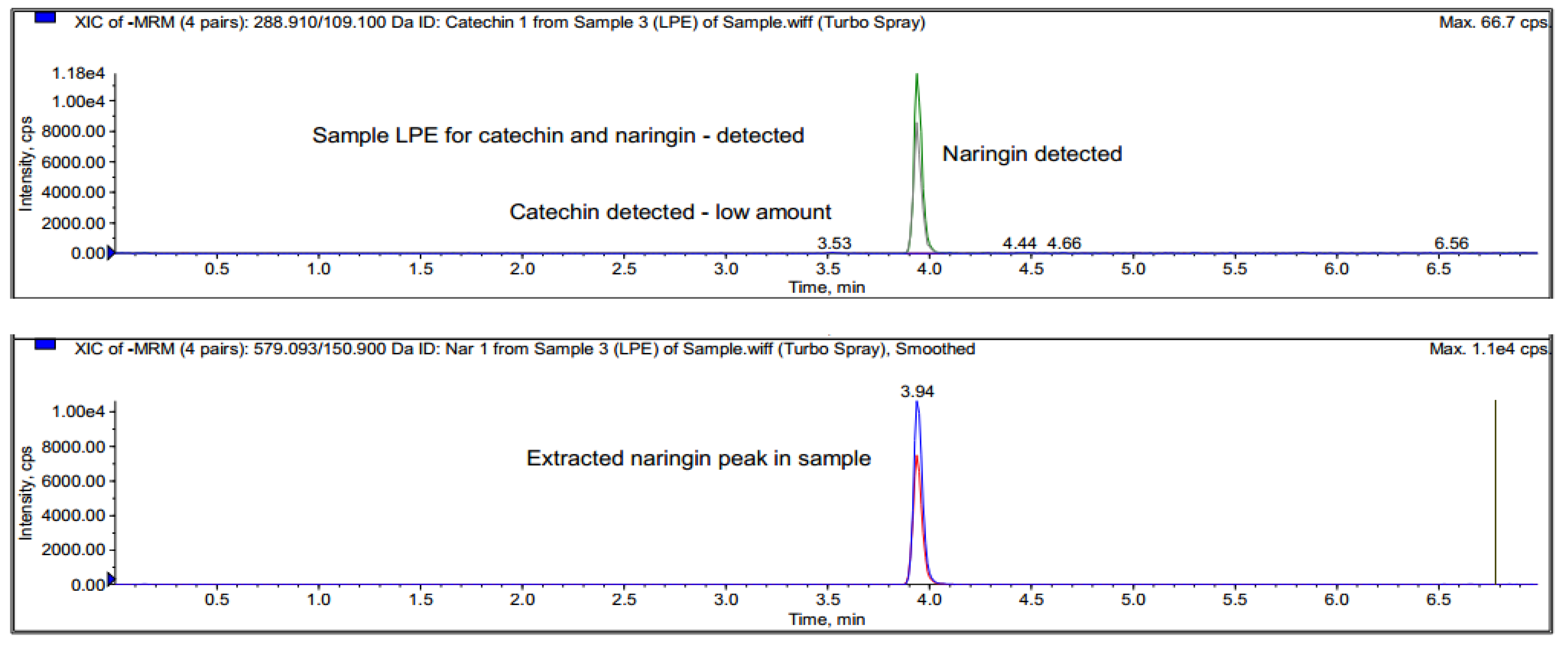
2.7. Identification of Chemical Constituents in Fractions A and E
2.7.1. GC-MS of Fraction A
2.7.2. HPTLC of Fraction E
2.7.3. LC-MS/MRM of Fraction E
2.8. Statistical Analysis
3. Results
3.1. Column Chromatography Purification
3.2. Antioxidant Studies
Free Radical Scavenging Studies
3.3. Fibroblast Cell Migration Studies
3.4. Identification of Major Compounds Present in Fractions A and E
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patel, S.; Srivastava, S.; Singh, M.R.; Singh, D. Mechanistic insight into diabetic wounds: Pathogenesis, molecular targets and treatment strategies to pace wound healing. Biomed. Pharm. 2019, 112, 108615. [Google Scholar] [CrossRef] [PubMed]
- Okonkwo, U.A.; Chen, L.; Ma, D.; Haywood, V.A.; Barakat, M.; Urao, N.; DiPietro, L.A. Compromised angiogenesis and vascular integrity in impaired diabetic wound healing. PLoS ONE 2020, 15, e0231962. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Cheng, Y.; Ye, J.; Cai, P.; Zhang, J.; Li, R.; Yang, Y.; Wang, Z.; Zhang, H.; Lin, C.; et al. bFGF Promotes the Migration of Human Dermal Fibroblasts under Diabetic Conditions through Reactive Oxygen Species Production via the PI3K/Akt-Rac1- JNK Pathways. Int. J. Biol. Sci. 2015, 11, 845–859. [Google Scholar] [CrossRef]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef]
- de Oliveira Gonzalez, A.C.; Costa, T.F.; De Araújo Andrade, Z.; Medrado, A.R.A.P. Wound healing—A literature review. Bras. Derm. 2016, 91, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Cen, R.; Wang, L.; He, Y.; Yue, C.; Tan, Y.; Li, L.; Lei, X. Dermal Fibroblast Migration and Proliferation Upon Wounding or Lipopolysaccharide Exposure is Mediated by Stathmin. Front. Pharm. 2022, 12, 781282. [Google Scholar] [CrossRef]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive oxygen species (ROS) and wound healing: The functional role of ROS and emerging ROS-modulating technologies for augmentation of the healing process: Reactive oxygen species and wound healing. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef]
- Carbone, F.; Bonaventura, A.; Montecucco, F. Neutrophil-Related Oxidants Drive Heart and Brain Remodeling After Ischemia/Reperfusion Injury. Front. Physiol. 2020, 10, 1587. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, M.; Kumar, N.V.A.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Fokou, P.V.T.; Azzini, E.; Peluso, E.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.A.; Niharika, P.; Reddy, H.; Reddy, N.V.; Manoj Kumar, M.G.; Pranitha, V. Antioxidant mix: A novel pulpotomy medicament: A scanning electron microscopy evaluation. Contemp. Clin. Dent. 2014, 5, 428–433. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Barku, V.Y.A. Wound Healing: Contributions from Plant Secondary Metabolite Antioxidants. In Wound Healing—Current Perspectives; Dogan, K.H., Ed.; IntechOpen: London, UK, 2019; pp. 49–63. [Google Scholar]
- Okechukwu, P.N.; Ekeuku, S.O.; Loshnie, S.; Akowuah, G.A. Anti-inflammatory, Analgesic, Antinociceptive and Antipyretic Investigation of Bioactive Constituents from Partial Purified Dichloromethane Crude Extracts from Leaves of Labisia pumila. IJPRS 2014, 3, 742–751. [Google Scholar]
- Okechukwu, P.N. Evaluation of anti-inflammatory, analgesic, antipyretic effect of Eicosane, Pentadecane, Octacosane, and Heneicosane. Asian J. Pharm. Clin. Res. 2020, 13, 29–35. [Google Scholar] [CrossRef]
- Samuagam, L.; Akowuah, G.A.; Okechukwu, P.N. Partial Purification and Antinociceptive Investigation of Extracts of Leaves of Labisia pumila. Asian J. Pharm. Clin. Res. 2011, 4, 44–46. [Google Scholar]
- Chuah, X.Q.; Okechukwu, P.N.; Amini, F.; Teo, S.S. Eicosane, pentadecane and palmitic acid: The effects in in vitro wound healing studies. Asian Pac. J. Trop. Biomed. 2018, 8, 490–499. [Google Scholar]
- Siyumbwa, S.N.; Ekeuku, S.O.; Amini, F.; Emerald, N.M.; Sharma, D.; Okechukwu, P.N. Wound Healing and Anti-bacterial Activities of 2-Pentadecanone in STZ-Induced Type 2 Diabetic Rats. Phcog. Mag. 2019, 15, 71–77. [Google Scholar]
- Khan, R.A.; Khan, M.R.; Sahreen, S.; Ahmed, M. Assessment of flavonoids contents and in vitro antioxidant activity of Launaea procumbens. Chem. Cent. J. 2012, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Tsuji, S.; Tonogai, Y. Analysis of Proanthocyanidins in Grape Seed Extracts, Health Foods and Grape Seed Oils. J. Health Sci. 2003, 49, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Lee, S.; Lee, Y.S.; Jung, S.H.; Park, Y.; Shin, K.H.; Kim, B.K. Anti-oxidant activities of the extracts from the herbs of Artemisia apiacea. J. Ethnopharmacol. 2003, 85, 69–72. [Google Scholar] [CrossRef]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Madan, S.; Singh, G.N.; Kumar, Y.; Kohli, K. Phytochemical analysis and free-radical scavenging activity of Flemingia strobilifera (Linn) R. Br. Res. J. Pharm. Biol. Chem. Sci. 2010, 1, 183–190. [Google Scholar]
- Sandesh, P.; Velu, V.; Singh, R.P. Antioxidant activities of tamarind (Tamarindus indica) seed coat extracts using in vitro and in vivo models. J. Food Sci. Technol. 2014, 51, 1965–1973. [Google Scholar] [CrossRef] [Green Version]
- Aarti, R.; Sushil, B.; Tenpe, C.R.; Yeole, P.G. Free radical scavenging activity of polyherbal formulation. Pharmacologyonline 2009, 3, 944–951. [Google Scholar]
- Jakovljevic, N.K.; Pavlovic, K.; Zujovic, T.; Kravic-Stevovic, T.; Jotic, A.; Markovic, I.; Lalic, N.M. In vitro models of insulin resistance: Mitochondrial coupling is differently affected in liver and muscle cells. Mitochondrion 2021, 61, 165–173. [Google Scholar] [CrossRef]
- Feraco, A.; Gorini, S.; Armani, A.; Camajani, E.; Rizzo, M.; Caprio, M. Exploring the Role of Skeletal Muscle in Insulin Resistance: Lessons from Cultured Cells to Animal Models. Int. J. Mol. Sci. 2021, 22, 9327. [Google Scholar] [CrossRef]
- D’Urso, M.; Kurniawan, N.A. Mechanical and Physical Regulation of Fibroblast-Myofibroblast Transition: From Cellular Mechanoresponse to Tissue Pathology. Front. Bioeng. Biotechnol. 2020, 8, 609653. [Google Scholar] [CrossRef]
- Addis, R.; Cruciani, S.; Santaniello, S.; Bellu, E.; Sarais, G.; Ventura, C.; Maioli, M.; Pintore, G. Fibroblast Proliferation and Migration in Wound Healing by Phytochemicals: Evidence for a Novel Synergic Outcome. Int. J. Med. Sci. 2020, 17, 1030–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimeno, A.; García-Giménez, J.L.; Audí, L.; Toran, N.; Andaluz, P.; Dasí, F.; Viña, J.; Pallardó, F.V. Decreased cell proliferation and higher oxidative stress in fibroblasts from Down Syndrome fetuses. Preliminary study. Biochim. Biophys. Acta 2014, 1842, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, J.L.; Wyant, W.A.; Abujamra, B.A.; Kirsner, R.S.; Jozic, I. Diabetic Wound-Healing Science. Medicina 2021, 57, 1072. [Google Scholar] [CrossRef]
- Li, W.; Terauchi, N.; Meng, D.; Miyamoto, N.; Tsutsumi, N.; Ura, K.; Takagi, Y. Antioxidant and fibroblast-activating activities of the by-product of skate chondroitin extractive production. Sustain. Chem. Pharm. 2021, 23, 100499. [Google Scholar] [CrossRef]
- Sarkar, P.; Stefi, R.V.; Pasupuleti, M.; Paray, B.A.; Al-Sadoon, M.K.; Arockiaraj, J. Antioxidant molecular mechanism of adenosyl homocysteinase from cyanobacteria and its wound healing process in fibroblast cells. Mol. Biol. Rep. 2020, 47, 1821–1834. [Google Scholar] [CrossRef]
- Boakye, Y.D.; Agyare, C.; Ayande, G.P.; Titiloye, N.; Asiamah, E.A.; Danquah, K.O. Assessment of Wound-Healing Properties of Medicinal Plants: The Case of Phyllanthus muellarianus. Front. Pharm. 2018, 9, 945. [Google Scholar] [CrossRef]
- Krishnan, N.; Velramar, B.; Ramatchandirin, B.; Abraham, G.C.; Duraisamy, N.; Pandiyan, R.; Velu, R.K. Effect of biogenic silver nanocubes on matrix metalloproteinases 2 and 9 expressions in hyperglycemic skin injury and its impact in early wound healing in streptozotocin-induced diabetic mice. Mater. Sci. Eng. 2018, 91, 146–152. [Google Scholar] [CrossRef]
- Dickinson, L.E.; Gerecht, S. Engineered Biopolymeric Scaffolds for Chronic Wound Healing. Front. Physiol. 2016, 7, 341. [Google Scholar] [CrossRef] [Green Version]
- Honnegowda, T.M.; Kumar, P.; Udupa, E.G.P.; Kumar, S.; Kumar, U.; Rao, P. Role of angiogenesis and angiogenic factors in acute and chronic wound healing. Plast. Aesthetic Res. 2015, 2, 243–249. [Google Scholar]
- Aleem, M.; Maqsood, H.; Younus, S.; Zafar, A.F.; Talpur, A.S.; Shakeel, H. Fibroblast Growth Factor 21 and Its Association With Oxidative Stress and Lipid Profile in Type 2 Diabetes Mellitus. Cureus 2021, 13, e17723. [Google Scholar] [CrossRef] [PubMed]
- Bagdas, D.; Etoz, B.C.; Gul, Z.; Ziyanok, S.; Inan, S.; Turacozen, O.; Gul, N.Y.; Topal, A.; Cinkilic, N.; Tas, S.; et al. In vivo systemic chlorogenic acid therapy under diabetic conditions: Wound healing effect and cytotoxicity/gentoxicity profile. Food Chem. Toxicol. 2015, 81, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Süntar, I.; Akkol, E.K.; Nahar, L.; Sarker, S.D. Wound healing and antioxidant properties: Do they coexist in plants? Free Rad. Antiox. 2012, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Balachandran, A.; Choi, S.B.; Beata, M.M.; Małgorzata, J.; Froemming, G.R.A.; Lavilla, C.A.; Billacura, M.P., Jr.; Siyumbwa, S.N.; Okechukwu, P.N. Antioxidant, Wound Healing Potential and In Silico Assessment of Naringin, Eicosane and Octacosane. Molecules 2023, 28, 1043. [Google Scholar] [CrossRef]
- Emran, T.B.; Islam, F.; Nath, N.; Sutradhar, H.; Das, R.; Mitra, S.; Alshahrani, M.M.; Alhasaniah, A.H.; Sharma, R. Naringin and Naringenin Polyphenols in Neurological Diseases: Understandings from a Therapeutic Viewpoint. Life 2022, 13, 99. [Google Scholar] [CrossRef]
- Vijayaragahvan, K.; Rajkumar, J.; Bukhari, S.N.A.; Al-Sayed, B.; Seyed, M.A. Chromolaena odorata: A neglected weed with a wide spectrum of pharmacological activities (Review). Mol. Med. Rep. 2017, 15, 1007–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Diao, Y.; Zhang, H.; Wang, S.; Zhang, Z.; Yu, B.; Huang, S.; Yang, H. Tannin extracts from immature fruits of Terminalia chebula Fructus Retz. promote cutaneous wound healing in rats. BMC Complement. Altern. Med. 2011, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Giovannelli, L.; Pitozzi, V.; Jacomelli, M.; Mulinacci, N.; Laurenzana, A.; Dolara, P.; Mocali, A. Protective Effects of Resveratrol Against Senescence-Associated Changes in Cultured Human Fibroblasts. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66A, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaisas, M.M.; Kshirsagar, S.B.; Sahane, R.S. Evaluation of wound healing activity of ferulic acid in diabetic rats. Int. Wound J. 2014, 11, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Afshar, M.; Hassanzadeh-Taheri, M.M.; Zardast, M.; Moghaddam, A. The Angiogenetic Effect of Resveratrol on Dermal Wound Healing in Balb/C Mice. Mod. Care J. 2017, 14, e66118. [Google Scholar] [CrossRef]
- Gu, X.; Luo, X.; Wang, Y.; He, Z.; Li, X.; Wu, K.; Zhang, Y.; Yang, Y.; Ji, J.; Luo, X. Ascorbic acid attenuates cell stress by activating the fibroblast growth factor 21/fibroblast growth factor receptor 2/adiponectin pathway in HepG2 cells. Mol. Med. Rep. 2019, 20, 2450–2458. [Google Scholar] [CrossRef]
- Wu, Q.J.; Zhang, T.N.; Chen, H.H.; Yu, X.F.; Lv, J.L.; Liu, Y.Y.; Liu, Y.S.; Zheng, G.; Zhao, J.Q.; Wei, Y.F.; et al. The sirtuin family in health and disease. Signal Transduct. Target. Ther. 2022, 7, 402. [Google Scholar] [CrossRef]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [Green Version]
- Villalba, J.M.; Alcaín, F.J. Sirtuin activators and inhibitors. Biofactors 2012, 38, 349–359. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, J.; Wang, J.; Liu, Q. The controversial links among calorie restriction, SIRT1, and resveratrol. Free. Radic. Biol. Med. 2011, 51, 250–256. [Google Scholar] [CrossRef]
- Chondrogianni, N.; Kapeta, S.; Chinou, I.; Vassilatou, K.; Papassideri, I.; Gonos, E.S. Anti-ageing and rejuvenating effects of quercetin. Exp. Gerontol. 2010, 45, 763–771. [Google Scholar]
- Okada, Y.; Okada, M. Quercetin, caffeic acid and resveratrol regulate circadian clock genes and aging-related genes in young and old human lung fibroblast cells. Mol. Biol. Rep. 2020, 47, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Velalar, C.N.; Ruan, R. Effects of epigallocatechin-3-gallate on mitochondrial integrity and antioxidative enzyme activity in the aging process of human fibroblast. Free. Radic. Biol. Med. 2008, 44, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Kim, B.K.; Park, K.Y.; Yokozawa, T.; Song, Y.O.; Cho, E.J. Anti-aging effects of cyanidin under a stress-induced premature senescence cellular system. Biol. Pharm. Bull. 2010, 33, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.R.; Choi, M.J.; Choi, J.M.; Ko, J.C.; Ko, J.Y.; Cho, E.J. Malvidin Protects WI-38 Human Fibroblast Cells Against Stress-induced Premature Senescence. J. Cancer Prev. 2016, 21, 32–40. [Google Scholar] [CrossRef]
- Liu, C.F.; Li, X.L.; Zhang, Z.L.; Qiu, L.; Ding, S.X.; Xue, J.X.; Zhao, G.P.; Li, J. Antiaging Effects of Urolithin A on Replicative Senescent Human Skin Fibroblasts. Rejuvenation Res. 2019, 22, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Torres-Vega, J.; Gómez-Alonso, S.; Pérez-Navarro, J.; Alarcón-Enos, J.; Pastene-Navarrete, E. Polyphenolic Compounds Extracted and Purified from Buddleja globosa Hope (Buddlejaceae) Leaves Using Natural Deep Eutectic Solvents and Centrifugal Partition Chromatography. Molecules 2021, 26, 2192. [Google Scholar] [CrossRef]
- Singh, M.; Govindarajan, R.; Nath, V.; Rawat, A.K.S.; Mehrotra, S. Antimicrobial, wound healing and antioxidant activity of Plagiochasma appendiculatum Lehm. et Lind. J. Ethnopharmacol. 2006, 107, 67–72. [Google Scholar] [CrossRef]
- Adetutu, A.; Morgan, W.A.; Corcoran, O. Antibacterial, antioxidant and fibroblast growth stimulation activity of crude extracts of Bridelia ferruginea leaf, a wound-healing plant of Nigeria. J. Ethnopharmacol. 2011, 133, 116–119. [Google Scholar] [CrossRef]
- Das, B.; Pal, D.; Haldar, A. A Review on Biological Activities and Medicinal Properties Of Clerodendrum infortunatum Linn. Int. J. Pharm. Pharm. Sci. 2014, 6, 41–43. [Google Scholar]
- Sereme, B.; Umar, F.Y.; Okechukwu, P.N. Anti-inflammatory, analgesic and antipyretic activity of cassava leaf extract. Asian J. Pharm. Clin. Res. 2013, 6, 89–92. [Google Scholar]
- Anwar, U.; Mohd Bohari, S.P. Effect Of Manihot Esculenta Aqueous Extract and Therapeutic Ultrasound in Accelerating The Wound Healing Process In Vitro. J. Teknol. 2019, 81, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Bahekar, S.E.; Kale, R.S. Evaluation of antioxidant activity of Manihot esculenta Crantz in wistar rats. J. Pharm. Bioallied. Sci. 2016, 8, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Noumedem, J.A.K.; Mihasan, M.; Lacmata, S.T.; Stefan, M.; Kuiate, J.R.; Kuete, V. Antibacterial activities of the methanol extracts of ten Cameroonian vegetables against Gram-negative multidrug-resistant bacteria. BMC Complement. Altern. Med. 2013, 13, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, B.; Hu, L.; Mei, W.; Zhou, K.; Wang, H.; Luo, Y.; Wei, X.; Dai, H. Antioxidant Phenolic Compounds of Cassava (Manihot esculenta) from Hainan. Molecules 2010, 16, 10157–10167. [Google Scholar] [CrossRef] [Green Version]
- Isenmann, E.; Trittel, L.; Diel, P. The effect of alpha lipoic acid on muscle strength recovery after a single and a short-term chronic supplementation—A study in healthy well-trained individuals after intensive resistance and endurance training. J. Int. Soc. Sport. Nutr. 2020, 17, 61. [Google Scholar] [CrossRef]
- Nasole, E.; Nicoletti, C.; Yang, Z.J.; Girelli, A.; Rubini, A.; Giuffreda, F.; Tano, A.D.; Camporesi, E.; Bosco, G. Effects of alpha lipoic acid and its R+ enantiomer supplemented to hyperbaric oxygen therapy on interleukin-6, TNF- α and EGF production in chronic leg wound healing. J. Enzym. Inhib. Med. Chem. 2014, 29, 297–302. [Google Scholar] [CrossRef]
- Zhao, H.; Huang, J.; Li, Y.; Lv, X.; Zhou, H.; Wang, H.; Xu, Y.; Wang, C.; Wang, J.; Liu, Z. ROS-scavenging hydrogel to promote healing of bacteria infected diabetic wounds. Biomaterials 2020, 258, 120286. [Google Scholar] [CrossRef]
- Kharaghani, D.; Gitigard, P.; Ohtani, H.; Kim, K.O.; Ullah, S.; Saito, Y.; Khan, M.Q.; Kim, I.S. Design and characterization of dual drug delivery based on in-situ assembled PVA/PAN core-shell nanofibers for wound dressing application. Sci. Rep. 2019, 9, 12640. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.C.; Chuang, F.S.; Venkatesan, M.; Cho, C.J.; Chen, P.Y.; Tzeng, Y.R.; Yu, Y.Y.; Rwei, S.P.; Kuo, C.C. Synthesis of Water Resistance and Moisture-Permeable Nanofiber Using Sodium Alginate-Functionalized Waterborne Polyurethane. Polymers 2020, 12, 2882. [Google Scholar] [CrossRef]
- Lee, C.H.; Huang, S.C.; Hung, K.C.; Cho, C.J.; Liu, S.J. Enhanced Diabetic Wound Healing Using Electrospun Biocompatible PLGA-Based Saxagliptin Fibrous Membranes. Nanomaterials 2022, 12, 3740. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions | Colour of Eluent | Dry Weight (g) | Yield Percentage (%) |
|---|---|---|---|
| Fraction A | Yellow | 0.878 ± 0.05 | 10.99 ± 0.03 |
| Fraction B | Dark green | 1.006 ± 0.02 | 12.60 ± 0.05 |
| Fraction C | Green | 0.548 ± 0.03 | 6.86 ± 0.08 |
| Fraction D | Orange | 0.760 ± 0.03 | 9.52 ± 0.04 |
| Fraction E | Orange | 0.554 ± 0.01 | 6.94 ± 0.01 |
| Fractions | TAC (mg AAE/g) | TPA (mg CAT/g) |
|---|---|---|
| Fraction A | 18.2 ± 5.29 | 43.9 ± 1.04 a |
| Fraction B | 10.7 ± 1.16 bc | 23.4 ± 3.41 a |
| Fraction C | 16.0 ± 6.93 b | 25.3 ± 1.00 a |
| Fraction D | 24.0 ± 1.00 | 42.5 ± 0.46 a |
| Fraction E | 30.2 ± 2.00 | 59.3 ± 1.06 |
| Assays | EC50 (µg/mL) | |||||
|---|---|---|---|---|---|---|
| Standard | Fraction A | Fraction B | Fraction C | Fraction D | Fraction E | |
| DPPH | 0.05 ± 0.02 | 27.21 ± 4.48 | 266.53 ± 51.16 c | 72.50 ± 11.08 d | 4.63 ± 3.63 | 2.88 ± 0.95 |
| NO | 328.9 ± 24.72 | 464.0 ± 5.27 d | 406.9 ± 5.48 d | 314.4 ± 6.69 | 195.7 ± 12.34 d | 109.8 ± 3.69 d |
| OH¯ | 54.98 ± 5.47 | 239.3 ± 6.87 d | 398.5 ± 9.6 d | 289.3 ± 7.91 d | 198.6 ± 8.07 c | 193.3 ± 3.88 b |
| SO¯ | 103.9 ± 0.94 | 162.7 ± 10.7 a | 703.0 ± 10.9 d | 561.7 ± 53.6 d | 260.3 ± 15.2 d | 413.1 ± 7.84 d |
| No | Compounds | Molecular Weight (MW) | Percentage Amount (PA) | Retention Time (mins) | |
|---|---|---|---|---|---|
| 1. |  | 2,3,3-trimethyl 1-butene | 98.186 | 2.16 | 3.762 |
 | 4,4-dimethyl-(Z)-2-pentene | ||||
| 2. |  | 6,10,14-trimethyl-2-pentadecanone | 268.478 | 4.46 | 14.302 |
| 3. |  | 1-octacosanol | 506.768 | 2.61 | 15.097 |
 | 1-docosene | 308.585 | |||
 | 9-nonadecene | 266.505 | |||
| 4. |  | Isophytol | 296.351 | 2.96 | 15.372 |
| 5. |  | Tetracosyl heptafluorobutyrate | 550.676 | 2.38 | 16.877 |
| 6. |  | Ethanol, 2-(octadecyloxy)- | 314.546 | 2.03 | 17.661 |
 | 1-(4-bromobutyl)-2-piperidinone | 234.133 | |||
 | 1-decanol, 2-hexyl- | 242.441 | |||
| 7. |  | Tricosane | 324.627 | 2.56 | 18.530 |
| 8. |  | Bis(2-ethylhexyl) phthalate | 390.556 | 17.80 | 20.659 |
 | Bis(2-propylpentyl) phthalate | ||||
| 9. |  | i-propyl 9-octadecenoate | 324.541 | 2.46 | 22.513 |
 | 9-octadecenoic acid | 282.461 | |||
| 10. |  | Pentadecane | 212.415 | 7.89 | 23.142 |
 | Heptadecane, 9-octyl- | 352.680 | |||
 | Heptacosane, 1-chloro- | 415.179 | |||
| 11. |  | Heneicosane | 296.574 | 11.72 | 24.756 |
 | Hexadecane, 1-iodo- | 352.338 | |||
 | Octacosane | 394.760 | |||
| 12. |  | Eicosane | 282.548 | 5.25 | 26.902 |
| 13. |  | 1,3-dithiolane-2-(28-norurs-12-en-17-yl)- | 500.885 | 2.65 | 27.932 |
 | 3,7,11-trimethyl-dodeca-2,4,6,10-tetraenal | 218.335 | |||
 | (22E)-3’,7β-dihydrocycloprop[7,8]-5α-ergost-22-en-3-one | NR | |||
| 14. |  | β-amyrin | 426.717 | 6.84 | 28.361 |
 | Urs-12-en-3-one | 424.702 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balachandran, A.; Siyumbwa, S.N.; Froemming, G.R.A.; Beata, M.-M.; Małgorzata, J.; Lavilla, C.A., Jr.; Billacura, M.P.; Okechukwu, P.N. In Vitro Antioxidant and Fibroblast Migration Activities of Fractions Eluded from Dichloromethane Leaf Extract of Marantodes pumilum. Life 2023, 13, 1409. https://doi.org/10.3390/life13061409
Balachandran A, Siyumbwa SN, Froemming GRA, Beata M-M, Małgorzata J, Lavilla CA Jr., Billacura MP, Okechukwu PN. In Vitro Antioxidant and Fibroblast Migration Activities of Fractions Eluded from Dichloromethane Leaf Extract of Marantodes pumilum. Life. 2023; 13(6):1409. https://doi.org/10.3390/life13061409
Chicago/Turabian StyleBalachandran, Abbirami, Stepfanie N. Siyumbwa, Gabriele R. A. Froemming, Morak-Młodawska Beata, Jeleń Małgorzata, Charlie A. Lavilla, Jr., Merell P. Billacura, and Patrick N. Okechukwu. 2023. "In Vitro Antioxidant and Fibroblast Migration Activities of Fractions Eluded from Dichloromethane Leaf Extract of Marantodes pumilum" Life 13, no. 6: 1409. https://doi.org/10.3390/life13061409