

Screening and Selection of Drought-Tolerant High-Yielding Chickpea Genotypes Based on Physio-Biochemical Selection Indices and Yield Trials

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Physiological Traits
2.2. Biochemical Traits
2.3. Morpho-Phenological Traits, Yield and Yield Attributing Traits
2.4. Statistical Analysis
3. Results
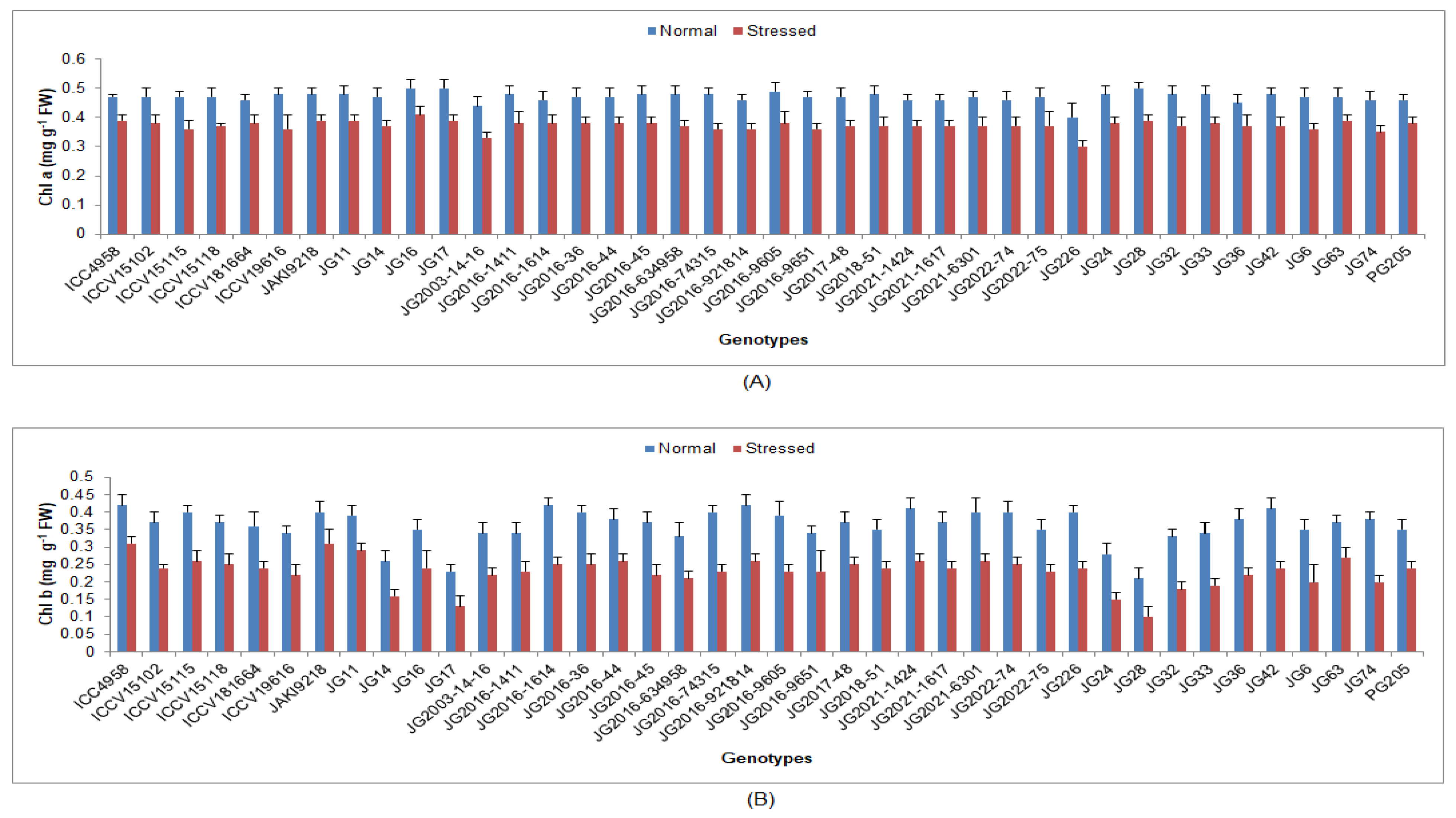
3.1. Effect of Terminal Drought Stress on Physiological Traits
3.2. Effect of Terminal Drought Stress on Biochemical Traits
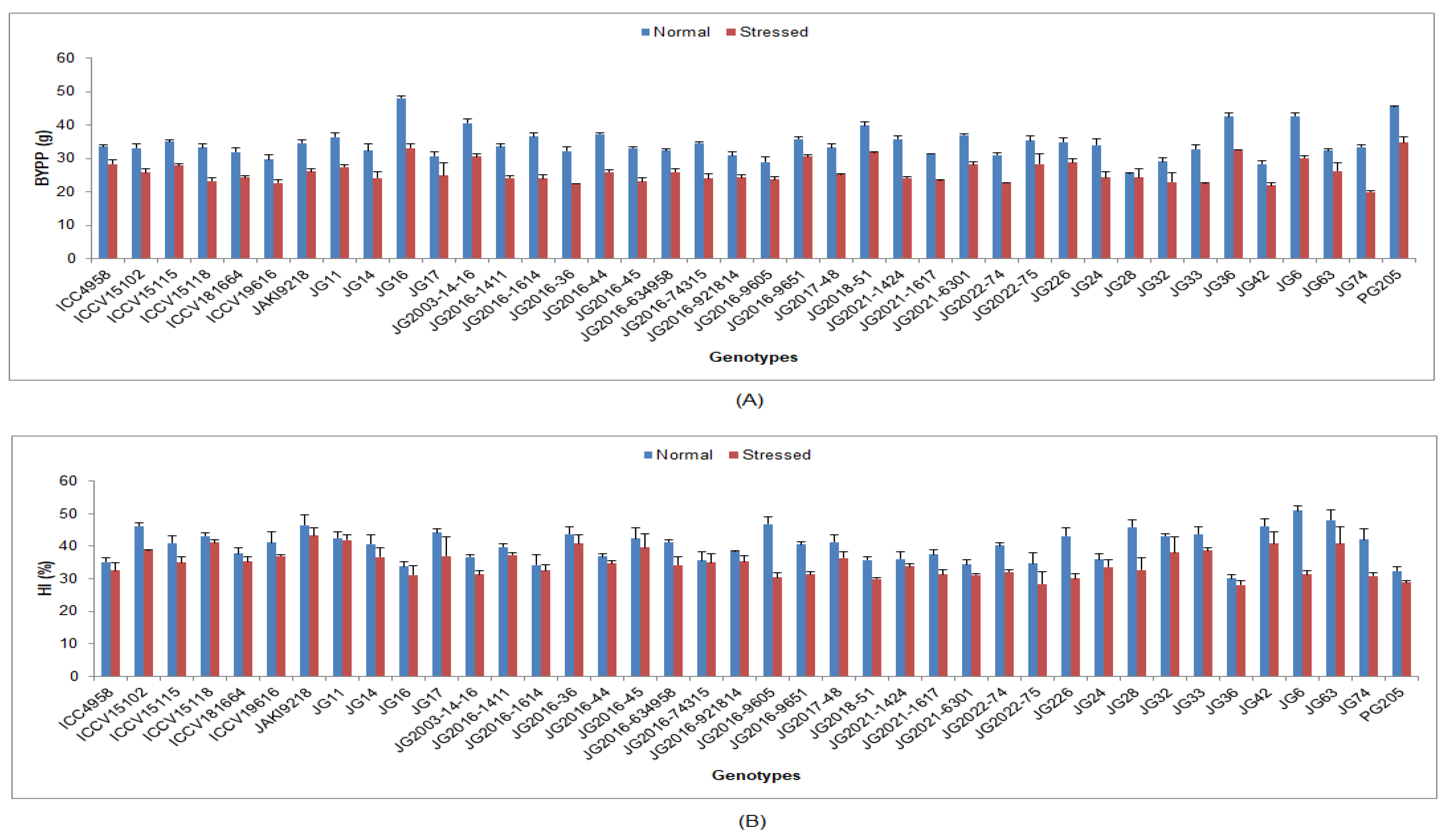
3.3. Effect of Terminal Drought Stress on Yield and Its Attributing Traits
3.4. Principle Component Biplot Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varol, I.S.; Kardes, Y.M.; Irik, H.A.; Kirnak, H.; Kaplan, M. Supplementary irrigations at different physiological growth stages of chickpea (Cicer arietinum L.) change grain nutritional composition. Food Chem. 2020, 303, 125402. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, J.; Malalgoda, M.; Storsley, J.; Malunga, L.; Netticadan, T.; Thandapilly, S.J. Health Benefits of Cereal Grain- and Pulse-Derived Proteins. Molecules 2022, 27, 3746. [Google Scholar] [CrossRef] [PubMed]
- Iquebal, M.A.; Soren, K.R.; Gangwar, P.; Shanmugavadivel, P.S.; Aravind, K.; Singla, D.; Jaiswal, S.; Jasrotia, R.S.; Chaturvedi, S.K.; Singh, N.P.; et al. Discovery of putative herbicide resistance genes and its regulatory network in chickpea using transcriptome sequencing. Front. Plant Sci. 2017, 8, 958. [Google Scholar] [CrossRef] [PubMed]
- Jameel, S.; Hameed, A.; Shah, T.M. Investigation of distinctive morpho-physio and biochemical alterations in desi chickpea at seedling stage under irrigation, heat, and combined stress. Front. Plant Sci. 2021, 12, 692745. [Google Scholar] [CrossRef]
- Thudi, M.; Roorkiwal, M.; Kudapa, H.; Chaturvedi, S.K.; Singh, N.P.; Varshney, R.K. An overview of chickpea research: From discovery to delivery. Pulse India 2017, 2, 22–25. [Google Scholar]
- Kaur, D.; Singh, G.; Sharma, P. Symbiotic parameters, productivity and profitability in Kabuli chickpea (Cicer arietinum L.) as influenced by application of phosphorus and biofertilizers. J. Soil Sci. Plant Nutr. 2020, 20, 2267–2282. [Google Scholar] [CrossRef]
- Faridy, J.C.M.; Stephanie, C.G.M.; Gabriela, M.M.O.; Cristian, J.M. Biological activities of chickpea in human health (Cicer arietinum L.). A review. Plant Foods Hum. Nutr. 2020, 75, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Yegrem, L. Nutritional composition, antinutritional factors, and utilization trends of Ethiopian chickpea (Cicer arietinum L.). Int. J. Food Sci. 2021, 2021, 5570753. [Google Scholar] [CrossRef]
- Yadav, R.K.; Tripathi, M.K.; Tiwari, S.; Tripathi, N.; Asati, R.; Patel, V.; Sikarwar, R.S.; Payasi, D.K. Breeding and Genomic Approaches towards Development of Fusarium Wilt Resistance in Chickpea. Life 2023, 13, 988. [Google Scholar] [CrossRef]
- FAOSTAT. Crop Statistics.2021. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 23 December 2022).
- Kushwah, A.; Bhatia, D.; Singh, G.; Singh, I.; Bindra, S.; Vij, S.; Singh, S. Phenotypic evaluation of genetic variability and selection of yield contributing traits in chickpea recombinant inbred line population under high temperature stress. Physiol. Mol. Biol. Plants. 2021, 27, 747–767. [Google Scholar] [CrossRef]
- Moreno, M.T.; Cubero, J.I. Variation in Cicer arietinum L. Euphytica 1978, 27, 465–485. [Google Scholar] [CrossRef]
- Solanki, R.S.; Babbar, A.; Tripathi, N. Genetic diversity analysis in kabuli chickpea (Cicer arietinum L.) genotypes based on quantitative traits and molecular markers. Bangladesh J. Bot. 2022, 53, 581–587. [Google Scholar] [CrossRef]
- Pande, S.; Siddique, K.H.M.; Kishore, G.K.; Bayaa, B.; Gaur, P.M.; Gowda, C.L.L.; Bretag, T.W.; Crouch, J.H. Ascochyta blight of chickpea (Cicer arietinum L.): A review of biology, pathogenicity, and disease management. Aust. J. Agric. Res. 2005, 56, 317–332. [Google Scholar] [CrossRef]
- Duke, J. Handbook of Legumes of World Economic Importance; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Shah, T.M.; Imran, M.; Atta, B.M.; Ashraf, M.Y.; Hameed, A.; Waqar, I.; Maqbool, M.A. Selection and screening of drought tolerant high yielding chickpea genotypes based on physio-biochemical indices and multi-environmental yield trials. BMC Plant Biol. 2020, 20, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sahu, K.K.; Chaudhary, J.L.; Puranik, H.V. Identification of weather parameters affecting the yield of chickpea under different growing environments. J. Pharmacogn. Phytochem. 2021, 10, 77–79. [Google Scholar]
- Shafiq, M.; Arif, M.; Akhtar, N.; Yousaf, M.; Ghaffar, A. Exogenous application of growth promoters can improve the chickpea productivity under terminal heat stress conditions by modulating the antioxidant enzyme system. Pak. J. Agri. Sci. 2021, 58, 35–42. [Google Scholar]
- Asati, R.; Tripathi, M.K.; Tiwari, S.; Yadav, R.K.; Tripathi, N. Molecular approaches in the development of drought tolerance in chickpea. Life 2022, 12, 1846. [Google Scholar] [CrossRef]
- Księżak, J.; Bojarszczuk, J. The effect of cropping method and botanical form on seed yielding and chemical composition of chickpeas (Cicer arietinum L.) grown under organic system. Agronomy 2020, 10, 801. [Google Scholar] [CrossRef]
- Shahzad, A.; Gul, H.; Ahsan, M.; Wang, D.; Fahad, S. Comparative genetic evaluation of maize inbred lines at seedling and maturity stages under drought stress. J. Plant Growth Regul. 2023, 42, 989–1005. [Google Scholar] [CrossRef]
- Habib, I.; Shahzad, K.; Rauf, M.; Ahmad, M.; Alsamadany, H.; Fahad, S.; Saeed, N.A. Dehydrin responsive HVA1 driven inducible gene expression enhanced salt and drought tolerance in wheat. Plant Physiol. Biochem. 2022, 180, 124–133. [Google Scholar] [CrossRef]
- Alam, H.; Zamin, M.; Adnan, M.; Shah, A.N.; Alharby, H.F.; Bamagoos, A.A.; Alabdallah, N.M.; Alzahrani, S.S.; Alharbi, B.M.; Saud, S.; et al. Exploring Suitability of Salsolaimbricata (Fetid Saltwort) for Salinity and Drought Conditions: A Step Toward Sustainable Landscaping Under Changing Climate. Front. Plant Sci. 2022, 13, 1–12. [Google Scholar] [CrossRef]
- Varshney, R.K.; Thudi, M.; Nayak, S.N.; Gaur, P.; Kashiwagi, J.; Krishnamurthy, L.; Jaganathan, D.; Koppolu, J.; Bohra, A.; Tripathi, S.; et al. Genetic dissection of drought tolerance in chickpea (Cicer arietinum L.). Theor. Appl. Genet. 2014, 127, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Tuberosa, R.; Salvi, S. Genomics-based approaches to improve drought tolerance of crops. Trends Plant Sci. 2006, 11, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, R.; Thudi, M.; Krishnamurthy, L.; Upadhyaya, H.D.; Kashiwagi, J.; Gowda, C.L.L.; Varshney, R.K. Association of mid-reproductive stage canopy temperature depression with the molecular markers and grain yields of chickpea (Cicer arietinum L.) germplasm under terminal drought. Field Crops Res. 2015, 174, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Shah, D.; Singh, M.P. Evaluation of chickpea (Cicer arietinum L.) genotypes for heat tolerance: A physiological assessment. Indian J. Plant Physiol. 2017, 22, 164–177. [Google Scholar] [CrossRef]
- Sachdeva, S.; Bharadwaj, C.; Patil, B.S.; Pal, M.; Roorkiwal, M.; Varshney, R.K. Agronomic Performance of Chickpea Affected by Drought Stress at Different Growth Stages. Agronomy 2022, 12, 995. [Google Scholar] [CrossRef]
- Dalvi, U.S.; Naik, R.M.; Lokhande, P.K. Antioxidant defense system in chickpea against drought stress at pre-and post-flowering stages. Indian J. Plant Physiol. 2018, 23, 16–23. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Amelioration of drought tolerance in wheat by the interaction of plant growth-promoting rhizobacteria. Plant Biol. 2016, 18, 992–1000. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Velikova, V.; Loreto, F. On the relationship between isoprene emission and thermotolerance in Phragmitesaustralis leaves exposed to high temperatures and during the recovery from a heat stress. Plant Cell Environ. 2005, 28, 318–327. [Google Scholar] [CrossRef]
- Naservafaei, S.; Sohrabi, Y.; Moradi, P.; Mac Sweeney, E.; Mastinu, A. Biological response of Lallemantiaiberica to brassinolide treatment under different watering conditions. Plants 2021, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Agarwal, P.K.; Jha, B. Improved salinity tolerance of Arachishypogaea (L.) by the interaction of halotolerant plant-growth-promoting rhizobacteria. J. Plant Growth Regul. 2012, 31, 195–206. [Google Scholar] [CrossRef]
- Sharma, A.; Gontia, I.; Agarwal, P.K.; Jha, B. Accumulation of heavy metals and its biochemical responses in Salicorniabrachiata, an extreme halophyte. Mar. Biol. Res. 2010, 6, 511–518. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B-and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Rani, A.; Devi, P.; Jha, U.C.; Sharma, K.D.; Siddique, K.H.; Nayyar, H. Developing climate-resilient chickpea involving physiological and molecular approaches with a focus on temperature and drought stresses. Front. Plant Sci. 2020, 10, 1759. [Google Scholar] [CrossRef]
- Awari, V.R.; Dalvi, U.S.; Lokhande, P.K.; Pawar, V.Y.; Mate, S.N.; Naik, R.M.; Mhase, L.B. Physiological and biochemical basis for moisture stress tolerance in chickpea under pot study. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1247–1259. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Ullah, A.; Lee, D.J.; Alghamdi, S.S.; Siddique, K.H. Desi chickpea genotypes tolerate drought stress better than kabuli types by modulating germination metabolism, trehalose accumulation, and carbon assimilation. Plant Physiol. Biochem. 2018, 126, 47–54. [Google Scholar] [CrossRef]
- Çevik, S.; Akpinar, G.; Yildizli, A.; Kasap, M.; Karaosmanoğlu, K.; Ünyayar, S. Comparative physiological and leaf proteome analysis between drought-tolerant chickpea Cicerreticulatum and drought-sensitive chickpea C. arietinum. J. Biosci. 2019, 44, 1–13. [Google Scholar] [CrossRef]
- Lokhande, P.K.; Naik, R.M.; Dalvi, U.S.; Mhase, L.B.; Harer, P.N. Antioxidative and root attributes response of chickpea parents and crosses under drought stress. Legume Res. 2019, 42, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Karimizadeh, R.; Mohammadi, M. Association of canopy temperature depression with yield of durum wheat genotypes under supplementary irrigated and rainfed conditions. Aust. J. Crop Sci. 2011, 5, 138–146. [Google Scholar]
- Shakeel, A.A.; Xiao-yu, X.; Long-chang, W.; Muhammad, F.S.; Chen, M.; Wang, L. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Armand, N.; Amiri, H.; Ismaili, A. Interaction of methanol spray and water-deficit stress on photosynthesis and biochemical characteristics of Phaseolus vulgaris L. cv. Sadry. Photochem. Photobiol. 2016, 92, 102–110. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.R.; Amiri, H.; Ismaili, A. Evaluation of photosynthesis, physiological, and biochemical responses of chickpea (Cicer arietinum L. cv. Pirouz) under water deficit stress and use of vermicompost fertilizer. J. Integr. Agric. 2018, 17, 2426–2437. [Google Scholar] [CrossRef] [Green Version]
- Seifikalhor, M.; Niknam, V.; Aliniaeifard, S.; Didaran, F.; Tsaniklidis, G.; Fanourakis, D.; Teymoorzadeh, M.; Mousavi, S.H.; Bosacchi, M.; Li, T. The regulatory role of γ-Aminobutyric acid in chickpea plants depends on drought tolerance and water scarcity level. Sci. Rep. 2022, 12, 1–17. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Role of ascorbic acid, glutathione and proline applied as singly or in sequence combination in improving chickpea plant through physiological change and antioxidant defense under different levels of irrigation intervals. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef] [Green Version]
- Jan, M.; Haq, T.U.; Sattar, H.; Butt, M.; Khaliq, A.; Arif, M.; Rauf, A. Evaluation and screening of promising drought tolerant chickpea (Cicer arietinum L.) genotypes based on physiological and biochemical attributes under drought conditions. Pak. J. Agric. Res. 2020, 33, 662–672. [Google Scholar] [CrossRef]
- Hussain, N.; Yasmeen, A.; Yousaf, M.M. Antioxidant status and their enhancements strategies for water stress tolerance in chickpea. Braz. J. Biol. 2021, 82, e237809. [Google Scholar] [CrossRef] [PubMed]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiwari, P.N.; Tiwari, S.; Sapre, S.; Babbar, A.; Tripathi, N.; Tiwari, S.; Tripathi, M.K. Screening and Selection of Drought-Tolerant High-Yielding Chickpea Genotypes Based on Physio-Biochemical Selection Indices and Yield Trials. Life 2023, 13, 1405. https://doi.org/10.3390/life13061405
Tiwari PN, Tiwari S, Sapre S, Babbar A, Tripathi N, Tiwari S, Tripathi MK. Screening and Selection of Drought-Tolerant High-Yielding Chickpea Genotypes Based on Physio-Biochemical Selection Indices and Yield Trials. Life. 2023; 13(6):1405. https://doi.org/10.3390/life13061405
Chicago/Turabian StyleTiwari, Prakash N., Sharad Tiwari, Swapnil Sapre, Anita Babbar, Niraj Tripathi, Sushma Tiwari, and Manoj Kumar Tripathi. 2023. "Screening and Selection of Drought-Tolerant High-Yielding Chickpea Genotypes Based on Physio-Biochemical Selection Indices and Yield Trials" Life 13, no. 6: 1405. https://doi.org/10.3390/life13061405