
Research Trends in C-Terminal Domain Nuclear Envelope Phosphatase 1


Abstract
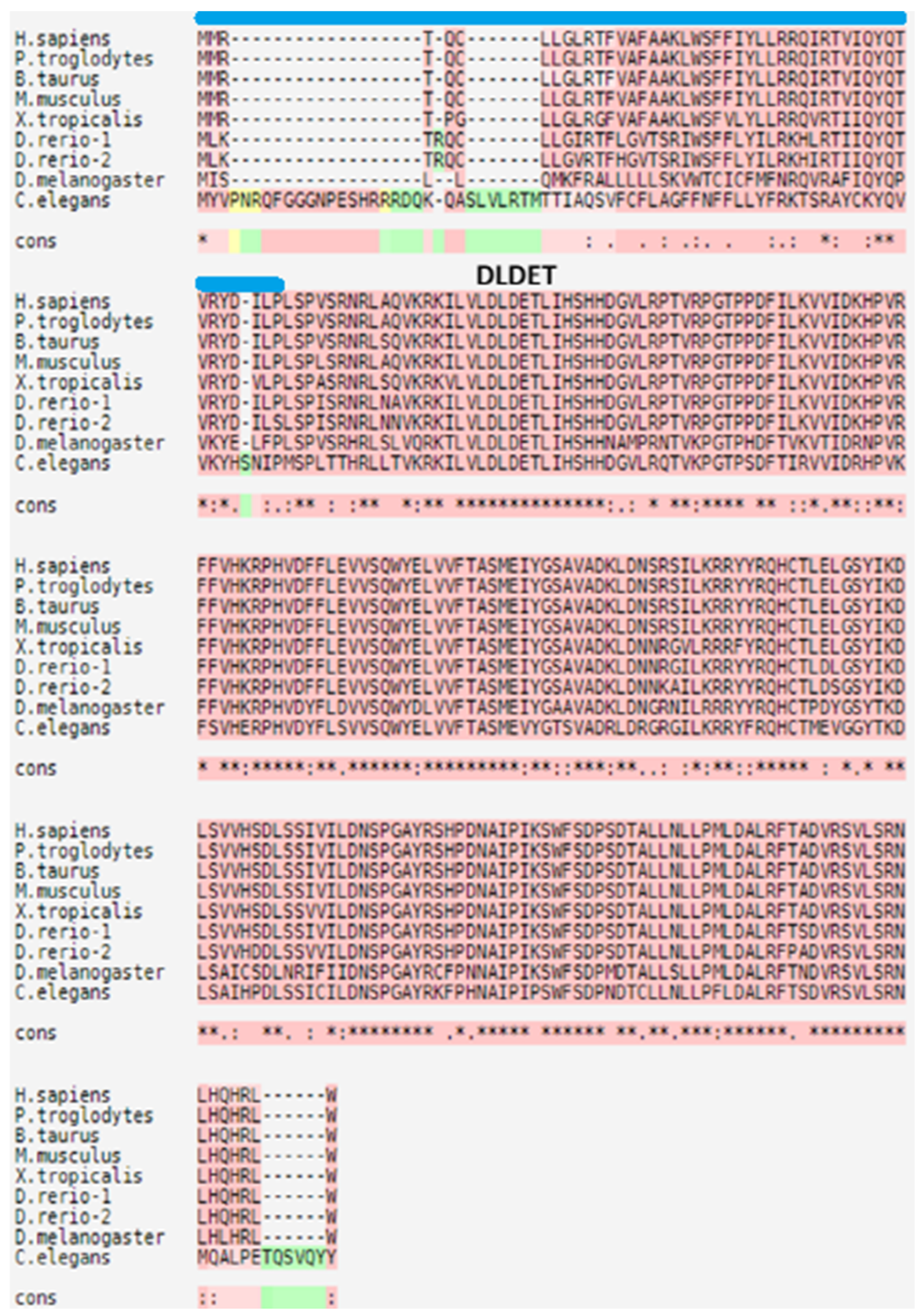
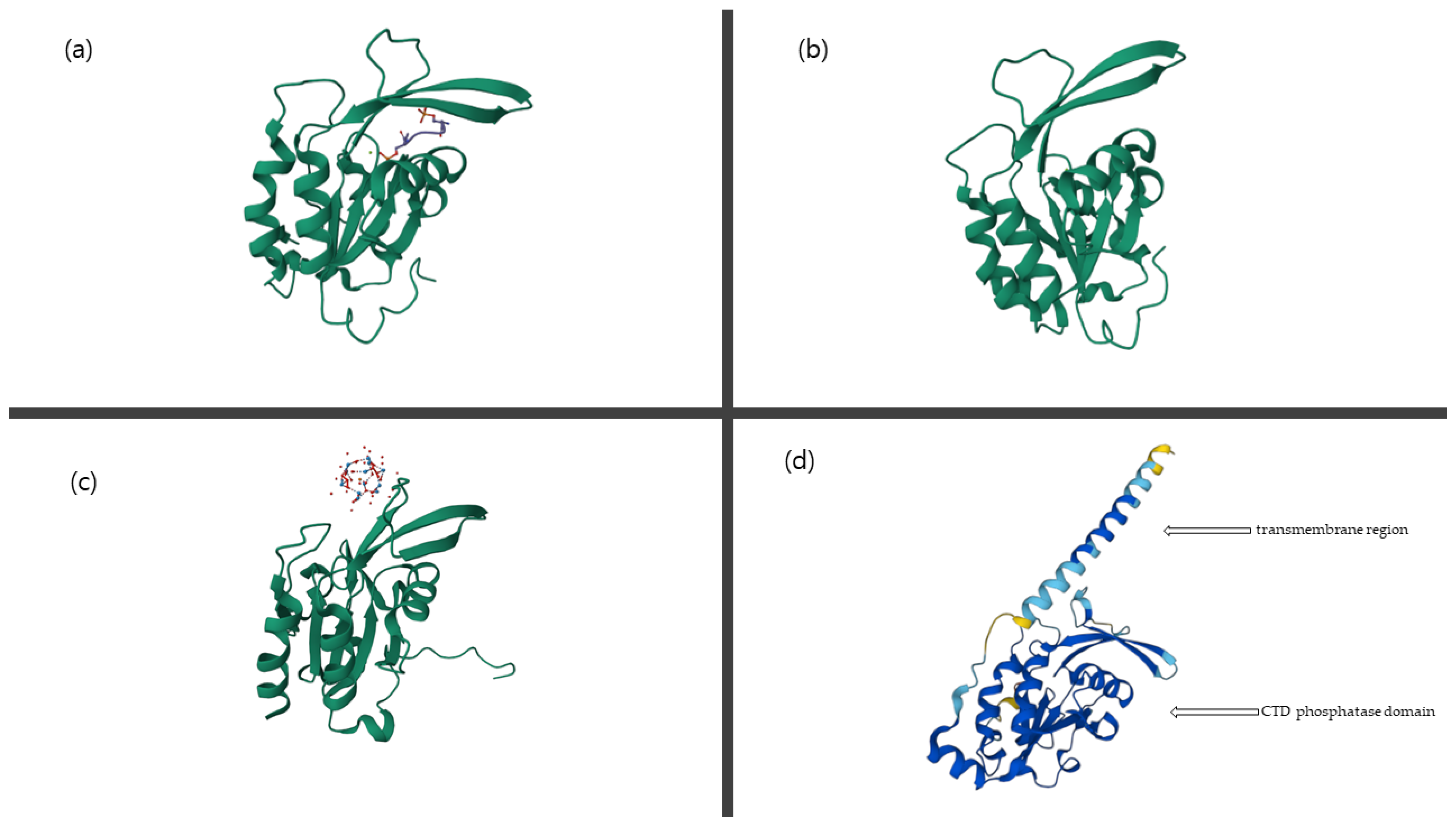
:1. Introduction

{kind=link}
{kind=link}
| Roles | Possible Substrates/Interacting Proteins | References |
|---|---|---|
| Nuclear membrane biogenesis | LIPIN1 PAH1 SUN2 | [22] [14] [23] |
| Nuclear pore complex insertion | Torsin | [24] |
| Nuclear positioning | Eps8L2 | [25] |
| Neural induction | BMPR | [11,13] |
| Kidney formation | BMPR SMAD 1/5 | [20,26] [27,28] |
| Primordial germ cell activation | unknown | [18] |
| Wing vein formation | pMAD DSRF | [19] [29,30] |
| Bone homeostasis Hemorrhage in the adult ovarian follicles | SMAD 2/3 unknown | [31] [32] |
| Tumor suppressor | MYC TOP2A CHEK1 | [8] [8] [8] |
| Control of birthweight Hepatosteatosis | unknown unknown | [33] [34] |
2. Biochemical Characterization of CTDNEP1
3. CTDNEP1 in Nuclear Membrane Biogenesis
4. CTDNEP1 in BMP-Mediated Biological Processes
5. CTDNEP1 in Other Biological Processes
6. Discussion and Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shao, L.; Ma, Y.; Fang, Q.; Huang, Z.; Wan, S.; Wang, J.; Yang, L. Role of protein phosphatase 2A in kidney disease (Review). Exp. Ther. Med. 2021, 22, 1236. [Google Scholar] [CrossRef] [PubMed]
- Harikrishna, R.R.; Kim, H.; Noh, K.; Kim, Y.J. The diverse roles of RNA polymerase II C-terminal domain phosphatase SCP1. BMB Rep. 2014, 47, 192–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, A.; Abbott, K.L.; Desjardins, A.; Di Lello, P.; Omichinski, J.G.; Legault, P. NMR structure of a complex formed by the carboxyl-terminal domain of human RAP74 and a phosphorylated peptide from the central domain of the FCP1 phosphatase. Biochemistry 2009, 48, 1964–1974. [Google Scholar] [CrossRef] [PubMed]
- Qiao, F.; Law, H.C.; Krieger, K.L.; Clement, E.J.; Xiao, Y.; Buckley, S.M.; Woods, N.T. Ctdp1 deficiency leads to early embryonic lethality in mice and defects in cell cycle progression in MEFs. Biol. Open 2021, 10, bio057232. [Google Scholar] [CrossRef]
- Yang, X.; Chen, Y.; Zhou, Y.; Wu, C.; Li, Q.; Wu, J.; Hu, W.W.; Zhao, W.Q.; Wei, W.; Wu, C.P.; et al. GPC5 suppresses lung cancer progression and metastasis via intracellular CTDSP1/AhR/ARNT signaling axis and extracellular exosome secretion. Oncogene 2021, 40, 4307–4323. [Google Scholar] [CrossRef]
- Kloet, D.E.; Polderman, P.E.; Eijkelenboom, A.; Smits, L.M.; van Triest, M.H.; van den Berg, M.C.; Groot Koerkamp, M.J.; van Leenen, D.; Lijnzaad, P.; Holstege, F.C.; et al. FOXO target gene CTDSP2 regulates cell cycle progression through Ras and p21(Cip1/Waf1). Biochem. J. 2015, 469, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; He, X.; Li, F.; Pan, H.; Huang, X.; Wen, X.; Zhang, H.; Li, B.; Ge, S.; Xu, X.; et al. The miR-181 family promotes cell cycle by targeting CTDSPL, a phosphatase-like tumor suppressor in uveal melanoma. J. Exp. Clin. Cancer Res. 2018, 37, 15. [Google Scholar] [CrossRef]
- Luo, Z.; Xin, D.; Liao, Y.; Berry, K.; Ogurek, S.; Zhang, F.; Zhang, L.; Zhao, C.; Rao, R.; Dong, X.; et al. Loss of phosphatase CTDNEP1 potentiates aggressive medulloblastoma by triggering MYC amplification and genomic instability. Nat. Commun. 2023, 14, 762. [Google Scholar] [CrossRef]
- Xiao, Y.; Chen, Y.; Peng, A.; Dong, J. The phosphatase CTDSPL2 is phosphorylated in mitosis and a target for restraining tumor growth and motility in pancreatic cancer. Cancer Lett. 2022, 526, 53–65. [Google Scholar] [CrossRef]
- Sun, S.; Liu, S.; Zhang, Z.; Zeng, W.; Sun, C.; Tao, T.; Lin, X.; Feng, X.H. Phosphatase UBLCP1 controls proteasome assembly. Open Biol. 2017, 7, 170042. [Google Scholar] [CrossRef] [Green Version]
- Satow, R.; Chan, T.C.; Asashima, M. Molecular cloning and characterization of dullard: A novel gene required for neural development. Biochem. Biophys. Res. Commun. 2002, 295, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Gentry, M.S.; Harris, T.E.; Wiley, S.E.; Lawrence, J.C., Jr.; Dixon, J.E. A conserved phosphatase cascade that regulates nuclear membrane biogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 6596–6601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satow, R.; Kurisaki, A.; Chan, T.C.; Hamazaki, T.S.; Asashima, M. Dullard promotes degradation and dephosphorylation of BMP receptors and is required for neural induction. Dev. Cell 2006, 11, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Khondker, S.; Han, G.S.; Carman, G.M. Phosphorylation-mediated regulation of the Nem1-Spo7/Pah1 phosphatase cascade in yeast lipid synthesis. Adv. Biol. Regul. 2022, 84, 100889. [Google Scholar] [CrossRef] [PubMed]
- Kwiatek, J.M.; Gutierrez, B.; Izgu, E.C.; Han, G.S.; Carman, G.M. Phosphatidic Acid Mediates the Nem1-Spo7/Pah1 Phosphatase Cascade in Yeast Lipid Synthesis. J. Lipid Res. 2022, 63, 100282. [Google Scholar] [CrossRef] [PubMed]
- Mirheydari, M.; Dey, P.; Stukey, G.J.; Park, Y.; Han, G.S.; Carman, G.M. The Spo7 sequence LLI is required for Nem1-Spo7/Pah1 phosphatase cascade function in yeast lipid metabolism. J. Biol. Chem. 2020, 295, 11473–11485. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.; Wang, J.; Li, S.; Sun, C.; Jiang, W.; Xin, Q.; Wang, J.; Qi, T.; Li, K.; Zhang, Z.; et al. A review of BMP and Wnt signaling pathway in the pathogenesis of pulmonary arterial hypertension. Clin. Exp. Hypertens. 2022, 44, 175–180. [Google Scholar] [CrossRef]
- Tanaka, S.S.; Nakane, A.; Yamaguchi, Y.L.; Terabayashi, T.; Abe, T.; Nakao, K.; Asashima, M.; Steiner, K.A.; Tam, P.P.; Nishinakamura, R. Dullard/Ctdnep1 modulates WNT signalling activity for the formation of primordial germ cells in the mouse embryo. PLoS ONE 2013, 8, e57428. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Matsuoka, S.; Enoki, A.; Yamamoto, T.; Furukawa, K.; Yamasaki, Y.; Nishida, Y.; Sugiyama, S. Negative modulation of bone morphogenetic protein signaling by Dullard during wing vein formation in Drosophila. Dev. Growth Differ. 2011, 53, 822–841. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Sharmin, S.; Taguchi, A.; Ohmori, T.; Fujimura, S.; Abe, T.; Kiyonari, H.; Komatsu, Y.; Mishina, Y.; Asashima, M.; et al. The phosphatase Dullard negatively regulates BMP signalling and is essential for nephron maintenance after birth. Nat. Commun. 2013, 4, 1398. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Muraoka, M.; Okada, H.; Toyoda, A.; Ajima, R.; Saga, Y. The RNA helicase DDX6 controls early mouse embryogenesis by repressing aberrant inhibition of BMP signaling through miRNA-mediated gene silencing. PLoS Genet. 2022, 18, e1009967. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Garland, M.; Dunaway-Mariano, D.; Allen, K.N. Homo sapiens dullard protein phosphatase shows a preference for the insulin-dependent phosphorylation site of lipin1. Biochemistry 2011, 50, 3045–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krshnan, L.; Siu, W.S.; Van de Weijer, M.; Hayward, D.; Guerrero, E.N.; Gruneberg, U.; Carvalho, P. Regulated degradation of the inner nuclear membrane protein SUN2 maintains nuclear envelope architecture and function. eLife 2022, 11, e81573. [Google Scholar] [CrossRef] [PubMed]
- Jacquemyn, J.; Foroozandeh, J.; Vints, K.; Swerts, J.; Verstreken, P.; Gounko, N.V.; Gallego, S.F.; Goodchild, R. Torsin and NEP1R1-CTDNEP1 phosphatase affect interphase nuclear pore complex insertion by lipid-dependent and lipid-independent mechanisms. EMBO J. 2021, 40, e106914. [Google Scholar] [CrossRef] [PubMed]
- Calero-Cuenca, F.J.; Osorio, D.S.; Carvalho-Marques, S.; Sridhara, S.C.; Oliveira, L.M.; Jiao, Y.; Diaz, J.; Janota, C.S.; Cadot, B.; Gomes, E.R. Ctdnep1 and Eps8L2 regulate dorsal actin cables for nuclear positioning during cell migration. Curr. Biol. 2021, 31, 1521–1530. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Oshima, K.; Fogo, A.; Hogan, B.L.; Ichikawa, I. Bone morphogenetic protein 4 regulates the budding site and elongation of the mouse ureter. J. Clin. Investig. 2000, 105, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, P.; Haag, J.; Campean, V.; Goldschmeding, R.; Atalla, A.; Amann, K.; Aigner, T. Bone morphogenetic protein-7 expression and activity in the human adult normal kidney is predominantly localized to the distal nephron. Kidney Int. 2006, 70, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Oxburgh, L.; Brown, A.C.; Muthukrishnan, S.D.; Fetting, J.L. Bone morphogenetic protein signaling in nephron progenitor cells. Pediatr. Nephrol. 2014, 29, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Guichard, A.; Biehs, B.; Sturtevant, M.A.; Wickline, L.; Chacko, J.; Howard, K.; Bier, E. rhomboid and Star interact synergistically to promote EGFR/MAPK signaling during Drosophila wing vein development. Development 1999, 126, 2663–2676. [Google Scholar] [CrossRef]
- Sieber, C.; Kopf, J.; Hiepen, C.; Knaus, P. Recent advances in BMP receptor signaling. Cytokine Growth Factor Rev. 2009, 20, 343–355. [Google Scholar] [CrossRef]
- Hayata, T.; Ezura, Y.; Asashima, M.; Nishinakamura, R.; Noda, M. Dullard/Ctdnep1 regulates endochondral ossification via suppression of TGF-beta signaling. J. Bone Min. Res. 2015, 30, 947. [Google Scholar] [CrossRef] [PubMed]
- Hayata, T.; Chiga, M.; Ezura, Y.; Asashima, M.; Katabuchi, H.; Nishinakamura, R.; Noda, M. Dullard deficiency causes hemorrhage in the adult ovarian follicles. Genes Cells 2018, 23, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekola-Ayele, F.; Zeng, X.; Chatterjee, S.; Ouidir, M.; Lesseur, C.; Hao, K.; Chen, J.; Tesfaye, M.; Marsit, C.J.; Workalemahu, T.; et al. Placental multi-omics integration identifies candidate functional genes for birthweight. Nat. Commun. 2022, 13, 2384. [Google Scholar] [CrossRef]
- Naderi, M.; Pazouki, A.; Arefian, E.; Hashemi, S.M.; Jamshidi-Adegani, F.; Gholamalamdari, O.; Soudi, S.; Azadmanesh, K.; Mirab Samiee, S.; Merat, S.; et al. Two Triacylglycerol Pathway Genes, CTDNEP1 and LPIN1, are Down-Regulated by hsa-miR-122-5p in Hepatocytes. Arch. Iran. Med. 2017, 20, 165–171. [Google Scholar]
- Han, S.; Bahmanyar, S.; Zhang, P.; Grishin, N.; Oegema, K.; Crooke, R.; Graham, M.; Reue, K.; Dixon, J.E.; Goodman, J.M. Nuclear envelope phosphatase 1-regulatory subunit 1 (formerly TMEM188) is the metazoan Spo7p ortholog and functions in the lipin activation pathway. J. Biol. Chem. 2012, 287, 3123–3137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donkor, J.; Sariahmetoglu, M.; Dewald, J.; Brindley, D.N.; Reue, K. Three mammalian lipins act as phosphatidate phosphatases with distinct tissue expression patterns. J. Biol. Chem. 2007, 282, 3450–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, G.S.; Siniossoglou, S.; Carman, G.M. The cellular functions of the yeast lipin homolog PAH1p are dependent on its phosphatidate phosphatase activity. J. Biol. Chem. 2007, 282, 37026–37035. [Google Scholar] [CrossRef] [Green Version]
- Huffman, T.A.; Mothe-Satney, I.; Lawrence, J.C., Jr. Insulin-stimulated phosphorylation of lipin mediated by the mammalian target of rapamycin. Proc. Natl. Acad. Sci. USA 2002, 99, 1047–1052. [Google Scholar] [CrossRef] [Green Version]
- Castro, V.; Calvo, G.; Avila-Perez, G.; Dreux, M.; Gastaminza, P. Differential Roles of Lipin1 and Lipin2 in the Hepatitis C Virus Replication Cycle. Cells 2019, 8, 1456. [Google Scholar] [CrossRef] [Green Version]
- Dou, J.Y.; Jiang, Y.C.; Hu, Z.H.; Yao, K.C.; Yuan, M.H.; Bao, X.X.; Zhou, M.J.; Liu, Y.; Li, Z.X.; Lian, L.H.; et al. Betulin Targets Lipin1/2-Meidated P2X7 Receptor as a Therapeutic Approach to Attenuate Lipid Accumulation and Metaflammation. Biomol. Ther. 2022, 30, 246–256. [Google Scholar] [CrossRef]
- Hamel, Y.; Mauvais, F.X.; Madrange, M.; Renard, P.; Lebreton, C.; Nemazanyy, I.; Pelle, O.; Goudin, N.; Tang, X.; Rodero, M.P.; et al. Compromised mitochondrial quality control triggers lipin1-related rhabdomyolysis. Cell Rep. Med. 2021, 2, 100370. [Google Scholar] [CrossRef] [PubMed]
- Jama, A.; Alshudukhi, A.A.; Burke, S.; Dong, L.; Kamau, J.K.; Voss, A.A.; Ren, H. Lipin1 plays complementary roles in myofibre stability and regeneration in dystrophic muscles. J. Physiol. 2023, 601, 961–978. [Google Scholar] [CrossRef] [PubMed]
- Kakehi, S.; Tamura, Y.; Ikeda, S.I.; Kaga, N.; Taka, H.; Ueno, N.; Shiuchi, T.; Kubota, A.; Sakuraba, K.; Kawamori, R.; et al. Short-term physical inactivity induces diacylglycerol accumulation and insulin resistance in muscle via lipin1 activation. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E766–E781. [Google Scholar] [CrossRef]
- Kihira, Y.; Fujimura, Y.; Tomita, S.; Tamaki, T.; Sato, E. Hypoxia-inducible factor-1alpha regulates Lipin1 differently in pre-adipocytes and mature adipocytes. Mol. Med. Rep. 2020, 22, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Kostic, M.; Koricanac, G.; Tepavcevic, S.; Stanisic, J.; Romic, S.; Culafic, T.; Ivkovic, T.; Stojiljkovic, M. Low-Intensity Exercise Affects Cardiac Fatty Acid Oxidation by Increasing the Nuclear Content of PPARalpha, FOXO1, and Lipin1 in Fructose-Fed Rats. Metab. Syndr. Relat. Disord. 2023, 21, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Salsman, J.; Foster, J.; Dellaire, G.; Ridgway, N.D. Lipid-associated PML structures assemble nuclear lipid droplets containing CCTalpha and Lipin1. Life Sci. Alliance 2020, 3, e202000751. [Google Scholar] [CrossRef]
- Ramani Sattiraju, S.; Jama, A.; Alshudukhi, A.A.; Edward Townsend, N.; Reynold Miranda, D.; Reese, R.R.; Voss, A.A.; Ren, H. Loss of membrane integrity drives myofiber death in lipin1-deficient skeletal muscle. Physiol. Rep. 2020, 8, e14620. [Google Scholar] [CrossRef]
- Shang, P.; Zheng, F.; Han, F.; Song, Y.; Pan, Z.; Yu, S.; Zhuang, X.; Chen, S. Lipin1 mediates cognitive impairment in fld mice via PKD-ERK pathway. Biochem. Biophys. Res. Commun. 2020, 525, 286–291. [Google Scholar] [CrossRef]
- Wang, M.; Xie, M.; Yu, S.; Shang, P.; Zhang, C.; Han, X.; Fan, C.; Chen, L.; Zhuang, X.; Chen, S. Lipin1 Alleviates Autophagy Disorder in Sciatic Nerve and Improves Diabetic Peripheral Neuropathy. Mol. Neurobiol. 2021, 58, 6049–6061. [Google Scholar] [CrossRef]
- Merta, H.; Carrasquillo Rodriguez, J.W.; Anjur-Dietrich, M.I.; Vitale, T.; Granade, M.E.; Harris, T.E.; Needleman, D.J.; Bahmanyar, S. Cell cycle regulation of ER membrane biogenesis protects against chromosome missegregation. Dev. Cell 2021, 56, 3364–3379. [Google Scholar] [CrossRef]
- Barger, S.R.; Penfield, L.; Bahmanyar, S. Coupling lipid synthesis with nuclear envelope remodeling. Trends Biochem. Sci. 2021, 47, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Penfield, L.; Shankar, R.; Szentgyorgyi, E.; Laffitte, A.; Mauro, M.S.; Audhya, A.; Muller-Reichert, T.; Bahmanyar, S. Regulated lipid synthesis and LEM2/CHMP7 jointly control nuclear envelope closure. J. Cell Biol. 2019, 219, e201908179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waite, K.A.; Eng, C. From developmental disorder to heritable cancer: It’s all in the BMP/TGF-beta family. Nat. Rev. Genet. 2003, 4, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Nishinakamura, R.; Sakaguchi, M. BMP signaling and its modifiers in kidney development. Pediatr. Nephrol. 2014, 29, 681–686. [Google Scholar] [CrossRef]
- Matsuda, S.; Yoshiyama, N.; Kunnapuu-Vulli, J.; Hatakeyama, M.; Shimmi, O. Dpp/BMP transport mechanism is required for wing venation in the sawfly Athalia rosae. Insect Biochem. Mol. Biol. 2013, 43, 466–473. [Google Scholar] [CrossRef]
- Jones, D.T.; Jager, N.; Kool, M.; Zichner, T.; Hutter, B.; Sultan, M.; Cho, Y.J.; Pugh, T.J.; Hovestadt, V.; Stutz, A.M.; et al. Dissecting the genomic complexity underlying medulloblastoma. Nature 2012, 488, 100–105. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Li, Q.; Wu, X.; Li, W.; Li, Q.; Zhang, J.; Li, M.; Zhang, D.; Zhao, H.; Zou, X.; et al. Deubiquitinase USP29 promotes gastric cancer cell migration by cooperating with phosphatase SCP1 to stabilize Snail protein. Oncogene 2020, 39, 6802–6815. [Google Scholar] [CrossRef]
- Sun, A.G.; Wang, M.G.; Li, B.; Meng, F.G. Down-regulation of miR-124 target protein SCP-1 inhibits neuroglioma cell migration. Eur. Rev. Med. Pharm. Sci. 2017, 21, 723–729. [Google Scholar]
- Wang, W.; Liao, P.; Shen, M.; Chen, T.; Chen, Y.; Li, Y.; Lin, X.; Ge, X.; Wang, P. SCP1 regulates c-Myc stability and functions through dephosphorylating c-Myc Ser62. Oncogene 2016, 35, 491–500. [Google Scholar] [CrossRef]
- Zou, J.; Pei, X.; Xing, D.; Wu, X.; Chen, S. LINC00261 elevation inhibits angiogenesis and cell cycle progression of pancreatic cancer cells by upregulating SCP2 via targeting FOXP3. J. Cell Mol. Med. 2021, 25, 9826–9836. [Google Scholar] [CrossRef]
- Ding, X.; Fan, K.; Hu, J.; Zang, Z.; Zhang, S.; Zhang, Y.; Lin, Z.; Pei, X.; Zheng, X.; Zhu, F.; et al. SCP2-mediated cholesterol membrane trafficking promotes the growth of pituitary adenomas via Hedgehog signaling activation. J. Exp. Clin. Cancer Res. 2019, 38, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senchenko, V.N.; Anedchenko, E.A.; Kondratieva, T.T.; Krasnov, G.S.; Dmitriev, A.A.; Zabarovska, V.I.; Pavlova, T.V.; Kashuba, V.I.; Lerman, M.I.; Zabarovsky, E.R. Simultaneous down-regulation of tumor suppressor genes RBSP3/CTDSPL, NPRL2/G21 and RASSF1A in primary non-small cell lung cancer. BMC Cancer 2010, 10, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashuba, V.I.; Pavlova, T.V.; Grigorieva, E.V.; Kutsenko, A.; Yenamandra, S.P.; Li, J.; Wang, F.; Protopopov, A.I.; Zabarovska, V.I.; Senchenko, V.; et al. High mutability of the tumor suppressor genes RASSF1 and RBSP3 (CTDSPL) in cancer. PLoS ONE 2009, 4, e5231. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Xiao, M.; Sun, B.; Zhang, Z.; Shen, T.; Duan, X.; Yu, P.B.; Feng, X.H.; Lin, X. C-terminal domain (CTD) small phosphatase-like 2 modulates the canonical bone morphogenetic protein (BMP) signaling and mesenchymal differentiation via Smad dephosphorylation. J. Biol. Chem. 2014, 289, 26441–26450. [Google Scholar] [CrossRef] [Green Version]
- Chasov, V.; Mirgayazova, R.; Zmievskaya, E.; Khadiullina, R.; Valiullina, A.; Stephenson Clarke, J.; Rizvanov, A.; Baud, M.G.J.; Bulatov, E. Key Players in the Mutant p53 Team: Small Molecules, Gene Editing, Immunotherapy. Front. Oncol. 2020, 10, 1460. [Google Scholar] [CrossRef]
- Chasov, V.; Zaripov, M.; Mirgayazova, R.; Khadiullina, R.; Zmievskaya, E.; Ganeeva, I.; Valiullina, A.; Rizvanov, A.; Bulatov, E. Promising New Tools for Targeting p53 Mutant Cancers: Humoral and Cell-Based Immunotherapies. Front. Immunol. 2021, 12, 707734. [Google Scholar] [CrossRef] [PubMed]


| Research Fields | Research Topics |
|---|---|
| Biochemical characterization | Determination of the three-dimensional structure of CTDNEP1. Identification of interacting mechanism between CTDNEP1 and NEP1-R1. Determination of complex structure of CTDNEP1 and its substrates (e.g., LIPIN1). Kinetic investigation of CTDNEP1 with its possible substrates or interacting proteins. Identification of a regulatory material of CTDNEP1. |
| Nuclear envelope formation | Determination of upstream signaling of CTDNEP1. Identification of downstream signaling after PA dephosphorylation. Identification of a novel mechanism for nuclear membrane biogenesis. |
| BMP signaling | Identification of novel substrates of CTDNEP1 as a BMP signaling regulator. Determination of whether CTDNEP1 directly dephosphorylates BMPR, SMAD, or pMAD. If not, the identification of the indirect pathway. Identification of a novel BMP-related signaling. Characterization of the different aspects of CTDNEP1 from other protein phosphatases’ action on BMP signaling. |
| Tumor suppressor | Identification of novel substrates of CTDNEP1 as a tumor suppressor. Characterization of the different aspects of CTDNEP1 from other protein phosphatases’ action on tumor signaling. Identification of a novel related tumor. Investigation on targeting mutant CTDNEP1 tumors. |
| Another signaling | Determination of the mechanism of CTDNEP1 in birthweight control and hepatosteatosis. Investigation of miR-122 effect on CTDNEP1-related signaling. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rallabandi, H.R.; Choi, H.; Cha, H.; Kim, Y.J. Research Trends in C-Terminal Domain Nuclear Envelope Phosphatase 1. Life 2023, 13, 1338. https://doi.org/10.3390/life13061338
Rallabandi HR, Choi H, Cha H, Kim YJ. Research Trends in C-Terminal Domain Nuclear Envelope Phosphatase 1. Life. 2023; 13(6):1338. https://doi.org/10.3390/life13061338
Chicago/Turabian StyleRallabandi, Harikrishna Reddy, Haewon Choi, Hyunseung Cha, and Young Jun Kim. 2023. "Research Trends in C-Terminal Domain Nuclear Envelope Phosphatase 1" Life 13, no. 6: 1338. https://doi.org/10.3390/life13061338