
Impact of Chorionic Somatomammotropin In Vivo RNA Interference Phenotype on Uteroplacental Expression of the IGF Axis
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
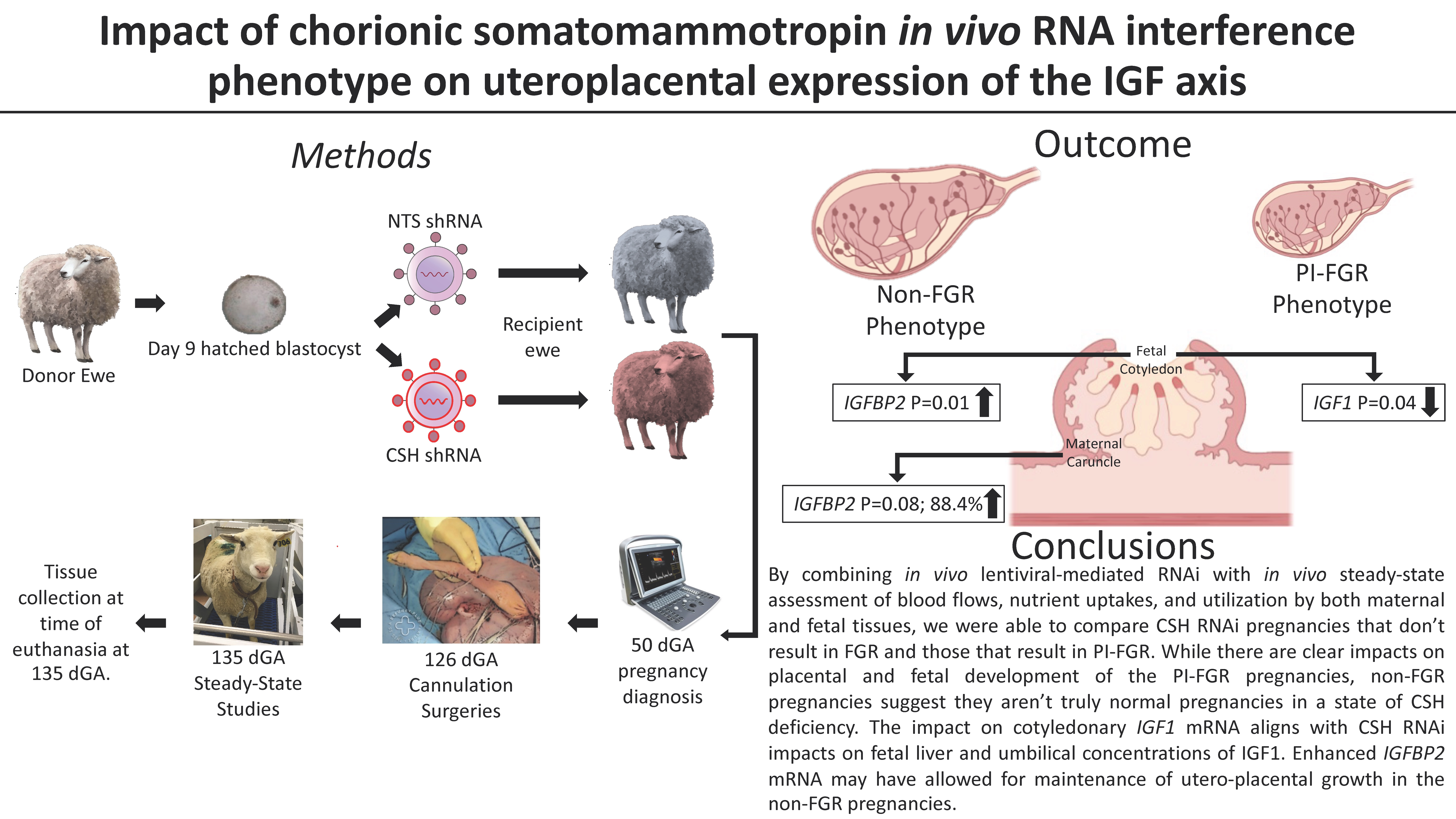
2.1. Experimental Groups
2.2. Lentiviral Generation
2.3. Generation of CSH RNAi Pregnancies
2.4. Surgical Instrumentation of Fetus and Ewe
2.5. Tissue Collection
2.6. RNA Isolation
2.7. Complimentary DNA Synthesis and Quantitative Real-Time PCR (qPCR)
2.8. Statistical Analysis
3. Results
3.1. Fetal and Placental Measurements
3.2. 135 dGA Plasma Hormone Concentrations
3.3. 135 dGA Placental IGF’s, IGFBP’s, and IGFR’s mRNA Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, R.L.; Owens, K.; Sumser, E.K.; Fry, M.V.; Stephens, K.K.; Chuecos, M.; Carrillo, M.; Schlabritz-Loutsevitch, N.; Jones, H.N. Nanoparticle mediated increased insulin-like growth factor 1 expression enhances human placenta syncytium function. Placenta 2020, 93, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-P.; Baker, J.; Perkins, A.S.; Robertson, E.J.; Efstratiadis, A. Mice carrying null mutations of the genes encoding insulin-like growth factor I (Igf-1) and type 1 IGF receptor (Igf1r). Cell 1993, 75, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.; Liu, J.P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef] [PubMed]
- de Vrijer, B.; Davidsen, M.L.; Wilkening, R.B.; Anthony, R.V.; Regnault, T.R.H. Altered Placental and Fetal Expression of IGFs and IGF-Binding Proteins Associated with Intrauterine Growth Restriction in Fetal Sheep During Early and Mid-Pregnancy. Pediatr. Res. 2006, 60, 507–512. [Google Scholar] [CrossRef]
- Denley, A.; Cosgrove, L.J.; Booker, G.W.; Wallace, J.C.; Forbes, B.E. Molecular interactions of the IGF system. Cytokine Growth Factor Rev. 2005, 16, 421–439. [Google Scholar] [CrossRef]
- Sibley, C.P.; Coan, P.M.; Ferguson-Smith, A.C.; Dean, W.; Hughes, J.; Smith, P.; Reik, W.; Burton, G.J.; Fowden, A.L.; Constância, M. Placental-specific insulin-like growth factor 2 (Igf2) regulates the diffusional exchange characteristics of the mouse placenta. Proc. Natl. Acad. Sci. USA 2004, 101, 8204–8208. [Google Scholar] [CrossRef]
- Coan, P.M.; Fowden, A.L.; Constancia, M.; Ferguson-Smith, A.C.; Burton, G.J.; Sibley, C.P. Disproportional effects of Igf2 knockout on placental morphology and diffusional exchange characteristics in the mouse. J. Physiol. 2008, 586, 5023–5032. [Google Scholar] [CrossRef]
- Ong, K.; Kratzsch, J.; Kiess, W.; ALSPAC Study Team; Costello, M.; Scott, C.; Dunger, D. Size at Birth and Cord Blood Levels of Insulin, Insulin-Like Growth Factor I (IGF-I), IGF-II, IGF-Binding Protein-1 (IGFBP-1), IGFBP-3, and the Soluble IGF-II/Mannose-6-Phosphate Receptor in Term Human Infants. J. Clin. Endocrinol. Metab. 2000, 85, 4266–4269. [Google Scholar] [CrossRef]
- Larsen, T.; Main, K.; Andersson, A.M.; Juul, A.; Greisen, G.; Skakkebæk, N.E. Growth hormone, insulin-like growth factor I and its binding proteins 1 and 3 in last trimester intrauterine growth retardation with increased pulsatility index in the umbilical artery. Clin. Endocrinol. 1996, 45, 315–319. [Google Scholar] [CrossRef]
- Ludwig, T.; Eggenschwiler, J.; Fisher, P.; D’Ercole, A.J.; Davenport, M.L.; Efstratiadis, A. Mouse mutants lacking the type 2 IGF receptor (IGF2R) are rescued from perinatal lethality in Igf2 and Igf1r null backgrounds. Dev. Biol. 1996, 177, 517–535. [Google Scholar] [CrossRef]
- Lau, M.M.; Stewart, C.E.; Liu, Z.; Bhatt, H.; Rotwein, P.; Stewart, C.L. Loss of the imprinted IGF2/cation-independent mannose 6-phosphate receptor results in fetal overgrowth and perinatal lethality. Genes Dev. 1994, 8, 2953–2963. [Google Scholar] [CrossRef] [PubMed]
- Handwerger, S. Clinical counterpoint: The Physiology of Placental Lactogen in Human Pregnancy. Endocr. Rev. 1991, 12, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.M.; Goetzmann, L.N.; Cantlon, J.D.; Jeckel, K.M.; Winger, Q.A.; Anthony, R.V. Development of ovine chorionic somatomammotropin hormone-deficient pregnancies. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R837–R846. [Google Scholar] [CrossRef] [PubMed]
- Jeckel, K.M.; Boyarko, A.C.; Bouma, G.J.; Winger, Q.A.; Anthony, R.V. Chorionic somatomammotropin impacts early fetal growth and placental gene expression. J. Endocrinol. 2018, 237, 301–310. [Google Scholar] [CrossRef]
- Tanner, A.R.; Lynch, C.S.; Kennedy, V.C.; Ali, A.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. CSH RNA Interference Reduces Global Nutrient Uptake and Umbilical Blood Flow Resulting in Intrauterine Growth Restriction. Int. J. Mol. Sci. 2021, 22, 8150. [Google Scholar] [CrossRef]
- Tanner, A.R.; Kennedy, V.C.; Lynch, C.S.; Hord, T.K.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. In Vivo investigation of ruminant placenta function and physiology—A review. J. Anim. Sci. 2022, 100, skac045. [Google Scholar] [CrossRef]
- Ali, A.; Swanepoel, C.M.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. Chorionic somatomammotropin RNA interference alters fetal liver glucose utilization. J. Endocrinol. 2020, 247, 169–180. [Google Scholar] [CrossRef]
- Tanner, A.R.; Lynch, C.S.; Ali, A.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. Impact of chorionic somatomammotropin RNA interference on uterine blood flow and placental glucose uptake in the absence of intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R138–R148. [Google Scholar] [CrossRef]
- Daikoku, N.H.; Tyson, J.; Graf, E.; Scott, C.; Smith, R.B.; Johnson, J.W.C.; King, T.M. The relative significance of human placental lactogen in the diagnosis of retarded fetal growth. Am. J. Obstet. Gynecol. 1979, 135, 516–521. [Google Scholar] [CrossRef]
- Lindberg, B.S.; Nilsson, B.A. Human placental lactogen (HPL) levels in abnormal pregnancies. Br. J. Obstet. Gynaecol. 1973, 80, 1046–1053. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; The National Academies Press: Washington, DC, USA, 2007.
- Kappes, S.M.; Warren, W.C.; Pratt, S.L.; Liang, R.; Anthony, R.V. Quantification and cellular localization of ovine placental lactogen messenger ribonucleic acid expression during mid- and late gestation. Endocrinology 1992, 131, 2829–2838. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.E.; Brown, L.D.; Thorn, S.R.; Limesand, S.W.; Davis, M.; Hay, W.W.; Rozance, P.J. Increased adrenergic signaling is responsible for decreased glucose-stimulated insulin secretion in the chronically hyperinsulinemic ovine fetus. Endocrinology 2015, 156, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, J.S.; Culpepper, C.B.; Brown, L.D.; Wesolowski, S.R.; Jonker, S.S.; Davis, M.A.; Limesand, S.W.; Wilkening, R.B.; Hay, W.W.; Rozance, P.J. Chronic anemic hypoxemia attenuates glucose-stimulated insulin secretion in fetal sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R492–R500. [Google Scholar] [CrossRef] [PubMed]
- Cilvik, S.N.; Wesolowski, S.R.; Anthony, R.V.; Brown, L.D.; Rozance, P.J. Late gestation fetal hyperglucagonaemia impairs placental function and results in diminished fetal protein accretion and decreased fetal growth. J. Physiol. 2021, 599, 3403–3427. [Google Scholar] [CrossRef] [PubMed]
- Ghidini, A. Idiopathic Fetal Growth Restriction. Obstet. Gynecol. Surv. 1996, 51, 376–382. [Google Scholar] [CrossRef]
- Alexander, I.; Anthony, F.; Letchworth, A.T. Placental protein profile and glucose studies in a normal pregnancy with ex-tremely low levels of human placental lactogen. Case report. Br. J. Obstet. Gynaecol. 1982, 89, 241–243. [Google Scholar] [CrossRef]
- Barbeiri, F.; Botticelli, A.; Consarino, R.; Genazzani, A.R.; Volpe, A. Failure of placenta to produce hPL in an otherwise un-eventful pregnancy: A case report. Biol. Res. Pregnancy Perinatol. 1986, 7, 131–133. [Google Scholar]
- Borody, I.B.; Carlton, M.A. Isolated defect in human placental lactogen synthesis in a normal pregnancy. Case report. Br. J. Obstet. Gynaecol. 1981, 88, 447–449. [Google Scholar] [CrossRef]
- Rygaard, K.; Revol, A.; Esquivel-Escobedo, D.; Beck, B.L.; Barrera-Saldaña, H.A. Absence of human placental lactogen and placental growth hormone (HGH-V) during pregnancy: PCR analysis of the deletion. Hum. Genet. 1998, 102, 87–92. [Google Scholar] [CrossRef]
- Sideri, M.; De Virgiliis, G.; Guidobono, F.; Borgese, N.; Sereni, L.P.; Nicolini, U.; Remotti, G. Immunologically undetectable human placental lactogen in a normal pregnancy. Br. J. Obstet. Gynaecol. 1983, 90, 771–773. [Google Scholar] [CrossRef]
- Simon, P.; Decoster, C.; Brocas, H.; Schwers, J.; Vassart, G. Absence of human chorionic somatomammotropin during pregnancy associated with two types of gene deletion. Hum. Genet. 1986, 74, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.M.; Regnault, T.R.H.; Limesand, S.W.; Hay, W.W., Jr.; Anthony, R.V. Investigating the causes of low birth weights in contrasting ovine paradigms. J. Physiol. 2005, 565, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.L. Sheep models of intrauterine growth restriction: Fetal adaptations and consequences. Clin. Exp. Pharmacol. Physiol. 2008, 35, 730–743. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Forhead, A.J. Endocrine regulation of fetal metabolism towards term. Dom. Anim. Endocrinol. 2022, 78, 106657. [Google Scholar] [CrossRef]
- Fowden, A.L.; Forhead, A.J. Endocrine mechanisms of intrauterine programming. Reproduction 2004, 127, 515–526. [Google Scholar] [CrossRef]
- Taylor, M.J.; Jenkin, G.; Robinson, J.S.; Thorburn, G.D.; Friesen, H.; Chan, J.S. Concentrations of placental lactogen in chron-ically catheterized ewes and fetuses in late pregnancy. J. Endocrinol. 1980, 85, 27–34. [Google Scholar] [CrossRef]
- Schoknecht, P.A.; Currie, W.B.; Bell, A.W. Kinetics of placental lactogen in mid- and late-gestation ovine fetuses. J. Endocrinol. 1992, 133, 95–100. [Google Scholar] [CrossRef]
- Freemark, M.; Keen, A.; Fowlkes, J.; Mularoni, T.; Comer, M.; Grandis, A.; Kodack, L. The placental lactogen receptor in maternal and fetal sheep liver: Regulation by glucose and role in the pathogenesis of fasting during pregnancy. Endocrinology 1992, 130, 1063–1070. [Google Scholar] [CrossRef]
- Lynch, C.S.; Kennedy, V.C.; Tanner, A.R.; Ali, A.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. Impact of Placental SLC2A3 Deficiency during the First-Half of Gestation. Int. J. Mol. Sci. 2022, 23, 12530. [Google Scholar] [CrossRef]
- Hay, W.W.; Sparks, J.W.; Battaglia, F.C.; Meschia, G. Maternal-fetal glucose exchange: Necessity of a three-pool model. Am. J. Physiol. Endocrinol. Metab. 1984, 246, E528–E534. [Google Scholar] [CrossRef]
- Schoknecht, P.A.; McGuire, M.A.; Cohick, W.S.; Currie, W.B.; Bell, A.W. Effect of chronic infusion of placental lactogen on ovine fetal growth in late gestation. Domest. Anim. Endocrinol. 1996, 13, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Baumann, M.U.; Schneider, H.; Malek, A.; Palta, V.; Surbek, D.V.; Sager, R.; Zamudio, S.; Illsley, N.P. Regulation of Human Trophoblast GLUT1 Glucose Transporter by Insulin-Like Growth Factor I (IGF-I). PLoS ONE 2014, 9, e106037. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.N.; Crombleholme, T.; Habli, M. Adenoviral-mediated placental gene transfer of IGF-1 corrects placental insufficiency via enhanced placental glucose transport mechanisms. PLoS ONE 2013, 8, e74632. [Google Scholar] [CrossRef] [PubMed]
- Wali, J.A.; De Boo, H.A.; Derraik, J.G.B.; Phua, H.H.; Oliver, M.H.; Bloomfield, F.H.; Harding, J.E. Weekly Intra-Amniotic IGF-1 Treatment Increases Growth of Growth-Restricted Ovine Fetuses and Up-Regulates Placental Amino Acid Transporters. PLoS ONE 2012, 7, e37899. [Google Scholar] [CrossRef]
- Oliver, M.H.; Harding, J.E.; Breier, B.H.; Evans, P.C.; Gluckman, P.D. Glucose but Not a Mixed Amino Acid Infusion Regulates Plasma Insulin-Like Growth Factor-I Concentrations in Fetal Sheep. Pediatr. Res. 1993, 34, 62–65. [Google Scholar] [CrossRef]
- Zhou, J.; Bondy, C. Insulin-like growth factor-II and its binding proteins in placental development. Endocrinology 1992, 131, 1230–1240. [Google Scholar] [CrossRef]
- Khandwala, H.M.; McCutcheon, I.E.; Flyvbjerg, A.; Friend, K.E. The effects of insulin-like growth factors on tumorigenesis and neoplastic growth. Endocr. Rev. 2000, 21, 215–244. [Google Scholar] [CrossRef]
- Pollak, M.N.; Schernhammer, E.S.; Hankinson, S.E. Insulin-like growth factors and neoplasia. Nat. Rev. Cancer 2004, 4, 505–518. [Google Scholar] [CrossRef]
- Scott, C.D.; Firth, S.M. The role of the M6P/IGF-II receptor in cancer: Tumor suppression or garbage disposal? Horm. Metab. Res. 2004, 36, 261–271. [Google Scholar]
- Kiess, W.; Haskell, J.F.; Lee, L.; Greenstein, L.A.; Miller, B.E.; Aarons, A.L.; Rechler, M.M.; Nissley, S.P. An antibody that blocks insulin-like growth factor (IGF) binding to the type II IGF receptor is neither an agonist nor an inhibitor of IGF-stimulated biologic responses in L6 myoblasts. J. Biol. Chem. 1987, 262, 12745–12751. [Google Scholar] [CrossRef]
- Nolan, C.M.; Kyle, J.W.; Watanabe, H.; Sly, W.S. Binding of insulin-like growth factor II (IGF-II) by human cation-independent mannose 6-phosphate receptor/IGF-II receptor expressed in receptor-deficient mouse L cells. Cell Regul. 1990, 1, 197–213. [Google Scholar] [CrossRef]
- Kornfeld, S. Structure and function of the mannose 6-phosphate/insulin-like growth factor II receptors. Annu. Rev. Biochem. 1992, 61, 307–330. [Google Scholar] [CrossRef]
- Clemmons, D.R. Use of mutagenesis to probe IGF-binding protein structure/function relationships. Endocr. Rev. 2001, 22, 800–817. [Google Scholar] [CrossRef] [PubMed]
- Firth, S.M.; Baxter, R. Cellular Actions of the Insulin-Like Growth Factor Binding Proteins. Endocr. Rev. 2002, 23, 824–854. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.I.; Clemmons, D.R. Insulin-Like Growth Factors and Their Binding Proteins: Biological Actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar] [CrossRef]
- Carrick, F.E.; Forbes, B.E.; Wallace, J.C. BIAcore analysis of bovine insulin-like growth factor (IGF)-binding protein-2 identifies major IGF binding site determinants in both the amino- and carboxyl-terminal domains. J. Biol. Chem. 2001, 276, 27120–27128. [Google Scholar] [CrossRef] [PubMed]
- Höflich, A.; Lahm, H.; Blum, W.; Kolb, H.; Wolf, E. Insulin-like growth factor-binding protein-2 inhibits proliferation of human embryonic kidney fibroblasts and of IGF-responsive colon carcinoma cell lines. FEBS Lett. 1998, 434, 329–334. [Google Scholar] [CrossRef]
- Hoeflich, A.; Fettscher, O.; Lahm, H.; Blum, W.F.; Kolb, H.J.; Engelhardt, D.; Wolf, E.; Weber, M.M. Overexpression of insulin-like growth factor-binding protein-2 results in increased tumorigenic potential in Y-1 adrenocortical tumor cells. Cancer Res. 2000, 60, 834–838. [Google Scholar]
- Oh, Y. Igf-independent regulation of breast cancer growth by IGF binding proteins. Breast Cancer Res. Treat. 1998, 47, 283–293. [Google Scholar] [CrossRef]
- Badinga, L.; Song, S.; Simmen, R.; Clarke, J.; Clemmons, D.R.; Simmen, F. Complex mediation of uterine endometrial epithelial cell growth by insulin-like growth factor-II (IGF-II) and IGF-binding protein-2. J. Mol. Endocrinol. 1999, 23, 277–285. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide | Sequence (5′–3′) |
|---|---|
| Control (NTS) shRNA sense | GAGTTAAAGGTTCGGCACGAATTCAAGAGATTCGTGCCGAACCTTTAACTC |
| CSH-targeting shRNA sense | AAGGCCAAAGTACTTGTAGACTTCAAGAGAGTCTACAAGTACTTTGGCCTT |
| cDNA | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Anneal, °C | Product, bp |
|---|---|---|---|---|
| RPS15 | atcattctgcccgagatggtg | tgctttacgggcttgtaggtg | 58 | 124 |
| IGF1 | tcgcatctctcttctatctggccctgt | acagtacatctccagcctcctcaga | 62 | 238 |
| IGF2 | gaccgcggcttctacttcag | aagaacttgcccacggggtat | 62 | 202 |
| IGFBP1 | tgatgaccgagtccagtgag | gtccagcgaagtctcacac | 62 | 247 |
| IGFBP2 | caatggcgaggagcactctg | tggggatgtgtagggaatag | 55 | 330 |
| IGFBP3 | ctcagagcacagacaccca | ggcatatttgagctccac | 54 | 335 |
| IGF1R | aactgtcatctccaacctc | caagcctcccactatcaac | 60 | 493 |
| IGF2R | gacttgtgtccagaccagattc | gccgtcgtcctcactctcatc | 58 | 674 |
| Variable | Experimental Group | p-Value | % Change | |
|---|---|---|---|---|
| NTS RNAi | CSH RNAi PI-FGR | |||
| Fetal weight, g | 4625.6 ± 37.1 | 3226.4 ± 325.8 | ≤0.01 | 30.2 |
| Fetal liver weight, g | 125.2 ± 3.32 | 74.2 ± 9.04 | ≤0.01 | 40.7 |
| Crown-rump length, cm | 51.4 ± 0.71 | 46.9 ± 1.37 | 0.02 | 8.76 |
| Placental weight, g | 517.7 ± 28.6 | 247.9 ± 20.5 | ≤0.01 | 52.1 |
| Total placentome number | 70.5 ± 5.86 | 58.0 ± 1.79 | 0.09 | 17.7 |
| Uterine blood flow, mL/min | 2477.6 ± 501.9 | 1453.7 ±403.9 | 0.03 | 41.3 |
| Umbilical blood flow, mL/min | 854.6 ± 41.3 | 496.9 ± 170.1 | ≤0.01 | 41.9 |
| Uterine glucose uptake, μmol/min | 423.0 ± 84.6 | 266.9 ± 75.2 | 0.04 | 36.9 |
| Umbilical glucose uptake, μmol/min | 137.5 ± 9.85 | 78.4 ± 28.5 | 0.02 | 43.0 |
| Uteroplacental glucose utilization, μmol/min | 292.7 ± 62.0 | 188.1 ± 75.2 | 0.17 | 35.7 |
| Uterine oxygen uptake, mmol/min | 3.01 ± 0.56 | 1.79 ± 0.49 | ≤0.01 | 40.3 |
| Umbilical oxygen uptake, mmol/min | 1.53 ± 0.09 | 0.93 ± 0.33 | 0.02 | 39.5 |
| Uteroplacental oxygen utilization, mmol/min | 1.53 ± 0.30 | 0.94 ± 0.35 | 0.06 | 38.4 |
| Variable | Experimental Group | p-Value | % Change | |
|---|---|---|---|---|
| NTS RNAi | CSH RNAi PI-FGR | |||
| Uterine Vein CSH, ng/mL | 371.2 ± 64.6 | 165.3 ± 49.5 | 0.05 | 55.5 |
| Umbilical Vein CSH, ng/mL | 32.0 ± 3.28 | 31.1 ± 6.88 | 0.89 | 2.86 |
| Uterine Artery Insulin, ng/mL | 0.54 ± 0.14 | 0.67 ± 0.19 | 0.60 | 22.5 |
| Umbilical Artery Insulin, ng/mL | 0.73 ± 0.10 | 0.35 ± 0.08 | 0.02 | 52.9 |
| Uterine Artery IGF1, ng/mL | 170.7 ± 13.8 | 151.9 ± 14.2 | 0.39 | 11.0 |
| Umbilical Artery IGF1, ng/mL | 167.5 ± 23.8 | 72.0 ± 20.13 | 0.02 | 57.0 |
| Uterine Artery Cortisol, ng/mL | 159.5 ± 62.2 | 121.3 ± 59.7 | 0.67 | 23.9 |
| Umbilical Artery Cortisol, ng/mL | 51.4 ± 4.66 | 129.2 ± 28.0 | 0.01 | 151.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hord, T.K.; Tanner, A.R.; Kennedy, V.C.; Lynch, C.S.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. Impact of Chorionic Somatomammotropin In Vivo RNA Interference Phenotype on Uteroplacental Expression of the IGF Axis. Life 2023, 13, 1261. https://doi.org/10.3390/life13061261
Hord TK, Tanner AR, Kennedy VC, Lynch CS, Winger QA, Rozance PJ, Anthony RV. Impact of Chorionic Somatomammotropin In Vivo RNA Interference Phenotype on Uteroplacental Expression of the IGF Axis. Life. 2023; 13(6):1261. https://doi.org/10.3390/life13061261
Chicago/Turabian StyleHord, Taylor K., Amelia R. Tanner, Victoria C. Kennedy, Cameron S. Lynch, Quinton A. Winger, Paul J. Rozance, and Russell V. Anthony. 2023. "Impact of Chorionic Somatomammotropin In Vivo RNA Interference Phenotype on Uteroplacental Expression of the IGF Axis" Life 13, no. 6: 1261. https://doi.org/10.3390/life13061261