Imaging Plate Autoradiography for Ingested Anthropogenic Cesium-137 in Butterfly Bodies: Implications for the Biological Impacts of the Fukushima Nuclear Accident


Abstract
:1. Introduction
2. Materials and Methods
2.1. Butterflies Reared with a 137Cs-Containing Artificial Diet
2.2. Butterflies Collected from Contaminated Fields
2.3. Imaging Plate Exposure
2.4. Exposure Periods
2.5. Data Acquisition and Analyses
2.6. Statistical Analyses
3. Results
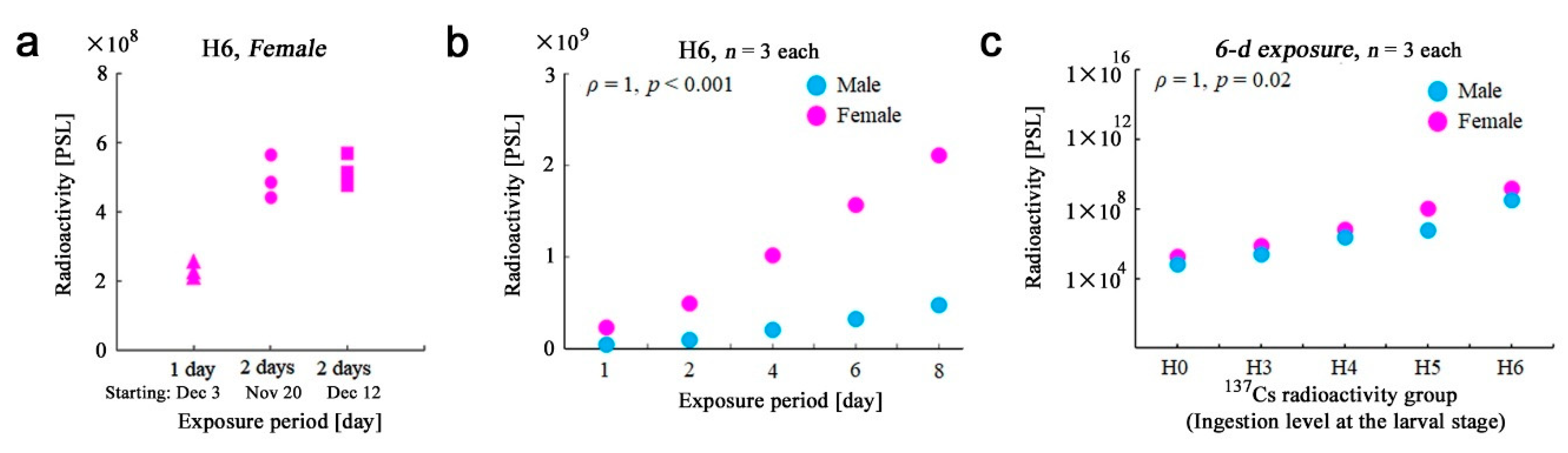
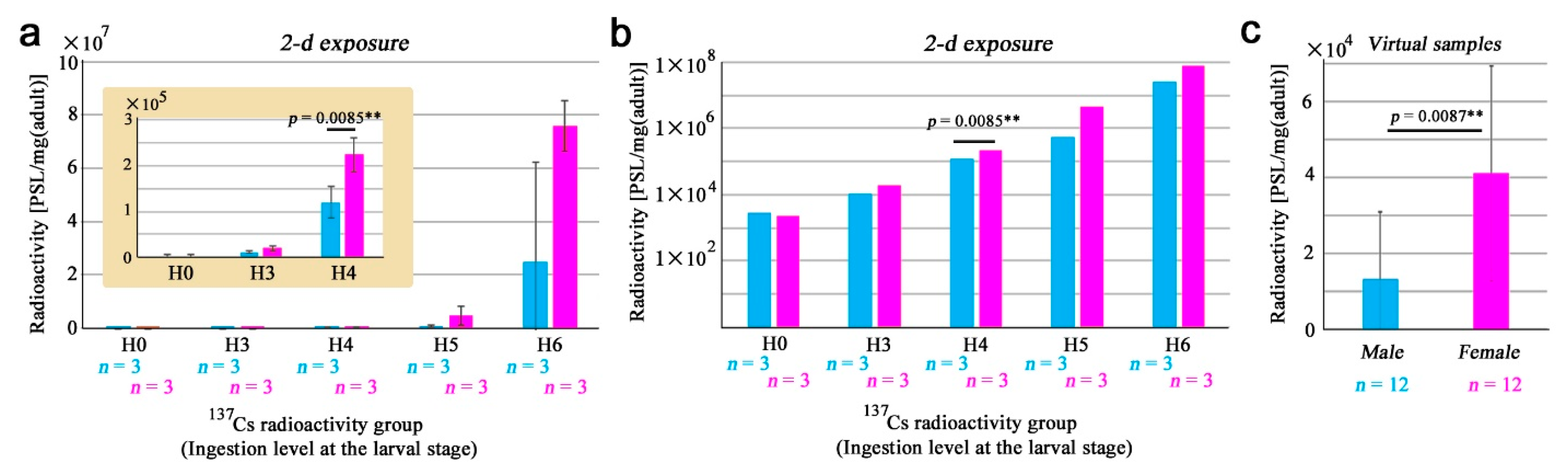
3.1. Adult Whole-Body Samples
3.2. Sex Differences in the Whole-Body Samples
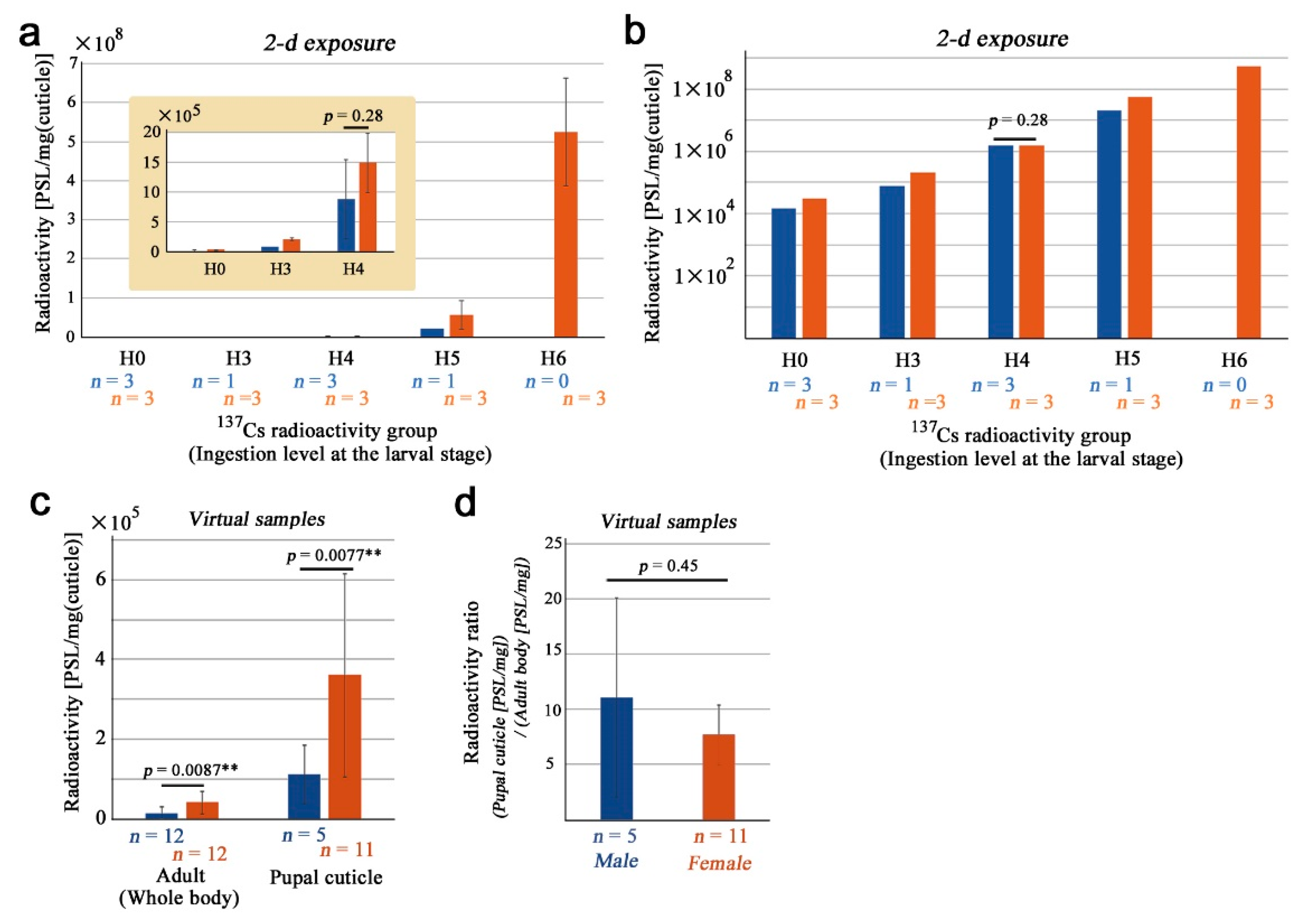
3.3. Pupal Cuticles and Their Sex Differences
3.4. Pupae
3.5. Body Parts of Adults
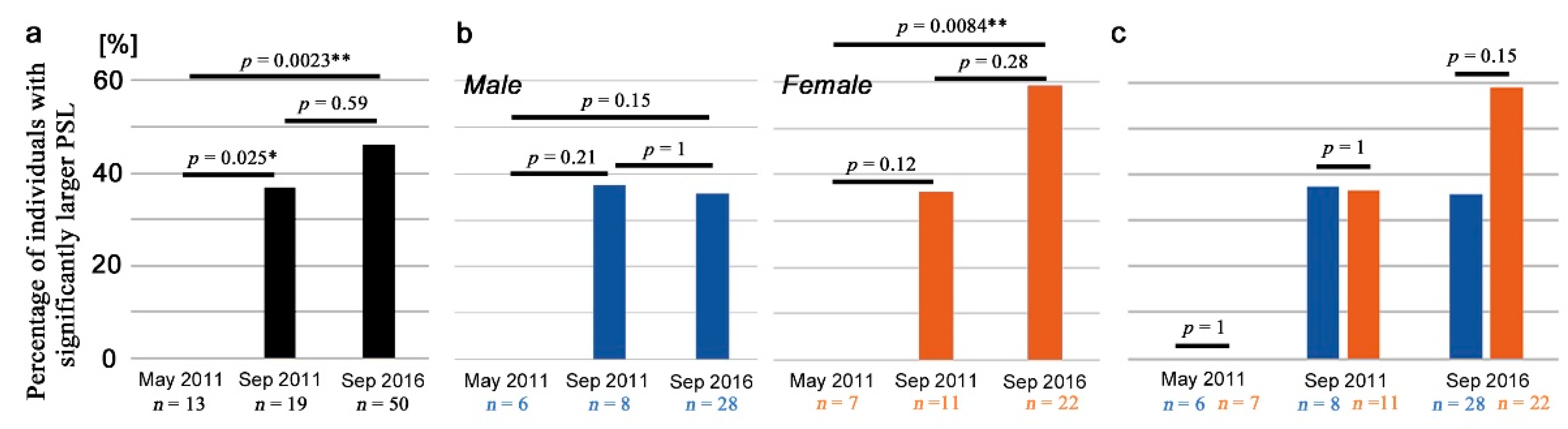
3.6. Field-Caught Adults
4. Discussion
4.1. Significance of This Study
4.2. Female Bias and Accumulation in Body Parts
4.3. No Condensation in Butterflies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Relationship between PSL and Becquerel
Appendix B. Detection Limit of Imaging Plate Autoradiography Used in This Study
Appendix C. Relationship between the Whole Body and Body Parts
Appendix D. Feasibility for Detecting 137Cs in Field Samples
Appendix E. Assimilation Rate and Adult Retention Rate
References
- Chino, M.; Nakayama, H.; Nagai, H.; Terada, H.; Katata, G.; Yamazawa, H. Preliminary estimation of released amount of 131I and 137Cs accidentally discharged from the Fukushima Daiichi nuclear power plant into the atmosphere. J. Nucl. Sci. Technol. 2011, 48, 1129–1134. [Google Scholar] [CrossRef]
- Kinoshita, N.; Sucki, K.; Sasa, K.; Kitagawa, J.; Ikarashi, S.; Nishimura, T.; Wong, Y.S.; Satou, Y.; Handa, K.; Takahashi, T.; et al. Assessment of individual radionuclide distributions from the Fukushima nuclear accident covering central-east Japan. Proc. Natl. Acad. Sci. USA 2011, 108, 19526–19529. [Google Scholar] [CrossRef] [PubMed]
- Hirose, K. 2011 Fukushima Dai-ichi nuclear power plant accident: Summary of regional radioactive deposition monitoring results. J. Environ. Radioact. 2012, 111, 13–17. [Google Scholar] [CrossRef]
- Torii, T.; Sugita, T.; Okada, C.E.; Reed, M.S.; Blumenthal, D.J. Enhanced analysis methods to derive the spatial distribution of 131I deposition on the ground by airborne surveys at an early stage after the Fukushima Daiichi nuclear power plant accident. Health Phys. 2013, 105, 192–200. [Google Scholar] [CrossRef]
- Le Petit, G.; Douysset, G.; Ducros, G.; Gross, P.; Achim, P.; Monfort, M.; Raymond, P.; Pontillon, Y.; Jutier, C.; Blanchard, X. Analysis of radionuclide releases from the Fukushima Dai-ichi nuclear power plant accident Part I. Pure Appl. Geophys. 2014, 171, 629–644. [Google Scholar] [CrossRef]
- Achim, P.; Monfort, M.; Le Pettit, G.; Gross, P.; Douysset, G.; Taffary, T.; Blanchard, X.; Moulin, C. Analysis of radionuclide releases from the Fukushima Dai-ichi nuclear power plant accident Part II. Pure Appl. Geophys. 2014, 171, 645–667. [Google Scholar] [CrossRef]
- Ministry of the Environment. Chapter 4. Radiation Protection. 4.4 Long-Term Effects. Posted on 31 March 2016. English Version. Available online: https://www.env.go.jp/en/chemi/rhm/basic-info/1st/04-04-08.html (accessed on 23 June 2022).
- Lokobauer, N.; Franić, Z.; Bauman, A.; Maračić, M.; Cesar, D.; Senčar, J. Radiation contamination after the Chernobyl nuclear accident and the effective dose received by the population of Croatia. J. Environ. Radioact. 1998, 41, 137–146. [Google Scholar] [CrossRef]
- Arapis, G.D.; Karandinos, M.G. Migration of 137Cs in the soil of sloping semi-natural ecosystems in Northern Greece. J. Environ. Radioact. 2004, 77, 133–142. [Google Scholar] [CrossRef]
- Tahir, S.N.A.; Jamil, K.; Zaidi, J.H.; Arif, M.; Ahmed, N. Activity concentration of 137Cs in soil samples from Punjab province (Pakistan) and estimation of gamma-ray dose rate for external exposure. Radiat. Prot. Dosim. 2006, 118, 345–351. [Google Scholar] [CrossRef]
- Ambrosino, F.; Stellato, L.; Sabbarese, C. A case study on possible radiological contamination in the Lo Uttara landfill site (Caserta, Italy). J. Phys. Conf. Ser. 2020, 1548, 012001. [Google Scholar] [CrossRef]
- Endo, S.; Kimura, S.; Takatsuji, T.; Nanasawa, K.; Imanaka, T.; Shizuma, K. Measurement of soil contamination by radionuclides due to the Fukushima Dai-ichi Nuclear Ppower Plant accident and associated estimated cumulative external dose estimation. J. Environ. Radioact. 2021, 111, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P.; Hagiwara, A.; Matsui, S.; Kasahara, S.; Kawatsu, K.; Nishiumi, I.; Suzuki, H.; Mousseau, T.A. Abundance of birds in Fukushima as judges from Chernobyl. Environ. Pollut. 2012, 164, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Murase, K.; Murase, J.; Horie, R.; Endo, K. Effects of the Fukushima Daiichi nuclear accident on goshawk reproduction. Sci. Rep. 2015, 5, 9405. [Google Scholar] [CrossRef] [PubMed]
- Bonisoli-Alquati, A.; Koyama, K.; Tedeschi, D.J.; Kitamura, W.; Sukuzi, H.; Ostermiller, S.; Arai, E.; Møller, A.P.; Mousseau, T.A. Abundance and genetic damage of barn swallows from Fukushima. Sci. Rep. 2015, 5, 9432. [Google Scholar] [CrossRef]
- Hiyama, A.; Nohara, C.; Kinjo, S.; Taira, W.; Gima, S.; Tanahara, A.; Otaki, J.M. The biological impacts of the Fukushima nuclear accident on the pale grass blue butterfly. Sci. Rep. 2012, 2, 570. [Google Scholar] [CrossRef] [PubMed]
- Hiyama, A.; Nohara, C.; Taira, W.; Kinjo, S.; Iwata, M.; Otaki, J.M. The Fukushima nuclear accident and the pale grass blue butterfly: Evaluating biological effects of long-term low-dose exposures. BMC Evol. Biol. 2013, 13, 168. [Google Scholar] [CrossRef]
- Akimoto, S. Morphological abnormalities in gall-forming aphids in a radiation-contaminated area near Fukushima Daiichi: Selective impact of fallout? Ecol. Evol. 2014, 4, 355–369. [Google Scholar] [CrossRef]
- Akimoto, S.I.; Li, Y.; Imanaka, T.; Sato, H.; Ishida, K. Effects of radiation from contaminated soil and moss in Fukushima on embryogenesis and egg hatching of the aphid Prociphilus oriens. J. Hered. 2018, 109, 199–205. [Google Scholar] [CrossRef]
- Taira, W.; Toki, M.; Kakinohana, K.; Sakauchi, K.; Otaki, J.M. Developmental and hemocytological effects of ingesting Fukushima’s radiocesium on the cabbage white butterfly Pieris rapae. Sci. Rep. 2019, 9, 2625. [Google Scholar] [CrossRef]
- Ochiai, K.; Hayama, S.; Nakiri, S.; Nakanishi, S.; Ishii, N.; Uno, T.; Kato, T.; Konno, F.; Kawamoto, Y.; Tsuchida, S.; et al. Low blood cell counts in wild Japanese monkeys after the Fukushima Daiichi nuclear disaster. Sci. Rep. 2014, 4, 5793. [Google Scholar] [CrossRef]
- Hayama, S.; Tsuchiya, M.; Ochiaki, K.; Nakiri, S.; Nakanishi, S.; Ishii, N.; Kato, T.; Tanaka, A.; Konno, F.; Kawamoto, Y.; et al. Small head size and delayed body weight growth in wild Japanese monkey fetuses after the Fukushima Daiichi nuclear disaster. Sci. Rep. 2017, 7, 3528. [Google Scholar] [CrossRef]
- Urushihara, Y.; Suzuki, T.; Shimizu, Y.; Ohtaki, M.; Kuwahara, Y.; Suzuki, M.; Uno, T.; Fujita, S.; Saito, A.; Yamashiro, H.; et al. Haematological analysis of Japanese macaques (Macaca fuscata) in the area affected by the Fukushima Daiichi Nuclear Power Plant accident. Sci. Rep. 2018, 8, 16748. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, T.; Yoshii, H.; Mizuno, S.; Shiraishi, H. Decline in intertidal biota after the 2011 Great East Japan Earthquake and Tsunami and the Fukushima nuclear disaster: Field observations. Sci. Rep. 2016, 6, 20416. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, G.; Shibato, J.; Imanaka, T.; Cho, K.; Kubo, A.; Kikuchi, S.; Satoh, K.; Kimura, S.; Ozawa, S.; Fukutani, S.; et al. Unraveling low-level gamma radiation-responsive changes in expression of early and late genes in leaves of rice seedlings at Iitate Village, Fukushima. J. Hered. 2014, 105, 723–738. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Ichikawa, S.; Kubota, M.; Hoshino, J.; Kubota, Y.; Maruyama, K.; Fuma, S.; Kawaguchi, I.; Yoschenko, V.I.; Yoshida, S. Morphological defects in native Japanese fir trees around the Fukushima Daiichi Nuclear Power Plant. Sci. Rep. 2015, 5, 13232. [Google Scholar] [CrossRef] [PubMed]
- Yoschenko, V.; Nanba, K.; Yoshida, S.; Watanabe, Y.; Takase, T.; Sato, N.; Keitoku, K. Morphological abnormalities in Japanese red pine (Pinus densiflora) at the territories contaminated as a result of the accident at Fukushima Dai-ichi Nuclear Power Plant. J. Environ. Radioact. 2016, 165, 60–67. [Google Scholar] [CrossRef]
- Shirôzu, T. The Standard of Butterflies in Japan; Gakken: Tokyo, Japan, 2006. (In Japanese) [Google Scholar]
- Yata, O. Iconographia Insectorum Japonicorum Colore Naturali Esita Vol. I; Hokuryukan: Tokyo, Japan, 2007. (In Japanese) [Google Scholar]
- Oda, H.; Kitazoe, N. Observation Encyclopedia of Lycaenid Butterflies; Kaisei-sha: Tokyo, Japan, 2002. (In Japanese) [Google Scholar]
- Hiyama, A.; Iwata, M.; Otaki, J.M. Rearing the pale grass blue Zizeeria maha (Lepidoptera, Lycaenidae): Toward the establishment of a lycaenid model system for butterfly physiology and genetics. Entomol. Sci. 2010, 13, 293–302. [Google Scholar] [CrossRef]
- Otaki, J.M.; Hiyama, A.; Iwata, M.; Kudo, T. Phenotypic plasticity in the range-margin population of the lycaenid butterfly Zizeeria maha. BMC Evol. Biol. 2010, 10, 252. [Google Scholar] [CrossRef]
- Hiyama, A.; Taira, W.; Nohara, C.; Iwasaki, M.; Kinjo, S.; Iwata, M.; Otaki, J.M. Spatiotemporal abnormality dynamics of the pale grass blue butterfly: Three years of monitoring (2011–2013) after the Fukushima nuclear accident. BMC Evol. Biol. 2015, 15, 15. [Google Scholar] [CrossRef]
- Sakauchi, K.; Taira, W.; Hiyama, A.; Imanaka, T.; Otaki, J.M. The pale grass blue butterfly in ex-evacuation zones 5.5 years after the Fukushima nuclear accident: Contributions of initial high-dose exposure to transgenerational effects. J. Asia-Pac. Entomol. 2020, 23, 242–252. [Google Scholar] [CrossRef]
- Taira, W.; Iwasaki, M.; Otaki, J.M. Body size distributions of the pale grass blue butterfly in Japan: Size rules and the status of the Fukushima population. Sci. Rep. 2015, 5, 12351. [Google Scholar] [CrossRef] [PubMed]
- Hiyama, A.; Taira, W.; Iwasaki, M.; Sakauchi, K.; Gurung, R.; Otaki, J.M. Geographical distribution of morphological abnormalities and wing color pattern modifications of the pale grass blue butterfly in northeastern Japan. Entomol. Sci. 2017, 20, 100–110. [Google Scholar] [CrossRef]
- Hiyama, A.; Taira, W.; Iwasaki, M.; Sakauchi, K.; Iwata, M.; Otaki, J.M. Morphological abnormality rate of the pale grass blue butterfly Zizeeria maha (Lepidoptera: Lycaenidae) in southwestern Japan: A reference data set for environmental monitoring. J. Asia-Pac. Entomol. 2017, 20, 1333–1339. [Google Scholar] [CrossRef]
- Sakauchi, K.; Taira, W.; Toki, M.; Iraha, Y.; Otaki, J.M. Overwintering states of the pale grass blue butterfly Zizeeria maha (Lepidoptera: Lycaenidae) at the time of the Fukushima nuclear accident in March 2011. Insects 2019, 10, 389. [Google Scholar] [CrossRef] [PubMed]
- Nohara, C.; Hiyama, A.; Taira, W.; Tanahara, A.; Otaki, J.M. The biological impacts of ingested radioactive materials on the pale grass blue butterfly. Sci. Rep. 2014, 4, 4946. [Google Scholar] [CrossRef] [PubMed]
- Nohara, C.; Taira, W.; Hiyama, A.; Tanahara, A.; Takatsuji, T.; Otaki, J.M. Ingestion of radioactively contaminated diets for two generations in the pale grass blue butterfly. BMC Evol. Biol. 2014, 14, 193. [Google Scholar] [CrossRef]
- Taira, W.; Hiyama, A.; Nohara, C.; Sakauchi, K.; Otaki, J.M. Ingestional and transgenerational effects of the Fukushima nuclear accident on the pale grass blue butterfly. J. Radiat. Res. 2015, 56, i2–i18. [Google Scholar] [CrossRef]
- Nohara, C.; Hiyama, A.; Taira, W.; Otaki, J.M. Robustness and radiation resistance of the pale grass blue butterfly from radioactively contaminated areas: A possible case of adaptive evolution. J. Hered. 2018, 109, 188–198. [Google Scholar] [CrossRef]
- Otaki, J.M. Fukushima’s lessons from the blue butterfly: A risk assessment of the human living environment in the post-Fukushima era. Integr. Environ. Assess. Manag. 2016, 12, 667–672. [Google Scholar] [CrossRef]
- Otaki, J.M.; Taira, W. Current status of the blue butterfly in Fukushima research. J. Hered. 2018, 109, 178–187. [Google Scholar] [CrossRef]
- Otaki, J.M. The pale grass blue butterfly as an indicator for the biological effect of the Fukushima Daiichi Nuclear Power Plant accident. In Low-Dose Radiation Effects on Animals and Ecosystems; Fukumoto, M., Ed.; Springer: Singapore, 2020; pp. 239–247. [Google Scholar] [CrossRef]
- Otaki, J.M.; Sakauchi, K.; Taira, W. The second decade of the blue butterfly in Fukushima: Untangling the ecological field effects after the Fukushima nuclear accident. Integr. Environ. Assess. Manag. 2022, 18, 1539–1550. [Google Scholar] [CrossRef] [PubMed]
- Takada, N.; Yamauchi, E.; Fujimoto, H.; Banno, Y.; Tsuchida, K.; Hashido, K.; Nakajima, Y.; Tu, Z.; Takahashi, M.; Fujii, H.; et al. A novel indicator for radiation sensitivity using the wing size reduction of Bombyx mori pupae caused by γ-ray irradiation. J. Insect Biotechnol. Sericol. 2006, 75, 161–165. [Google Scholar] [CrossRef]
- Ahmad, S.; Hussain, A.; Ullah, F.; Jamil, M.; Ali, A.; Ali, S.; Luo, Y. 60Co-γ radiation alters developmental stages of Zeugodacus cucurbitae (Diptera: Tephritidae) through apoptosis pathways gene expression. J. Insect Sci. 2021, 21, 16. [Google Scholar] [CrossRef]
- Gurung, R.D.; Taira, W.; Sakauchi, K.; Iwata, M.; Hiyama, A.; Otaki, J.M. Tolerance of high oral doses of nonradioactive and radioactive caesium chloride in the pale grass blue butterfly Zizeeria maha. Insects 2019, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Kinouchi, T.; Fujii, T.; Imanaka, T.; Takahashi, T.; Fukutani, S.; Maki, D.; Nohtomi, A.; Takahashi, S. Observation of morphological abnormalities in silkworm pupae after feeding 137CsCl-supplemented diet to evaluate the effects of low dose-rate exposure. Sci. Rep. 2020, 10, 16055. [Google Scholar] [CrossRef] [PubMed]
- Otaki, J.M. Understanding low-dose exposure and field effects to resolve the field-laboratory paradox: Multifaceted biological effects from the Fukushima nuclear accident. In New Trends in Nuclear Science; Awwad, N.S., AlFaify, S.A., Eds.; IntechOpen: London, UK, 2018; pp. 49–71. [Google Scholar] [CrossRef]
- Sakauchi, K.; Taira, W.; Toki, M.; Tsuhako, M.; Umetsu, K.; Otaki, J.M. Nutrient imbalance of the host plant for larvae of the pale grass blue butterfly may mediate the field effect of low-dose radiation exposure in Fukushima: Dose-dependent changes in the sodium content. Insects 2021, 12, 149. [Google Scholar] [CrossRef]
- Sakauchi, K.; Taira, W.; Otaki, J.M. Metabolomic response of the creeping wood sorel Oxalis corniculata to low-dose radiation exposure from Fukushima’s contaminated soil. Life 2021, 11, 990. [Google Scholar] [CrossRef]
- Sakauchi, K.; Taira, W.; Otaki, J.M. Metabolomic profiles of the creeping wood sorel Oxalis corniculata in radioactively contaminated fields in Fukushima: Dose-dependent changes in key metabolites. Life 2022, 12, 115. [Google Scholar] [CrossRef]
- Morita, A.; Sakauchi, K.; Taira, W.; Otaki, J.M. Ingestional toxicity of radiation-dependent metabolites of the host plant for the pale grass blue butterfly: A mechanism of field effects of radioactive pollution in Fukushima. Life 2022, 12, 615. [Google Scholar] [CrossRef]
- Fukuda, T.; Kino, Y.; Abe, Y.; Yamashiro, H.; Kuwahara, Y.; Nihei, H.; Sano, Y.; Irisawa, A.; Shimura, T.; Obata, Y.; et al. Distribution of artificial radionuclides in abandoned cattle in the evacuation zone of the Fukushima Daiichi nuclear power plant. PLoS ONE 2013, 8, e54312. [Google Scholar] [CrossRef]
- Sato, I.; Okada, K.; Sasaki, J.; Chida, H.; Satoh, H.; Miura, K.; Kikuchi, K.; Otani, K.; Sato, S. Distribution of radioactive cesium and stable cesium in cattle kept on a highly contaminated area of Fukushima nuclear accident. Anim. Sci. J. 2015, 86, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Fuma, S.; Kubota, Y.; Ihara, S.; Takahashi, H.; Watanabe, Y.; Aono, T.; Soeda, H.; Yoshida, S. Radiocaesium contamination of wild boars in Fukushima and surrounding regions after the Fukushima nuclear accident. J. Environ. Radioact. 2016, 164, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Tanoi, K.; Uchida, K.; Doi, C.; Nihei, N.; Hirose, A.; Kobayashi, N.I.; Sugita, R.; Nobori, T.; Nakanishi, T.M.; Kanno, M.; et al. Investigation of radiocesium distribution in organs of wild boar grown in Iitate, Fukushima after the Fukushima Daiichi nuclear power plant accident. J. Radioanal. Nucl. Chem. 2016, 307, 741–746. [Google Scholar] [CrossRef]
- Saito, R.; Nemoto, Y.; Tsukada, H. Relationship between radiocaesium in muscle and physicochemical fractions of radiocaesium in the stomach of wild boar. Sci. Rep. 2020, 10, 6796. [Google Scholar] [CrossRef]
- Morimoto, M.; Kobayashi, J.; Kino, Y. Radiation dose and gene expression analysis of wild boar 10 years after the Fukushima Daiichi Nuclear Plant accident. Sci. Rep. 2022, 12, 18653. [Google Scholar] [CrossRef]
- Hayama, S.; Nakiri, S.; Nakanishi, S.; Ishii, N.; Uno, T.; Kato, T.; Konno, F.; Kawamoto, Y.; Tsuchida, S.; Ochiai, K.; et al. Concentration of radiocesium in the wild Japanese manokey (Macaca fuscata) over the first 15 months after the Fukushima Daiichi nuclear disaster. PLoS ONE 2013, 8, e68530. [Google Scholar] [CrossRef]
- Omi, T.; Nakiri, S.; Nakanishi, S.; Ishii, N.; Uno, T.; Konno, F.; Inagaki, T.; Sakamoto, A.; Shito, M.; Udagawa, C.; et al. Concentrations of 137Cs radiocaesium in the organs and tissues of low-dose-exposed wild Japanese monkeys. BMC Res. Notes 2020, 13, 121. [Google Scholar] [CrossRef]
- Tsuboi, J.; Abe, S.; Fujimoto, K.; Kaeriyama, H.; Ambe, D.; Matsuda, K.; Enomoto, M.; Tomiya, A.; Morita, T.; Ono, T.; et al. Exposure of a herbivorous fish to 134Cs and 137Cs from the riverbed following the Fukushima disaster. J. Environ. Radioact. 2015, 141, 32–37. [Google Scholar] [CrossRef]
- Ishii, N.; Furota, T.; Kagami, M.; Tagami, K.; Uchida, S. Inequality in the distribution of 137Cs contamination within freshwater fish bodies and its affecting factors. Sci. Rep. 2021, 11, 5769. [Google Scholar] [CrossRef]
- Yoshimura, M.; Akama, A. Radioactive contamination of aquatic insects in a stream impacted by the Fukushima nuclear power station accident. Hydrobiologia 2014, 722, 19–30. [Google Scholar] [CrossRef]
- Ishii, Y.; Miura, H.; Jo, J.; Tsuji, H.; Saito, R.; Koarai, K.; Hagiwara, H.; Urushidate, T.; Nishikiori, T.; Wada, T.; et al. Radiocesium-bearing microparticles cause a large variation in 137Cs activity concentration in the aquatic insect Stenopsyche marmorata (Tricoptera: Stenopsychidae) in the Ota River, Fukushima, Japan. PLoS ONE 2022, 17, e0268629. [Google Scholar] [CrossRef]
- Iwasa, M.; Nakaya, F.; Kabeya, H.; Sato, K.; Ishikawa, S.-I.; Takahashi, T. Radiocesium concentrations in invertebrates and their environmental media at two distances from the Fukushima Dai-ichi Nuclear Power Plant during 3-6 years after the 2011 accident. Environ. Pollut. 2020, 267, 115657. [Google Scholar] [CrossRef] [PubMed]
- Fuma, S.; Ihara, S.; Takahashi, H.; Inaba, O.; Sato, Y.; Kubota, Y.; Watanabe, Y.; Kawaguchi, I.; Aono, T.; Soeda, H.; et al. Radiocaesium contamination and dose rate estimation of terrestrial and freshwater wildlife in the exclusion zone of the Fukushima Dai-ichi Nuclear Power Plant accident. J. Environ. Radioact. 2017, 171, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Ayabe, Y.; Kanasashi, T.; Hijii, N.; Takenaka, C. Radiocesium contamination of the web spider Nephila clavate (Nephilidae: Arachnida) 1.5 years after the Fukushima Dai-ichi Nuclear Power Plant accident. J. Envion. Radioact. 2014, 127, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Ishii, Y.; Abe, H.; Mitachi, K.; Watanabe, T.; Niizato, T. Translocation of radiocesium released by the Fukushima Daiichi Nuclear Power Plant accident in Japanese chestnut and chestnut weevil larvae. Hortic. J. 2017, 86, 139–144. [Google Scholar] [CrossRef]
- Murakami, M.; Ohte, N.; Suzuki, T.; Ishii, N.; Igarashi, Y.; Tanoi, K. Biological proliferation of cesium-137 through the detrital food chain in a forest ecosystem in Japan. Sci. Rep. 2014, 4, 3599. [Google Scholar] [CrossRef]
- Tanaka, S.; Hatakeyama, K.; Takahashi, S.; Adati, T. Radioactive contamination of arthropods from different trophic levels in hilly and mountainous areas after the Fukushima Daiichi nuclear power plant accident. J. Environ. Radioact. 2016, 164, 104–112. [Google Scholar] [CrossRef]
- Ishii, Y.; Hayashi, S.; Takamura, N. Radiocesium transfer in forest insect communities after the Fukushima Dai-ichi Nuclear Power Plant accident. PLoS ONE 2017, 12, e0171133. [Google Scholar] [CrossRef]
- Iwasa, M.; Sato, K.; Ishikawa, S.; Takahashi, T.; Kabeya, H.; Nakaya, F. Radiocesium contaminations and transfer in cyclorrhaphous flies (Diptera: Muscidae, Calliphoridae) at three distances from the Fukushima Dai-ichi Nuclear Power Plant after the 2011 accident. Appl. Entomol. Zool. 2022, 57, 81–91. [Google Scholar] [CrossRef]
- Nakanishi, H.; Mori, A.; Takeda, K.; Tanaka, H.; Kobayashi, N.; Tanoi, K.; Yamakawa, T.; Mori, S. Discovery of radioactive silver (110mAg) in spiders and other fauna in the terrestrial environment after the meltdown of Fukushima Dai-ichi nuclear power plant. Proc. Jpn. Acad. Ser. B 2015, 91, 160–174. [Google Scholar] [CrossRef]
- Ayabe, Y.; Yoshida, T.; Kanasashi, T.; Hayashi, A.; Fukushi, A.; Hijii, N.; Takenaka, C. Web-building spider Nephila clavate (Nephilidae: Arachnida) can represent 137Cs contamination of arthropod communities and bioavailable 137Cs in forest soil at Fukushima, Japan. Sci. Total Environ. 2019, 687, 1176–1185. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Adati, T.; Takahashi, T.; Fujiwara, K.; Takahashi, S. Concentrations and biological half-life of radioactive cesium in epigeic earthworms after the Fukushima Dai-ichi Nuclear Power Plant accident. J. Environ. Radioact. 2018, 192, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Murakami, H.; Hatta, T.; Kitazawa, H.; Yamada, H.; Yaita, T.; Kogure, T. Speciation of radioactive soil particles in the Fukushima contaminated area by IP autoradiography and microanalyses. Environ. Sci. Technol. 2014, 48, 13053–13059. [Google Scholar] [CrossRef]
- Nakanishi, T.M.; Kobayashi, N.I.; Tanoi, K. Radioactive cesium deposition on rice, wheat, peach tree and soil after nuclear accident in Fukushima. J. Radioanal. Nucl. Chem. 2013, 296, 985–989. [Google Scholar] [CrossRef]
- Nakanishi, T.M. Agricultural implications of the Fukushima nuclear accident. J. Radiat. Res. 2016, 57, i47–i52. [Google Scholar] [CrossRef]
- Furuta, E. Semi-quantitative analysis of leaf surface contamination by radioactivity from the Fukushima Daiichi nuclear power plant accident using HPGe and imaging plate. J. Radioanal. Nucl. Chem. 2013, 297, 337–342. [Google Scholar] [CrossRef]
- Miura, T.; Mimura, M.; Kobayashi, D.; Komiyama, C.; Sekimoto, H.; Miyamoto, M.; Kitamura, A. Radioactive pollution and accumulation of radionuclides in wild plants in Fukushima. J. Plant. Res. 2014, 127, 5–10. [Google Scholar] [CrossRef]
- Ikka, T.; Nishina, Y.; Kamoshita, M.; Oya, Y.; Okuno, K.; Morita, A. Radiocesium uptake through leaf surfaces of tea plants (Camellia sinensis L.). J. Environ. Radioact. 2018, 182, 70–73. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Sawano, K.; Furuhama, K.; Mori, C.; Yamada, K. An autoradiogram of skeletal muscle from a pig raised on a farm within 20 km of the Fukushima Daiichi Nuclear Power Plant. J. Vet. Med. Sci. 2013, 75, 93–94. [Google Scholar] [CrossRef]
- Mousseau, T.A.; Dingle, H. Maternal effects in insect life histories. Annu. Rev. Entomol. 1991, 36, 511–534. [Google Scholar] [CrossRef]
- Mousseau, T.; Fox, C.W. The adaptive significance of maternal effects. Trends Ecol. Evol. 1998, 13, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Uller, T. Developmental plasticity and the evolution of parental effects. Trends Ecol. Evol. 2008, 23, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Woestmann, L.; Saastamoinen, M. The importance of trans-generational effects in Lepidoptera. Curr. Zool. 2016, 62, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Yabu, S.; Otaki, J.M. The plastic larval body color of the pale grass blue butterfly Zizeeria maha (Lepidoptera: Lycaenidae) in response to the host plant color: The maternal effect on crypsis. Insects 2023, 14, 202. [Google Scholar] [CrossRef]
- Hancock, S.; Vo, N.T.K.; Omar-Nazir, L.; Batlle, J.V.I.; Otaki, J.M.; Hiyama, A.; Byun, S.H.; Seymour, C.B.; Mothersill, C. Transgenerational effects of historic radiation dose in pale grass blue butterflies around Fukushima following the Fukushima Dai-ichi Nuclear Power Plant meltdown accident. Environ. Res. 2019, 168, 230–240. [Google Scholar] [CrossRef]
- Strathie, L.W.; Nicolson, S.W. Post-eclosion diuresis in a flightless insect, the silkmoth Bombyx mori. Physiol. Entomol. 1993, 18, 435–439. [Google Scholar] [CrossRef]
- Inoue, T.A.; Ito, T.; Hagiya, H.; Hata, T.; Asaoka, K.; Yokohari, F.; Niihara, K. K+ excretion: The other purpose for puddling behavior in Japanese Papilio butterflies. PLoS ONE 2015, 10, e0126632. [Google Scholar] [CrossRef]
- Sakauchi, K.; Taira, W.; Otaki, J.M. Instruction, Table, Picture Sheet, and Original Pictures: Morphological Abnormalities in the Field Samples of the Pale Grass Blue Butterfly for Three Years after the Fukushima Nuclear Accident. Figshare Collection. 2023. Available online: https://figshare.com/collections/Instruction_table_picture_sheet_and_original_pictures_Morphological_abnormalities_in_the_field_samples_of_the_pale_grass_blue_butterfly_for_three_years_after_the_Fukushima_nuclear_accident/6425981 (accessed on 11 March 2023).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Month and Year | May 2011 | September 2011 | September 2016 | |||
|---|---|---|---|---|---|---|
| Sex | Male | Female | Male | Female | Male | Female |
| Number of individuals examined | 6 | 7 | 8 | 11 | 28 | 22 |
| Number of positive individuals with significantly larger PSL values *2 | 0 | 0 | 3 | 4 | 10 | 13 |
| Percentage of positive individuals with significantly larger PSL values | 0% | 0% | 37.5% | 36.4% | 35.7% | 59.1% |
| Mean PSL ± SD *3 | na | na | 16,338 ± 6939 | 17,450 ± 2899 | 15,144 ± 3394 | 14,961 ± 4731 |
| Maximum value of mean PSL | na | na | 23,206 ± 8217 | 19,858 ± 9993 | 20,750 ± 8266 | 24,891 ± 5086 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakauchi, K.; Otaki, J.M. Imaging Plate Autoradiography for Ingested Anthropogenic Cesium-137 in Butterfly Bodies: Implications for the Biological Impacts of the Fukushima Nuclear Accident. Life 2023, 13, 1211. https://doi.org/10.3390/life13051211
Sakauchi K, Otaki JM. Imaging Plate Autoradiography for Ingested Anthropogenic Cesium-137 in Butterfly Bodies: Implications for the Biological Impacts of the Fukushima Nuclear Accident. Life. 2023; 13(5):1211. https://doi.org/10.3390/life13051211
Chicago/Turabian StyleSakauchi, Ko, and Joji M. Otaki. 2023. "Imaging Plate Autoradiography for Ingested Anthropogenic Cesium-137 in Butterfly Bodies: Implications for the Biological Impacts of the Fukushima Nuclear Accident" Life 13, no. 5: 1211. https://doi.org/10.3390/life13051211