


Chromosome Painting in Cercopithecus petaurista (Schreber, 1774) Compared to Other Monkeys of the Cercopithecini Tribe (Catarrhini, Primates)

Abstract
:1. Introduction
2. Materials and Methods
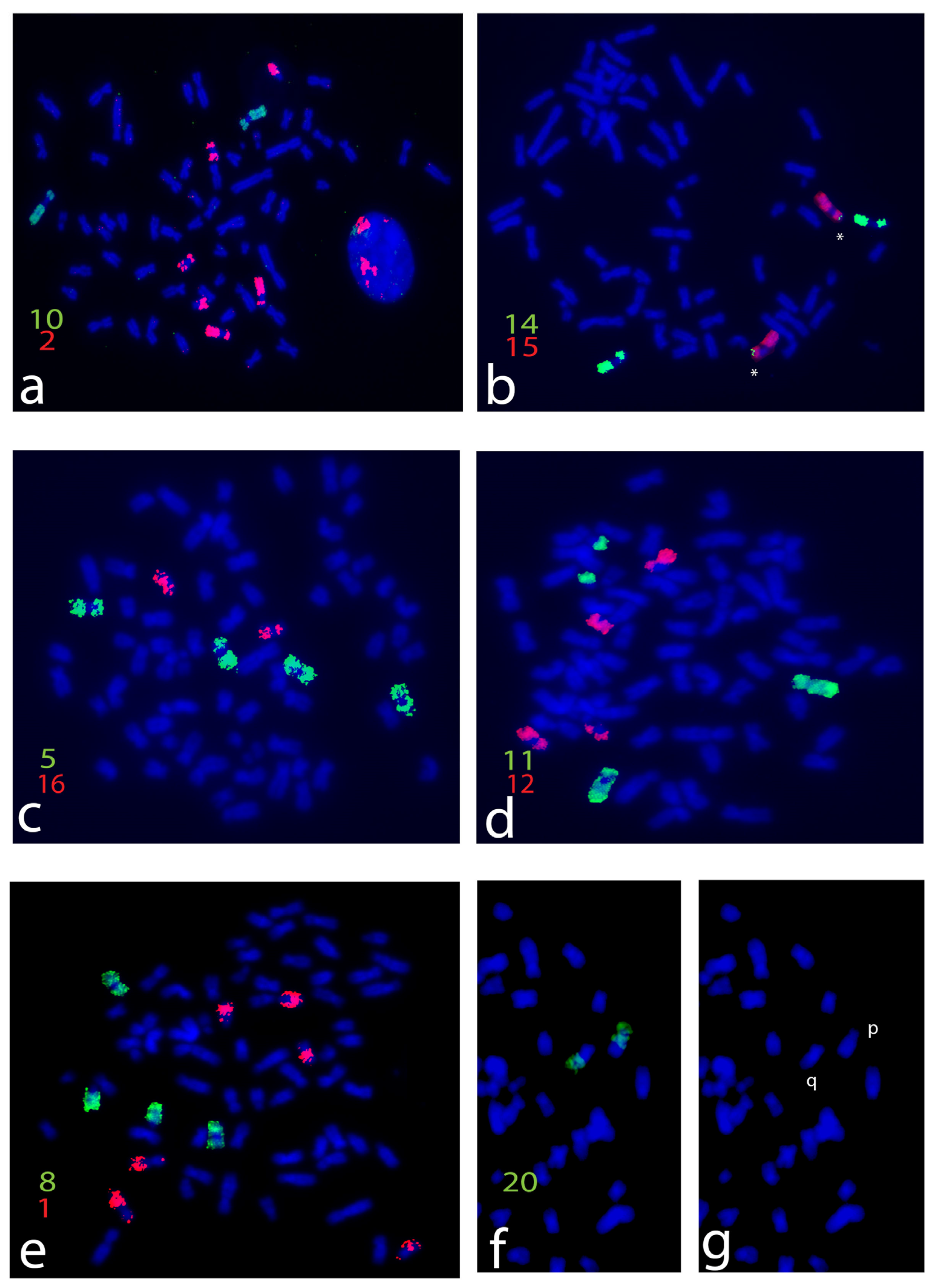
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilbert, C.C.; Bibi, F.; Hill, A.; Beech, M.J. Early guenon from the late Miocene Baynunah Formation, Abu Dhabi, with implications for cercopithecoid biogeography and evolution. Proc. Natl. Acad. Sci. USA 2014, 111, 10119–10124. [Google Scholar] [CrossRef]
- Butynski, T.M. The guenons: An overview of diversity and taxonomy. In The Guenons: Diversity and Adaptation in African Monkeys; Glenn, M.E., Cords, M., Eds.; Kluwer Academic Publishers: New York, NY, USA, 2002; pp. 3–13. [Google Scholar]
- Rusell, A.; Mittermeier, A.B.; Rylands, D.E.W. Handbook of the Mammals of the World; Primates Lynx Edicions: Barcelona, Spain, 2013; Volume 3, ISBN 978-84-96553-89-7. [Google Scholar]
- Lo Bianco, S.; Masters, J.; Sineo, L. The evolution of the cercopithecine: A (post)modern synthesis. Evol. Anthropol. 2017, 26, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Dutrillaux, B.; Muleris, M.; Couturier, J. Chromosomal evolution of Cercopithecinae. In A Primate Radiation: Evolutionary Biology of the African Guenons; Gautier-Hion, A., Bourlière, F., Gautier, J.-P., Eds.; Cambridge University Press: Cambridge, UK, 1988; pp. 150–159. [Google Scholar]
- Dutrillaux, B. Chromosomal evolution in primates: Tentative phylogeny from Microcebus murinus (Prosimian) to man. Hum. Genet. 1979, 48, 251–314. [Google Scholar] [CrossRef] [PubMed]
- Stanyon, R.; Bruening, R.; Stone, G.; Shearin, A.; Bigoni, F. Reciprocal painting between humans, De Brazza’s and patas monkeys reveals a major bifurcation in the Cercopithecini phylogenetic tree. Cytogenet. Genome Res. 2005, 108, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Tosi, A.J.; Detwiler, K.M.; Disotell, T.R. X-chromosomal window into the evolutionary history of the guenons (Primates: Cercopithecini). Mol. Phylogenetics Evol. 2005, 36, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Tosi, A.J.; Melnick, D.J.; Disotell, T.R. Sex chromosome phylogenetics indicate a single transition to terrestriality in the guenons (tribe Cercopithecini). J. Hum. Evol. 2004, 46, 223–237. [Google Scholar] [CrossRef]
- Xing, J.; Wang, H.; Zhang, Y.; Ray, D.A.; Tosi, A.J.; Disotell, T.R.; Batzer, M.A. A mobile element-based evolutionary history of guenons (tribe Cercopithecini). BMC Biol. 2007, 5, 5. [Google Scholar] [CrossRef]
- Cardini, A.; Elton, S. Variation in guenon skulls (I): Species divergence, ecological and genetic differences. J. Hum. Evol. 2008, 54, 615–637. [Google Scholar] [CrossRef]
- Martin, R.D.; McLarnon, A.M. Quantitative comparisons of the skull and teeth in Guenons. In A Primate Radiation: Evolutionary Biology of the African Guenons; Gautier-Hion, A., Ed.; Cambridge University Press: Cambridge, UK, 1988; pp. 160–183. [Google Scholar]
- Dumas, F.; Sineo, L. Chromosomal dynamics in Cercopithecini studied by Williams-Beuren probe mapping. Caryologia 2010, 63, 435–442. [Google Scholar]
- Sineo, L. The Still under Construction Cercopithecinae Phylogeny. J. Primatol. 2012, 1, 2. [Google Scholar] [CrossRef]
- Sineo, L. The banded karyotype of Cercopithecus mitis maesi compared with the karyotypes of C. albogularis samango and C. nictitans stampflii. I. J. Primatol. 1990, 11, 541–552. [Google Scholar]
- Sineo, L.; Lo Bianco, S.; Picone, B. Evidence of a chromosomal polymorphism unique to Cercopithecini. A key factor in the Tribe definition. J. Primatol. 2015, 4. [Google Scholar] [CrossRef]
- Groves, C.P. Primate Taxonomy; Smithsonian Institution Press: Washington, DC, USA, 2001. [Google Scholar]
- Grubb, P.; Butynski, T.M.; Oates, J.F.; Bearder, S.K.; Disotell, T.R.; Groves, C.P.; Struhsaker, T.T. Assessment of the diversity of African primates. Int. J. Primatol. 2003, 24, 1301–1357. [Google Scholar] [CrossRef]
- Takahashi, K.; Terai, Y.; Nishida, M.; Okada, N. Phylogenetic relationships and ancient incomplete lineage sorting among cichlid fishes in Lake Tanganyika as revealed by analysis of the insertion of retroposons. Mol. Biol. Evol. 2001, 18, 2057–2066. [Google Scholar] [CrossRef] [PubMed]
- Detwiler, K.M. Hybridization between Red-tailed Monkeys (Cercopithecus ascanius) and Blue Monkeys (C. mitis) in East African Forests. In The Guenons: Diversity and Adaptation in African Monkeys; Glenn, M.E., Cords, M., Eds.; Kluwer Academic Publishers: New York, NY, USA, 2002; p. 79. [Google Scholar]
- Detwiler, K.M. Mitochondrial DNA Analyses of Cercopithecus Monkeys Reveal a Localized Hybrid Origin for C. mitis doggetti in Gombe National Park, Tanzania. Int. J. Primatol. 2019, 40, 28–52. [Google Scholar] [CrossRef]
- De Jong, Y.A.; Butynski, T.M. Three sykes’s monkey Cercopithecus mitis× vervet monkey Chlorocebus pygerythrus hybrids in Kenya. Primate Conserv. 2010, 25, 43–56. [Google Scholar] [CrossRef]
- Furo, I.D.O.; Kretschmer, R.; O’Brien, P.C.; Pereira, J.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Phylogenetic analysis and karyotype evolution in two species of core Gruiformes: Aramides cajaneus and Psophia viridis. Genes 2020, 11, 307. [Google Scholar] [CrossRef]
- Farias, J.C.; Santos, N.; Bezerra, D.P.; Sotero-Caio, C.G. Chromosome Painting in Lonchorhina aurita Sheds Light onto the Controversial Phylogenetic Position of Sword-Nosed Bats (Chiroptera, Phyllostomidae). Cytogenet. Genome Res. 2021, 161, 569–577. [Google Scholar] [CrossRef]
- Knytl, M.; Smolík, O.; Kubíčková, S.; Tlapáková, T.; Evans, B.J.; Krylov, V. Chromosome divergence during evolution of the tetraploid clawed frogs, Xenopus mellotropicalis and Xenopus epitropicalis as revealed by Zoo-FISH. PLoS ONE 2017, 12, e0177087. [Google Scholar] [CrossRef]
- Dumas, F.; Sineo, L.; Ishida, T. Taxonomic identification of Aotus (Platyrrhinae) through cytogenetics, Identificazione tassonomica di Aotus (Platyrrhinae) mediante la citogenetica. J. Biol. Res. Boll. Della Soc. Ital. Biol. Sper. 2015, 88, 65–66. [Google Scholar]
- Dumas, F.; Sineo, L. The evolution of human synteny 4 by mapping sub-chromosomal specific probes in Primates. Caryologia 2014, 67, 281–291. [Google Scholar] [CrossRef]
- Wienberg, J.; Stanyon, R.; Jauch, A.; Cremer, T. Homologies in human and Macaca fuscata chromosomes revealed by in situ suppression hybridization with human chromosome specific DNA libraries. Chromosoma 1992, 101, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Wienberg, J. Fluorescence in situ hybridization to chromosomes as a tool to understand human and primate genome evolution. Cytogenet. Genome Res. 2004, 108, 139–160. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.J.; Menninger, J.C.; Nash, W.G. Atlas of Mammalian Chromosomes; Wiley-Liss: Hoboken, NJ, USA, 2006. [Google Scholar]
- Moulin, S.; Gerbault-Seureau, M.; Dutrillaux, B.; Richard, F.A. Phylogenomics of African guenons. Chromosome Res. 2008, 16, 783–799. [Google Scholar] [CrossRef] [PubMed]
- Stanyon, R.; Rocchi, M.; Bigoni, F.; Archidiacono, N. Evolutionary molecular cytogenetics of catarrhine primates: Past, present and future. Cytogenet. Genome Res. 2012, 137, 273–284. [Google Scholar] [CrossRef]
- Tolomeo, D.; Capozzi, O.; Chiatante, G.; Sineo, L.; Ishida, T.; Archidiacono, N.; Stanyon, R. Eight million years of maintained heterozygosity in chromosome homologs of cercopithecine monkeys. Chromosoma 2020, 129, 57–67. [Google Scholar] [CrossRef]
- Finelli, P.; Stanyon, R.; Plesker, R.; Ferguson-Smith, M.A.; O’brien, P.; Wienberg, J. Reciprocal chromosome painting shows that the great difference in diploid number between human and African green monkey is mostly due to non-Robertsonian fissions. Mamm. Genome 1999, 10, 713–718. [Google Scholar] [CrossRef]
- Finelli, P. Analisi citogenetica sulla filogenesi delle catarrhinae tramite ibridazione in situ interspecifiche. In Dissertation; Università Degli Studi di Bari: Bari, Italy, 1996. [Google Scholar]
- Ventura, M.; Antonacci, F.; Cardone, M.F.; Stanyon, R.; D’Addabbo, P.; Cellamare, A.; Sprague, L.J.; Eichler, E.E.; Archidiacono, N.; Rocchi, M. Evolutionary formation of new centromeres in macaque. Science 2007, 316, 243–246. [Google Scholar] [CrossRef]
- Ruiz Herrera, A.; Garcia, F.; Azzalin, C.; Giuliotto, E.; Egozucue JPonsa’, F.M.; Garcia, M. Distribution of intrachromosomal telomeric sequences (ITS) on Macaca fascicularis (Primates) chromosomes and their implication for chromosome evolution. Hum. Genet. 2002, 110, 578–586. [Google Scholar] [CrossRef]
- Dumas, F.; Cuttaia, I.; Sineo, L. Chromosomal distribution of interstitial telomeric sequences in nine neotropical primates (Platyrrhini): Possible implications in evolution and phylogeny. J. Zool. Syst. Evol. Res. 2016, 54, 226–236. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Schillaci, O.; Sineo, L.; Dumas, F. Distribution of interstitial telomeric sequences in primates and the pygmy tree shrew (Scandentia). Cytogenet. Genome Res. 2017, 151, 141–150. [Google Scholar] [CrossRef]
- Ceraulo, S.; Perelman, P.L.; Mazzoleni, S.; Rovatsos, M.; Dumas, F. Repetitive sequence distribution on Saguinus, Leontocebus and Leontopithecus tamarins (Platyrrhine, Primates) by mapping telomeric (TTAGGG) motifs and rDNA loci. Biology 2021, 10, 844. [Google Scholar] [CrossRef]
- Dumas, F.; Perelman, P.L.; Biltueva, L.; Roelke-Parker, M.E. Retrotransposon mapping in spider monkey genomes of the family Atelidae (Platyrrhini, Primates) shows a high level of LINE-1 amplification. J. Biol. Res. Boll. Della Soc. Ital. Biol. Sper. 2022, 95, 10725. [Google Scholar] [CrossRef]
- Dumas, F.; Houck, M.L.; Bigoni, F.; Perelman, P.; Romanenko, S.A.; Stanyon, R. Chromosome painting of the pygmy tree shrew shows that no derived cytogenetic traits link primates and scandentia. Cytogenet. Genome Res. 2012, 136, 175–179. [Google Scholar]
- Ceraulo, S.; Milioto, V.; Dumas, F. Centromeric enrichment of LINE-1 retrotransposon in two species of South American monkeys Alouatta belzebul and Ateles nancymaae (Platyrrhini, Primates). Caryologia 2021, 74, 111–119. [Google Scholar] [CrossRef]
- Ceraulo, S.; Perelman, P.L.; Dumas, F. Massive LINE-1 retrotransposon enrichment in tamarins of the Cebidae family (Platyrrhini, Primates) and its significance for genome evolution. J. Zool. Syst. Evol. Res. 2021, 59, 2553–2561. [Google Scholar] [CrossRef]
- Fernàndez, R.; Barragàn, M.; Bullejos, M.; Marchal, J.; Diaz de la Guardia, R.; Sanchez, A. New C-band protocol by heat denaturation in the presence of formamide. Hereditas 2002, 137, 145–148. [Google Scholar] [CrossRef]
- Stanyon, R.; Sineo, L. Citotassonomia e filogenesi del genere Cercopithecus. Antropol. Contemp. 1983, 6, 237–252. [Google Scholar]
- Stanyon, R.; Rocchi, M.; Capozzi, O.; Roberto, R.; Misceo, D.; Ventura, M.; Archidiacono, N. Primate chromosome evolution: Ancestral karyotypes, marker order and neocentromeres. Chromosome Res. 2008, 16, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Graphodatsky, A.S.; Trifonov, V.A.; Stanyon, R. The genome diversity and karyotype evolution of mammals. Mol. Cytogenet. 2011, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gifalli-Iughetti, C.; Koiffmann, C.P. Synteny of human chromosomes 14 and 15 in the platyrrhines (Primates, Platyrrhini). Genet. Mol. Biol. 2009, 32, 786–791. [Google Scholar] [CrossRef]
- Capozzi, O.; Archidiacono, N.; Lorusso, N.; Stanyon, R.; Rocchi, M. The 14/15 association as a paradigmatic example of tracing karyotype evolution in New World monkeys. Chromosoma 2016, 125, 747–756. [Google Scholar] [CrossRef]
- Romagno, D.; Chiarelli, B.; Sineo, L. Studio Dell’evoluzione dei Cromosomi Dell’uomo Attraverso il Mappaggio di” loci” Specifici Nei Primati Non Umani: Il Caso Del Cromosoma 15. Antropo. 2001; pp. 45–52. Available online: https://dialnet.unirioja.es/servlet/articulo?codigo=4749731 (accessed on 21 February 2023).
- Scardino, R.; Milioto, V.; Proskuryakova, A.A.; Serdyukova, N.A.; Perelman, P.L.; Dumas, F. Evolution of the human chromosome 13 synteny: Evolutionary rearrangements, plasticity, human disease genes and cancer breakpoints. Genes 2020, 11, 383. [Google Scholar] [CrossRef]
- Robinson, T.J.; Ruiz-Herrera, A.; Avise, J.C. Hemiplasy and homoplasy in the karyotypic phylogenies of mammals. Proc. Natl. Acad. Sci. USA 2008, 105, 14477–14481. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | EPA (2n = 54) | CAE (2n = 60) | CPE (2n = 66) | CER (2n = 66) | CNE (2n = 62) | CNI (2n = 70) | CWO (2n = 72) |
|---|---|---|---|---|---|---|---|
| HSA paints | |||||||
| 1 | 1 | 2 | 3 | 3 | 4 | 3 | 4 |
| 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 |
| 3 | 2 | 2 | 3 | 3 | 3 | 3 | 3 |
| 4 | 1 | 2 | 1 | 1 | 1 | 2 | 3 |
| 5 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| 6 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| 7 | 2 | 2 | 1 | 1 | 1 | 2 | 2 |
| 8 | 1 | 1 | 2 | 2 | 1 (with 1) | 2 | 1 |
| 9 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 10 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 11 | 1 | 1 | 2 | 2 | 1 | 2 | 2 |
| 12 | 1 | 1 | 2 | 2 | 1 | 2 | 2 |
| 13 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 14 | 1 | 2 | 2 | 2 | 2 | 2 | 2 |
| 14/15 | + | + | + | + | + | + | + |
| 15 | 2 | 2 | 1 | 1 | 2 | 1 | 1 |
| 16 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 17 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 18 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 19 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 20 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 20/21 | + | + | + | + | + | + | + |
| 21 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 22 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milioto, V.; Sineo, L.; Dumas, F. Chromosome Painting in Cercopithecus petaurista (Schreber, 1774) Compared to Other Monkeys of the Cercopithecini Tribe (Catarrhini, Primates). Life 2023, 13, 1203. https://doi.org/10.3390/life13051203
Milioto V, Sineo L, Dumas F. Chromosome Painting in Cercopithecus petaurista (Schreber, 1774) Compared to Other Monkeys of the Cercopithecini Tribe (Catarrhini, Primates). Life. 2023; 13(5):1203. https://doi.org/10.3390/life13051203
Chicago/Turabian StyleMilioto, Vanessa, Luca Sineo, and Francesca Dumas. 2023. "Chromosome Painting in Cercopithecus petaurista (Schreber, 1774) Compared to Other Monkeys of the Cercopithecini Tribe (Catarrhini, Primates)" Life 13, no. 5: 1203. https://doi.org/10.3390/life13051203