
UVA/UVB Irradiation Exerts a Distinct Phototoxic Effect on Human Keratinocytes Compared to Human Malignant Melanoma Cells
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Equipment
2.2. Cell Lines and Cell Culture
2.3. Cell Irradiation with UVA, UVB, and UVA/UVB
2.4. Cell Viability Assessment
2.5. Evaluation of Cell Morphology and Confluence
2.6. Immunofluorescence Staining of Cellular Components
2.7. RT-qPCR Analysis
2.8. RealTime-Glo™ Annexin V Apoptosis and Necrosis Assay
2.9. Statistical Analysis
3. Results
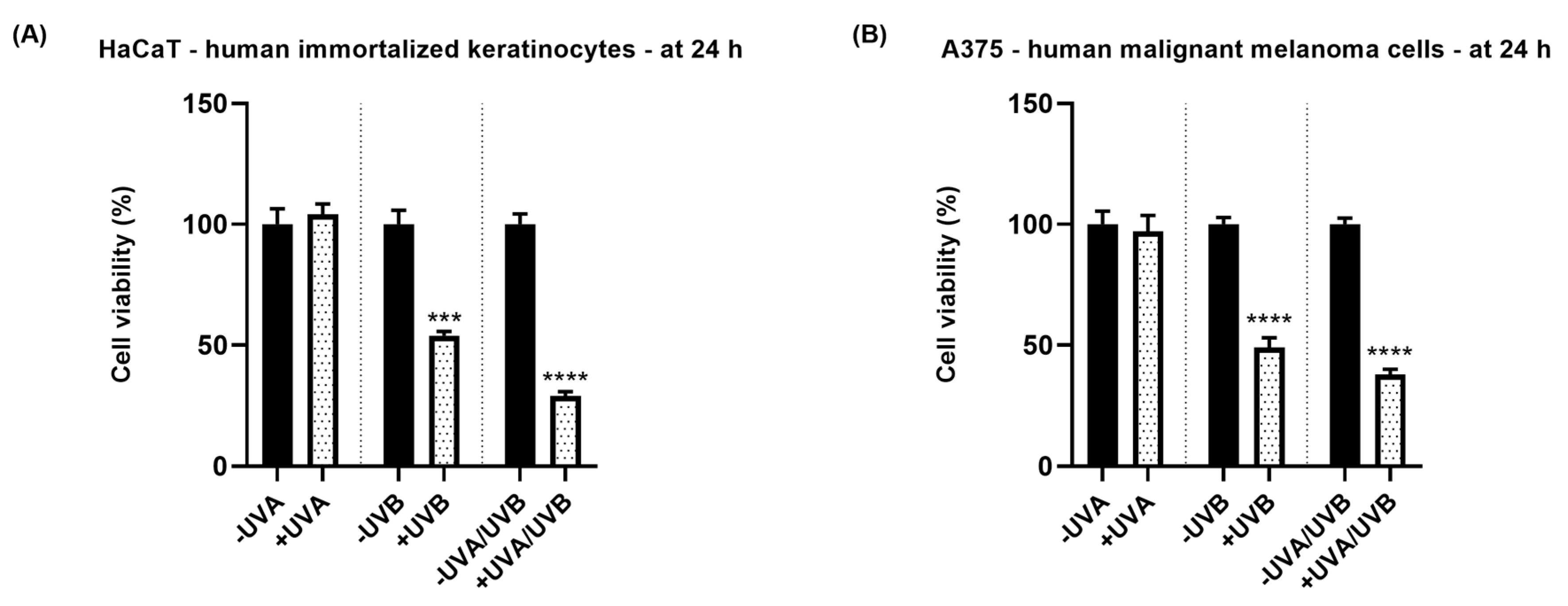
3.1. Cell Viability Assessment
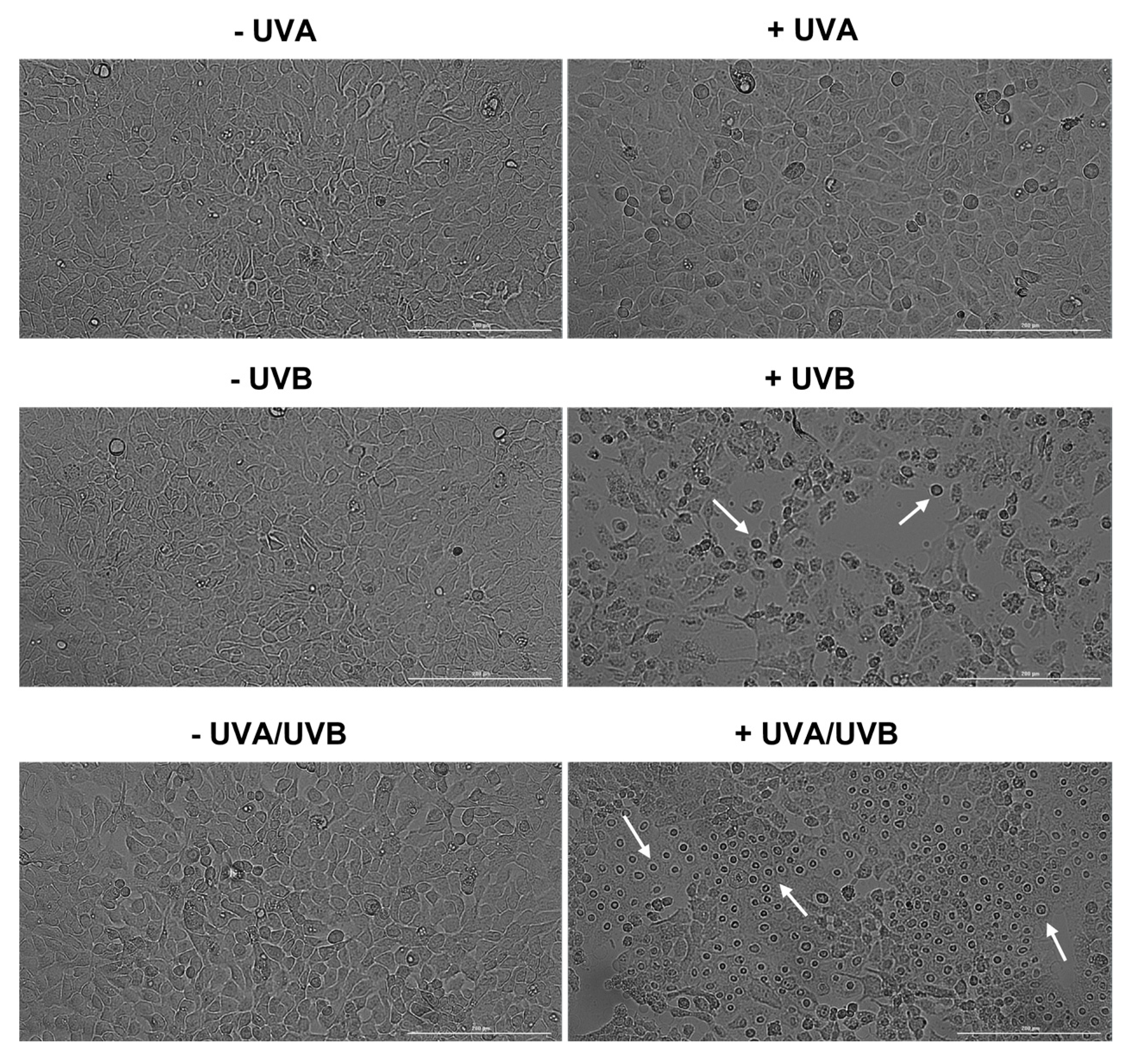
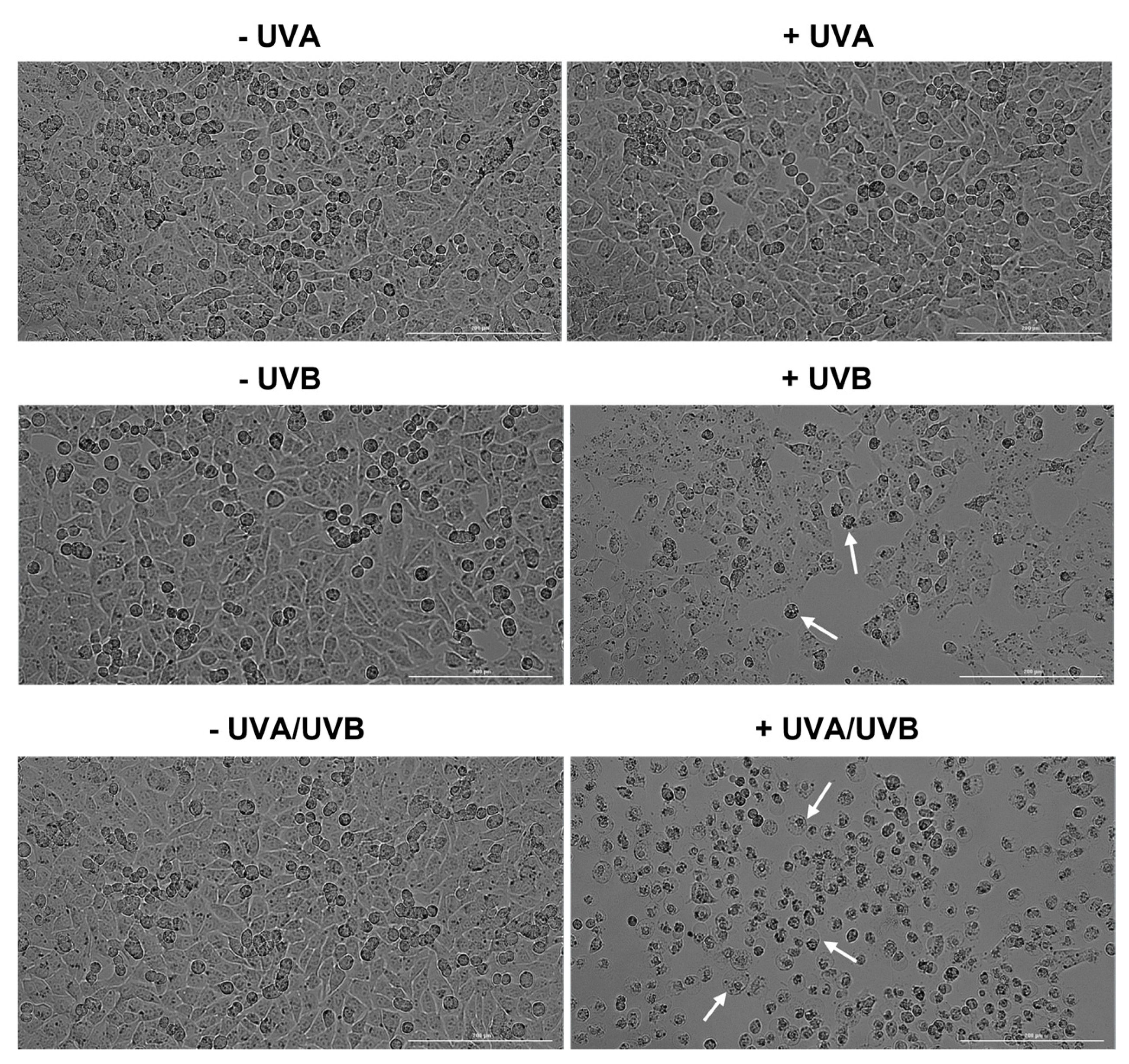
3.2. Evaluation of Cell Morphology and Confluence
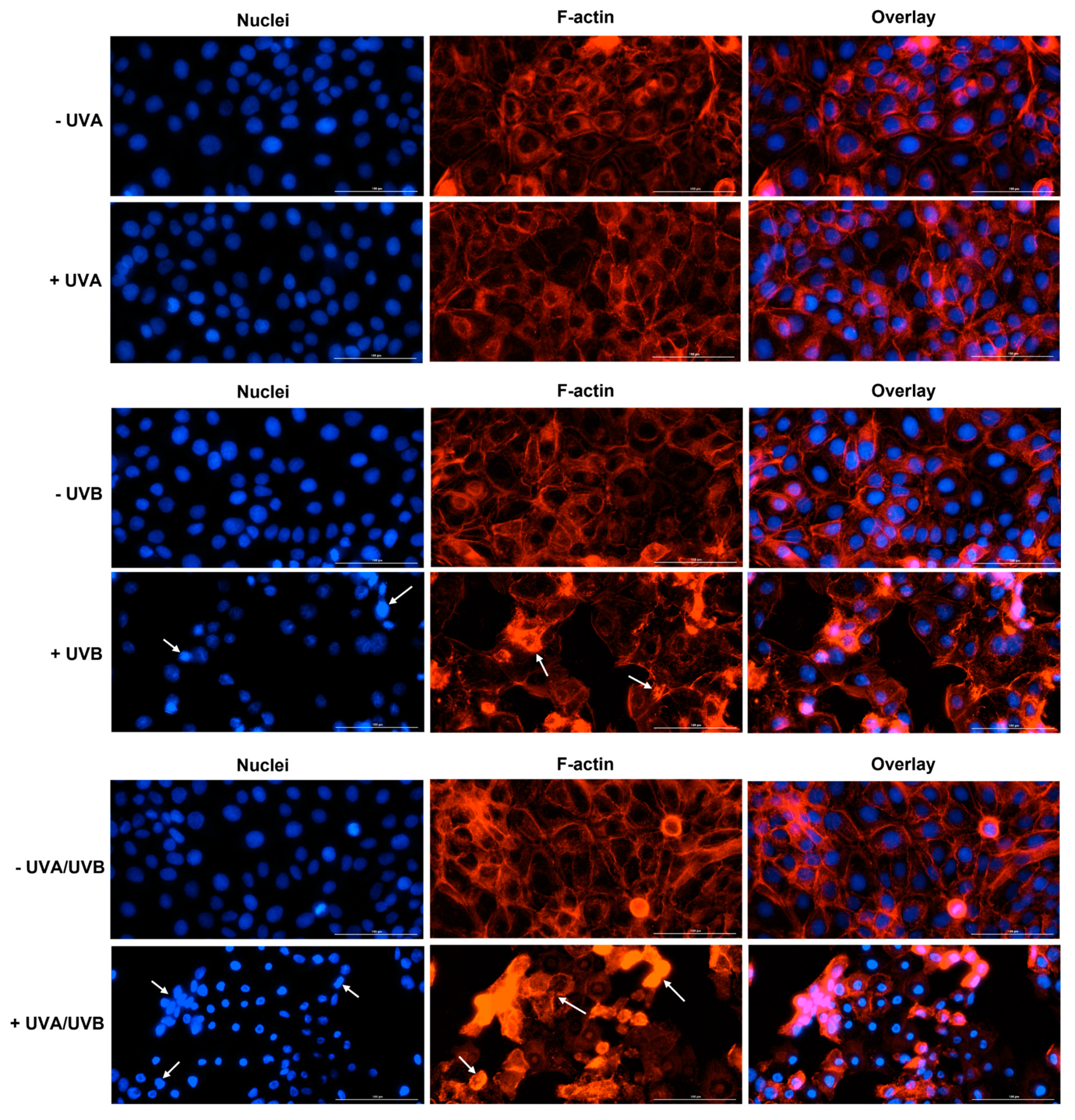
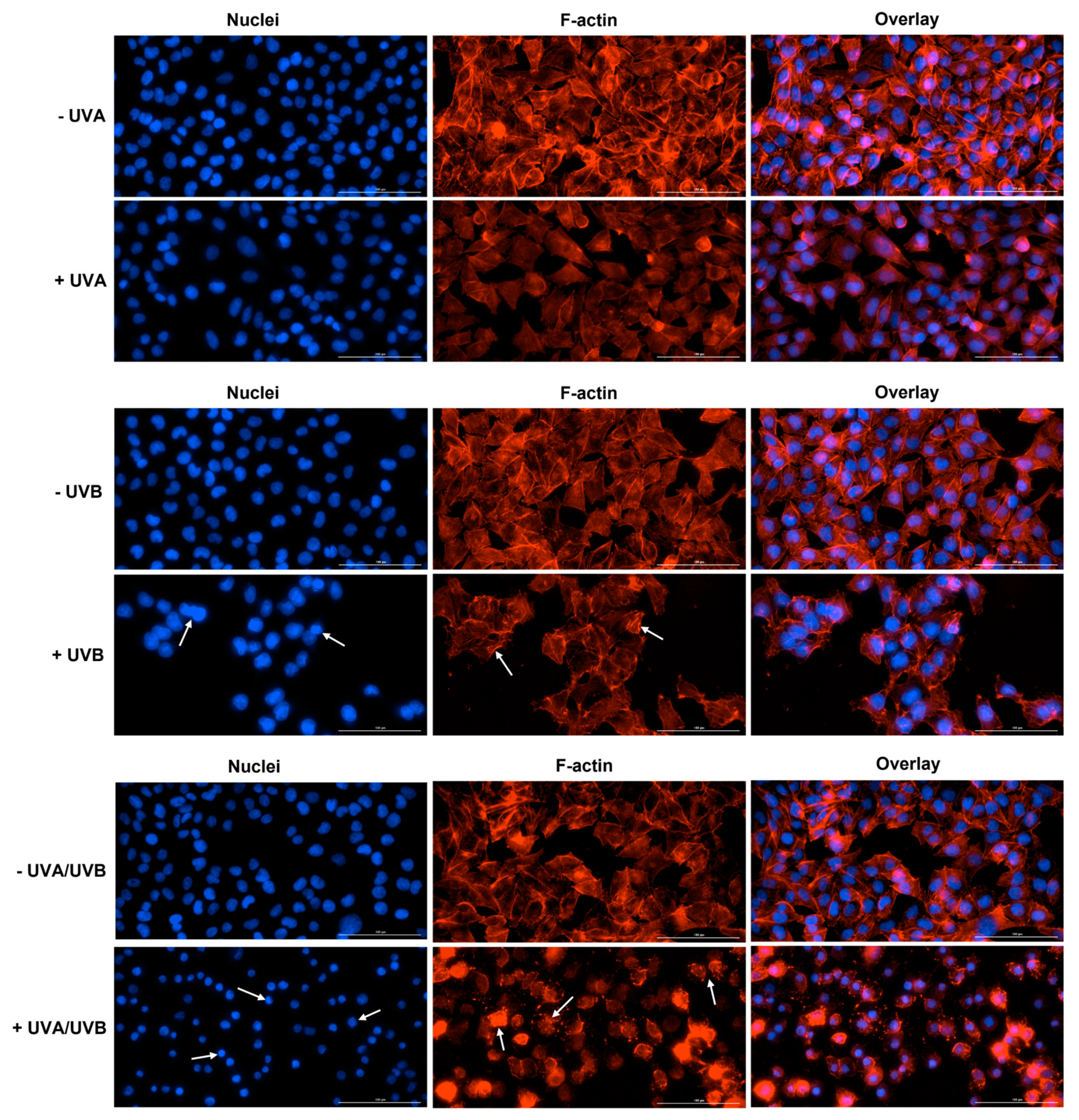
3.3. Immunofluorescence Staining of Cell Nuclei and F-Actin
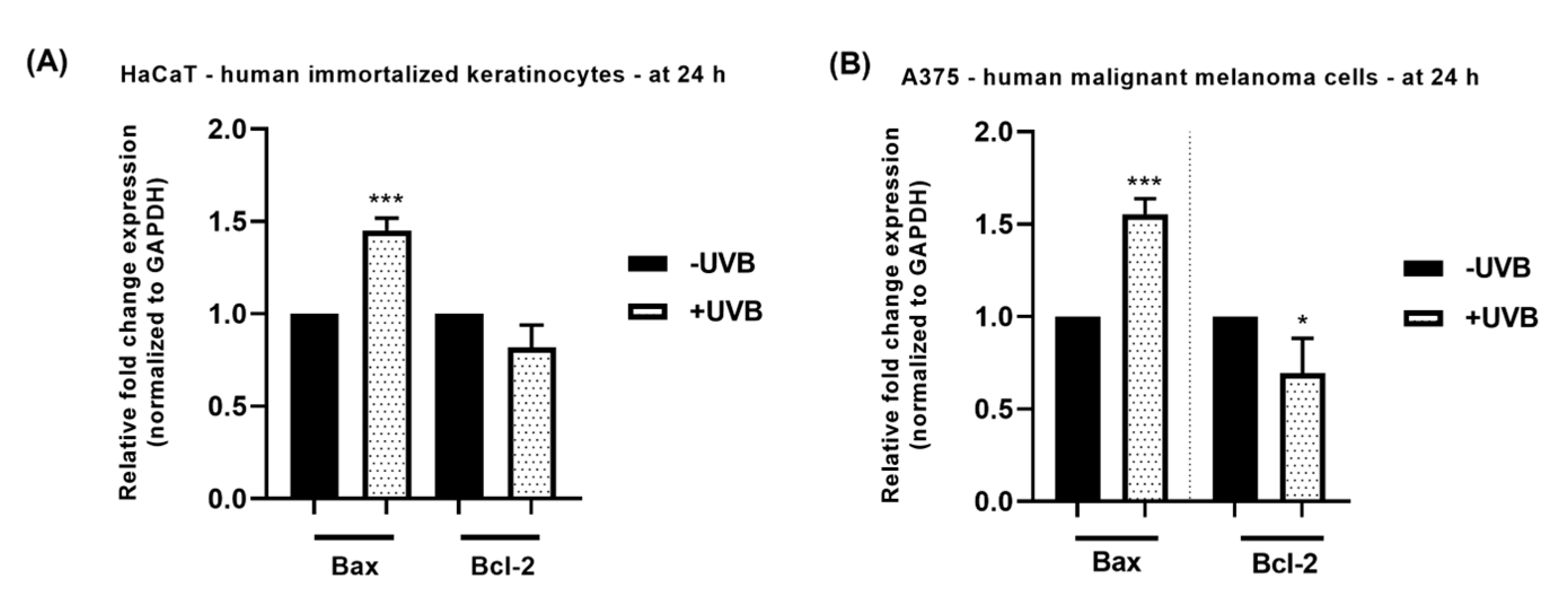
3.4. RT-qPCR Analysis
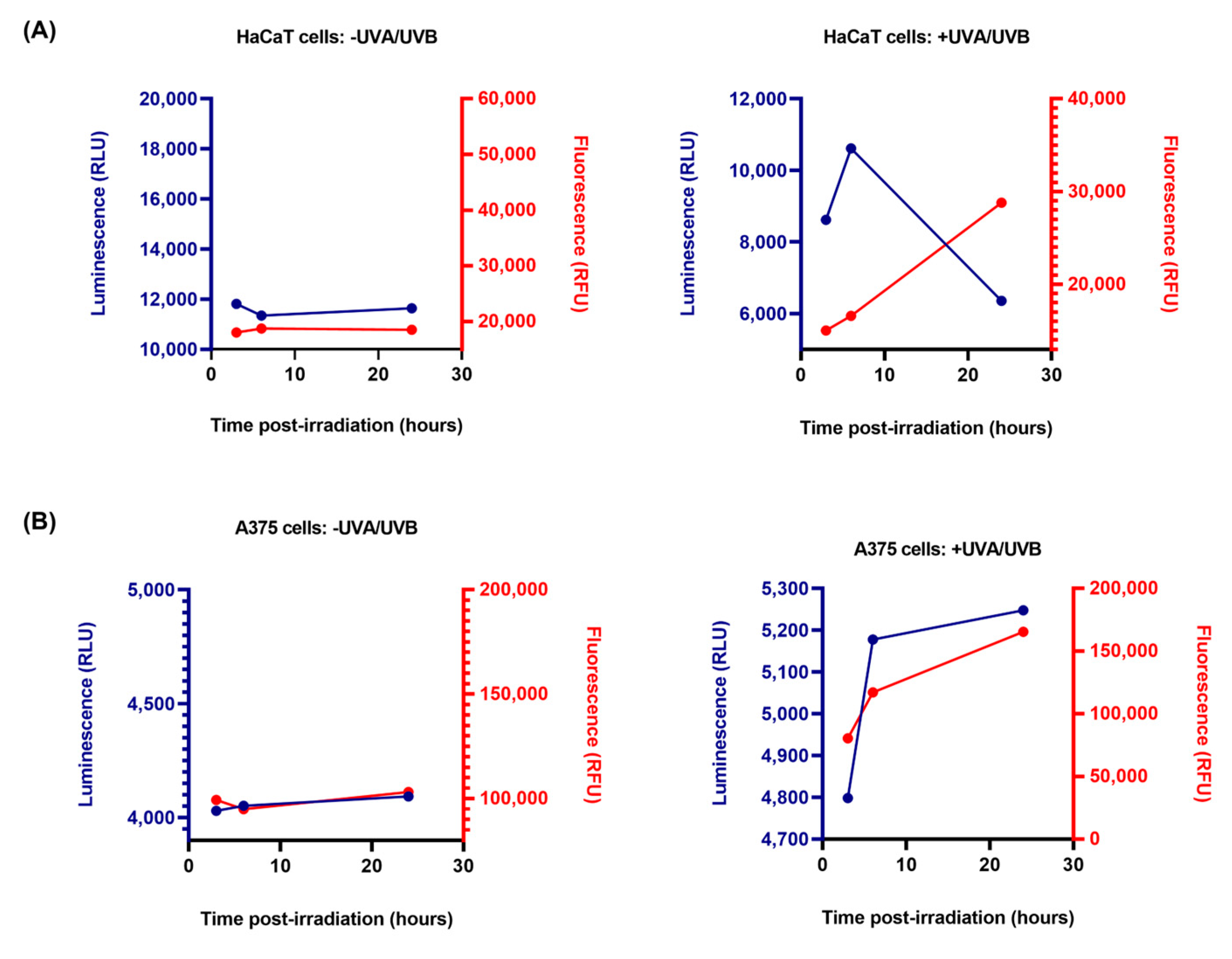
3.5. RealTime-Glo™ Annexin V Apoptosis and Necrosis Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marionnet, C.; Tricaud, C.; Bernerd, F. Exposure to non-extreme solar UV daylight: Spectral characterization, effects on skin and photoprotection. Int. J. Mol. Sci. 2015, 16, 68–90. [Google Scholar] [CrossRef] [PubMed]
- Amaro-Ortiz, A.; Yan, B.; D’Orazio, J.A. Ultraviolet radiation, aging and the skin: Prevention of damage by topical cAMP manipulation. Molecules. 2014, 19, 6202–6219. [Google Scholar] [CrossRef] [PubMed]
- Misovic, M.; Milenkovic, D.; Martinovic, T.; Ciric, D.; Bumbasirevic, V.; Kravic-Stevovic, T. Short-term exposure to UV-A, UV-B, and UV-C irradiation induces alteration in cytoskeleton and autophagy in human keratinocytes. Ultrastruct. Pathol. 2013, 37, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Sample, A.; He, Y.Y. Mechanisms and prevention of UV-induced melanoma. Photodermatol. Photoimmunol. Photomed. 2018, 34, 13–24. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Teresa, M.; Petersen, S.; Prakash, G. UV Light Effects on Proteins: From Photochemistry to Nanomedicine. Mol. Photochem.-Var. Asp. 2012, 125–158. [Google Scholar] [CrossRef]
- Juzeniene, A.; Moan, J. Beneficial effects of UV radiation other than via vitamin D production. Dermatoendocrinology 2012, 4, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.; George, J.; Aucoin, D.; Bower, J.; Burrell, S.; Gilbert, R.; Bower, N. The Pathogenesis and Clinical Management of Cutaneous Melanoma: An Evidence-Based Review. J. Med. Imaging Radiat. Sci. 2019, 50, 460–469. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, N.; Yin, C.; Zhu, B.; Li, X. Ultraviolet Radiation and Melanomagenesis: From Mechanism to Immunotherapy. Front. Oncol. 2020, 10, 951. [Google Scholar] [CrossRef]
- Jenkins, R.W.; Fisher, D.E. Treatment of Advanced Melanoma in 2020 and Beyond. J. Investig. Dermatol. 2021, 141, 23–31. [Google Scholar] [CrossRef]
- Moran, B.; Silva, R.; Perry, A.S.; Gallagher, W.M. Epigenetics of malignant melanoma. Semin. Cancer Biol. 2018, 51, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Mohania, D.; Chandel, S.; Kumar, P.; Verma, V.; Digvijay, K.; Tripathi, D.; Choudhury, K.; Mitten, S.K.; Shah, D. Ultraviolet radiations: Skin defense-damage mechanism. Adv. Exp. Med. Biol. 2017, 996, 71–87. [Google Scholar] [PubMed]
- Rünger, T.M. Mechanisms of Melanoma Promotion by Ultraviolet Radiation. J. Investig. Dermatol. 2016, 136, 1751–1752. [Google Scholar] [CrossRef]
- Craig, S.; Earnshaw, C.H.; Virós, A. Ultraviolet light and melanoma. J. Pathol. 2018, 244, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Liu-Smith, F.; Jia, J.; Zheng, Y. UV-induced molecular signaling differences in melanoma and non-melanoma skin cancer. Adv. Exp. Med. Biol. 2017, 996, 27–40. [Google Scholar]
- Walczak, K.; Kazimierczak, P.; Szalast, K.; Plech, T. Uvb radiation and selected tryptophan-derived ahr ligands—Potential biological interactions in melanoma cells. Int. J. Mol. Sci. 2021, 22, 7500. [Google Scholar] [CrossRef]
- Souza, C.; Mônico, D.A.; Tedesco, A.C. Implications of dichlorofluorescein photoinstability for detection of UVA-induced oxidative stress in fibroblasts and keratinocyte cells. Photochem. Photobiol. Sci. 2020, 19, 40–48. [Google Scholar] [CrossRef]
- Coricovac, D.; Farcas, C.; Nica, C.; Pinzaru, I.; Simu, S.; Stoian, D.; Soica, C.; Proks, M.; Avram, S.; Navolan, D.; et al. Ethinylestradiol and levonorgestrel as active agents in normal skin, and pathological conditions induced by UVB exposure: In vitro and in ovo assessments. Int. J. Mol. Sci. 2018, 19, 3600. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable in Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Tang, Z.; Tong, X.; Huang, J.; Liu, L.; Wang, D.; Yang, S. Research progress of keratinocyte-programmed cell death in UV-induced Skin photodamage. Photodermatol. Photoimmunol. Photomed. 2021, 37, 442–448. [Google Scholar] [CrossRef]
- Rajnochová Svobodová, A.; Ulrichová, J.; Vostálová, J. Human keratinocyte cell line as a suitable alternative model for in vitro phototoxicity testing. An. Bras. Dermatol. 2019, 94, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Avram, S.; Coricovac, D.E.; Pavel, I.Z.; Pinzaru, I.; Ghiulai, R.; Baderca, F.; Soica, C.; Muntean, D.; Branisteanu, D.E.; Spandidos, D.A.; et al. Standardization of A375 human melanoma models on chicken embryo chorioallantoic membrane and Balb/c nude mice. Oncol. Rep. 2017, 38, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Hung, C.T.; Keller, J.J.; Lin, H.C.; Wu, Y.J. Proteomic analysis of retinal pigment epithelium cells after exposure to UVA radiation. BMC Ophthalmol. 2019, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Murray, H.C.; Maltby, V.E.; Smith, D.W.; Bowden, N.A. Nucleotide excision repair deficiency in melanoma in response to UVA. Exp. Hematol. Oncol. 2016, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Ouhtit, A. Deregulation of cell growth and apoptosis in UV-induced melanomagenesis. Front. Biosci.-Elite 2020, 12, 223–236. [Google Scholar] [CrossRef]
- Pan, T.; Zhu, J.; Hwu, W.J.; Jankovic, J. The Role of Alpha-Synuclein in Melanin Synthesis in Melanoma and Dopaminergic Neuronal Cells. PLoS ONE 2012, 7, 3–10. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, H.; Yang, H.; Liang, J.; Li, X. Low-dose UVA radiation-induced adaptive response in cultured human dermal fibroblasts. Int. J. Photoenergy 2012, 2012, 167425. [Google Scholar] [CrossRef]
- Cha, H.J.; Kim, O.Y.; Lee, G.T.; Lee, K.S.; Lee, J.H.; Park, I.C.; Lee, S.J.; Kim, Y.R.; Ahn, K.J.; An, I.S.; et al. Identification of ultraviolet B radiation-induced microRNAs in normal human dermal papilla cells. Mol. Med. Rep. 2014, 10, 1663–1670. [Google Scholar] [CrossRef]
- Khalil, C.; Shebaby, W. UVB damage onset and progression 24 h post exposure in human-derived skin cells. Toxicol. Rep. 2017, 4, 441–449. [Google Scholar] [CrossRef]
- Khalil, C. In Vitro UVB induced Cellular Damage Assessment Using Primary Human Skin Derived Fibroblasts. MOJ Toxicol. 2015, 1, 138–143. [Google Scholar] [CrossRef]
- Wang, P.W.; Hung, Y.C.; Lin, T.Y.; Fang, J.Y.; Yang, P.M.; Chen, M.H.; Pan, T.L. Comparison of the biological impact of UVA and UVB upon the skin with functional proteomics and immunohistochemistry. Antioxidants 2019, 8, 569. [Google Scholar] [CrossRef] [PubMed]
- Bajgar, R.; Moukova, A.; Chalupnikova, N.; Kolarova, H. Differences in the effects of broad-band UVA and narrow-band UVB on epidermal keratinocytes. Int. J. Environ. Res. Public Health 2021, 18, 12480. [Google Scholar] [CrossRef] [PubMed]
- Santacruz-Perez, C.; Tonolli, P.N.; Ravagnani, F.G.; Baptista, M.S. Photochemistry of Lipofuscin and the Interplay of UVA and Visible Light in Skin Photosensitivity. In Photochemistry and Photophysics—Fundamentals to Applications; IntechOpen: London, UK, 2018. [Google Scholar]
- Nakyai, W.; Saraphanchotiwitthaya, A.; Viennet, C.; Humbert, P.; Viyoch, J. An In Vitro Model for Fibroblast Photoaging Comparing Single and Repeated UVA Irradiations. Photochem. Photobiol. 2017, 93, 1462–1471. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-Q.; Wu, J.-F.; Xiao, X.-Q.; Xiao, Q.; Wang, J.; Zuo, F.-G. Narrow-band UVB radiation promotes dendrite formation by activating Rac1 in B16 melanoma cells. Mol. Clin. Oncol. 2013, 1, 858–862. [Google Scholar] [CrossRef]
- Ibuki, Y.; Komaki, Y.; Yang, G.; Toyooka, T. Long-wavelength UVA enhances UVB-induced cell death in cultured keratinocytes: DSB formation and suppressed survival pathway. Photochem. Photobiol. Sci. 2021, 20, 639–652. [Google Scholar] [CrossRef]
- Ziegler, U.; Groscurth, P. Morphological features of cell death. News Physiol. Sci. 2004, 19, 124–128. [Google Scholar] [CrossRef]
- Voss, A.K.; Strasser, A. The essentials of developmental apoptosis. F1000Research 2020, 9, 1–12. [Google Scholar] [CrossRef]
- Lu, S.J.; Feng, Q.; Park, J.S.; Vida, L.; Lee, B.S.; Strausbauch, M.; Wettstein, P.J.; Honig, G.R.; Lanza, R. Biologic properties and enucleation of red blood cells from human embryonic stem cells. Blood 2008, 112, 4475–4484. [Google Scholar] [CrossRef]
- Moras, M.; Lefevre, S.D.; Ostuni, M.A. From erythroblasts to mature red blood cells: Organelle clearance in mammals. Front. Physiol. 2017, 8, 01076. [Google Scholar] [CrossRef]
- Westman, J.; Grinstein, S.; Marques, P.E. Phagocytosis of Necrotic Debris at Sites of Injury and Inflammation. Front. Immunol. 2020, 10, 3030. [Google Scholar] [CrossRef]
- Paunescu, V.; Bojin, F.M.; Gavriliuc, O.I.; Taculescu, E.A.; Ianos, R.; Ordodi, V.L.; Iman, V.F.; Tatu, C.A. Enucleation: A possible mechanism of cancer cell death. J. Cell. Mol. Med. 2014, 18, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Desouza, M.; Gunning, P.W.; Stehn, J.R. The actin cytoskeleton as a sensor and mediator of apoptosis. Bioarchitecture. 2012, 2, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Povea-Cabello, S.; Oropesa-Ávila, M.; de la Cruz-Ojeda, P.; Villanueva-Paz, M.; De La Mata, M.; Suárez-Rivero, J.M.; Álvarez-Córdoba, M.; Villalón-García, I.; Cotán, D.; Ybot-González, P.; et al. Dynamic reorganization of the cytoskeleton during apoptosis: The two coffins hypothesis. Int. J. Mol. Sci. 2017, 18, 2393. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, Y.T.; Cardella, G.D.; Forti, F.L. Exoenzyme C3 transferase lowers actin cytoskeleton dynamics, genomic stability and survival of malignant melanoma cells under UV-light stress. J. Photochem. Photobiol. B Biol. 2020, 209, 111947. [Google Scholar] [CrossRef] [PubMed]
- Nawrot-Hadzik, I.; Choromańska, A.; Abel, R.; Preissner, R.; Saczko, J.; Matkowski, A.; Hadzik, J. Cytotoxic effect of vanicosides a and b from reynoutria sachalinensis against melanotic and amelanotic melanoma cell lines and in silico evaluation for inhibition of brafv600e and mek1. Int. J. Mol. Sci. 2020, 21, 4611. [Google Scholar] [CrossRef] [PubMed]
- Kupcho, K.; Niles, A.; Shultz, J.; Grailer, J.; Zhou, W.; Hurst, R.; Hartnett, J.; Riss, T.; Lazar, D.; Cali, J. Abstract 4296: A real-time annexin V method for monitoring programmed cell death. Cancer Res. 2017, 77, 4296. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward | Reverse |
|---|---|---|
| GAPDH * | 5′ AAG-GTG-AAG-GTC-GGA-GTC-AAC 3′ | 5′ GGG-GTC-ATT-GAT-GGC-AAC-AAT-A 3′ |
| Bax | 5′ GCCGGGTTGTCGCCCTTTT 3′ | 5′ CCGCTCCCGGAGGAAGTCCA 3′ |
| Bcl-2 | 5′ CGGGAGATGTCGCCCCTGGT 3′ | 5′ GCATGCTGGGGCCGTACAGT 3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gag, O.; Dinu, Ș.; Manea, H.; Marcovici, I.; Pînzaru, I.; Popovici, R.; Crăiniceanu, Z.; Gyori, Z.; Iovănescu, G.; Chiriac, S. UVA/UVB Irradiation Exerts a Distinct Phototoxic Effect on Human Keratinocytes Compared to Human Malignant Melanoma Cells. Life 2023, 13, 1144. https://doi.org/10.3390/life13051144
Gag O, Dinu Ș, Manea H, Marcovici I, Pînzaru I, Popovici R, Crăiniceanu Z, Gyori Z, Iovănescu G, Chiriac S. UVA/UVB Irradiation Exerts a Distinct Phototoxic Effect on Human Keratinocytes Compared to Human Malignant Melanoma Cells. Life. 2023; 13(5):1144. https://doi.org/10.3390/life13051144
Chicago/Turabian StyleGag, Otilia, Ștefania Dinu, Horațiu Manea, Iasmina Marcovici, Iulia Pînzaru, Ramona Popovici, Zorin Crăiniceanu, Zsolt Gyori, Gheorghe Iovănescu, and Sorin Chiriac. 2023. "UVA/UVB Irradiation Exerts a Distinct Phototoxic Effect on Human Keratinocytes Compared to Human Malignant Melanoma Cells" Life 13, no. 5: 1144. https://doi.org/10.3390/life13051144