

Beneficial Microorganisms Improve Agricultural Sustainability under Climatic Extremes
 , ,
, ,  and
and 
Abstract
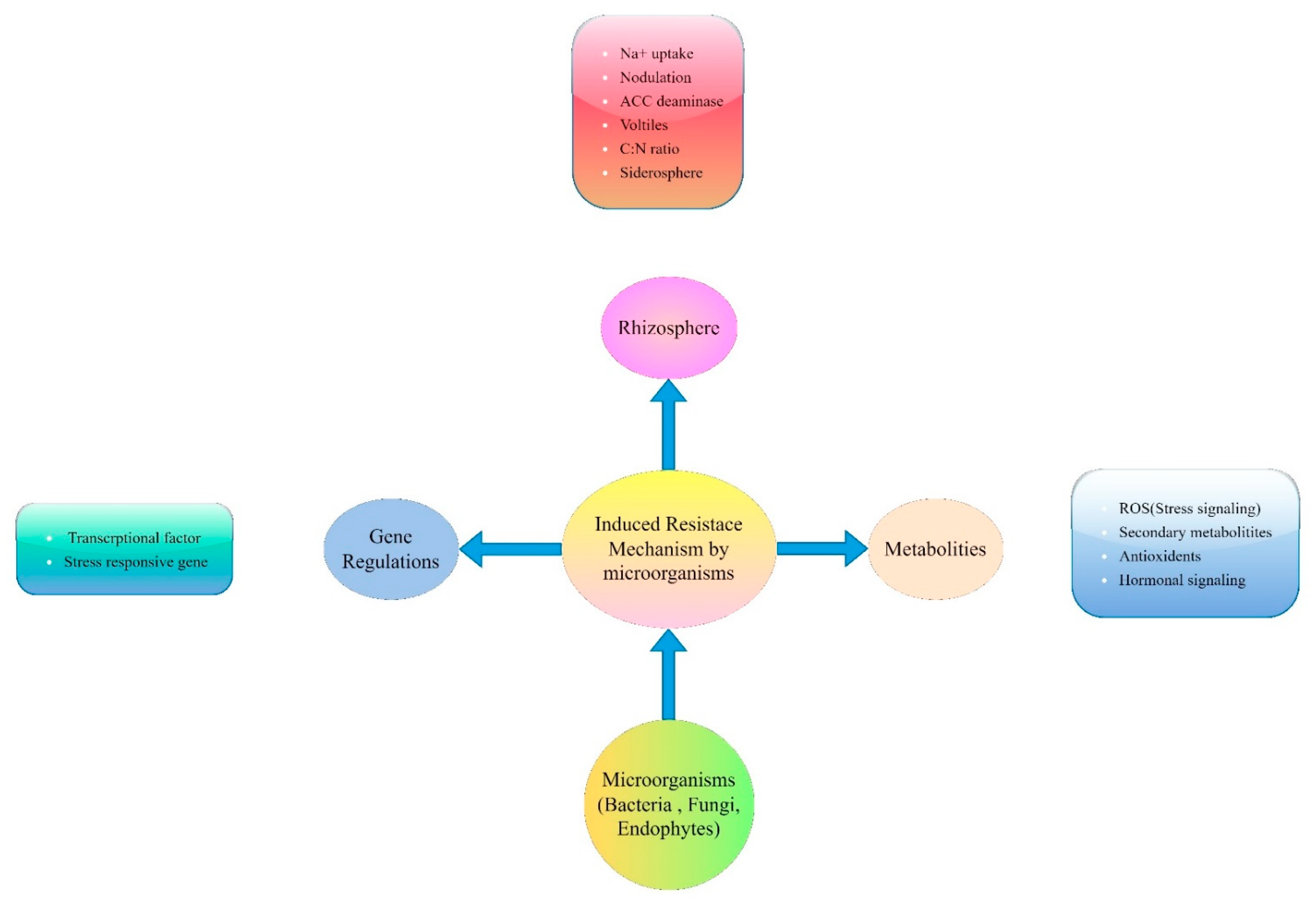
:1. Introduction
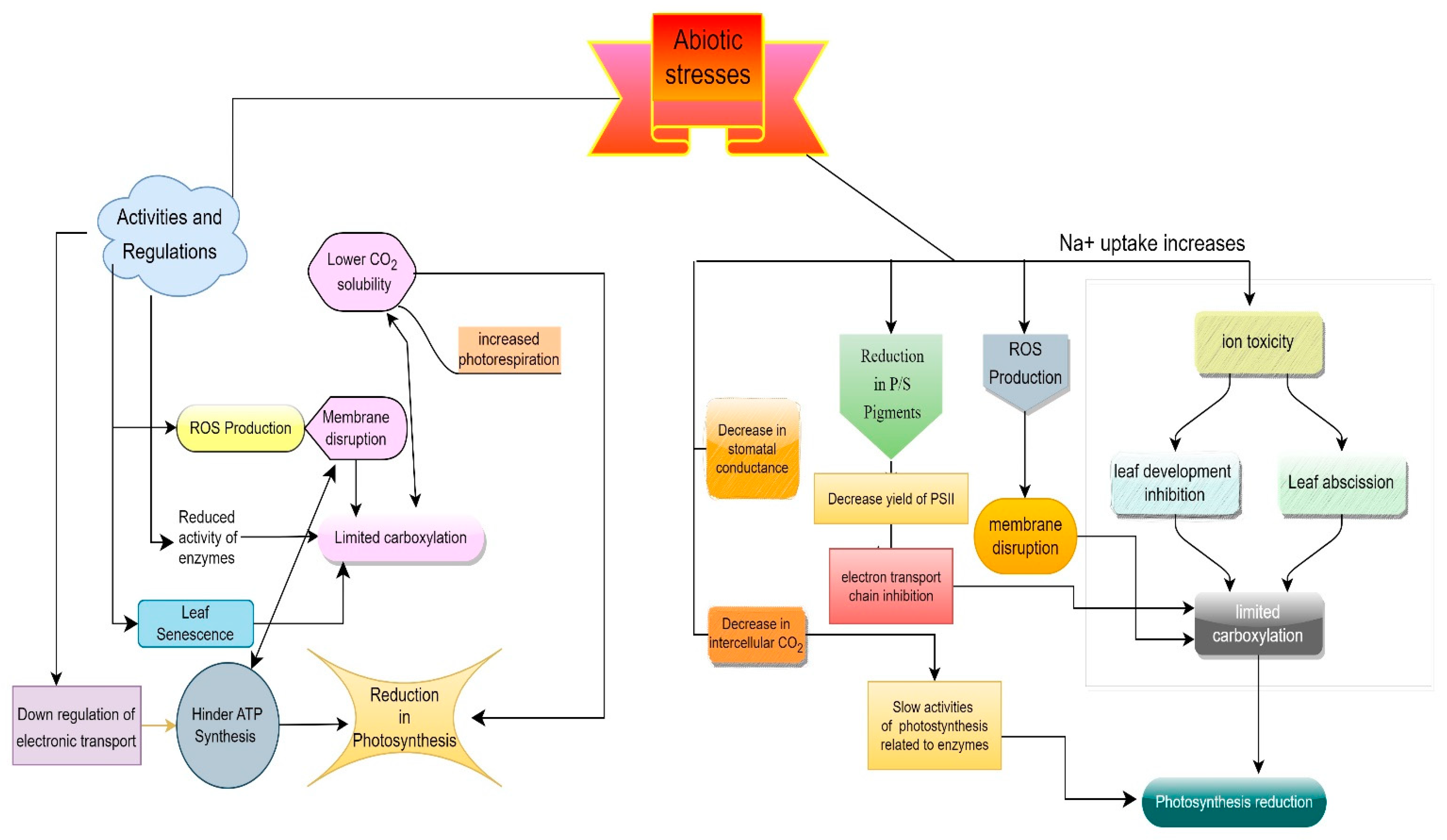
2. Drought Stress
3. Salt Stress
4. Heavy Metals
5. High Temperature
6. Low Temperature
7. Flood Stress and Oxygen Deficit
8. Light Stress
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, S.; Zuo, T.; Ni, W. Important roles of glycinebetaine in stabilizing the structure and function of the photosystem II complex under abiotic stresses. Planta 2020, 251, 36. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; Balestrini, R.; Chitarra, W.; Marsico, A.D.; Nerva, L. Getting ready with the priming: Innovative weapons against biotic and abiotic crop enemies in a global changing scenario. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Academic Press: Cambridge, MA, USA, 2020; pp. 35–56. [Google Scholar] [CrossRef]
- Ali, S.; Eum, H.; Cho, J.; Dan, L.; Khan, F.; Dairaku, K.; Shrestha, M.L.; Hwang, S.; Nasim, W.; Khan, I.A.; et al. Assessment of climate extremes in future projections downscaled by multiple statistical downscaling methods over Pakistan. Atmos. Res. 2019, 222, 114–133. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping plant responses to biotic stress by chlorophyll fluorescence imaging. Front. Plant Sci. 2019, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- Pawłowicz, I.; Masajada, K. Aquaporins as a link between water relations and photosynthetic pathway in abiotic stress tolerance in plants. Gene 2019, 687, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Oliveira, C.E.; Zoz, T.; Jalal, A.; Vendruscolo, E.P.; Nogueira, T.A.R.; Jani, A.D.; Teixeira Filho, M.C.M. Tolerance and Adaptability of Tomato Genotypes to Saline Irrigation. Crops 2022, 2, 306–322. [Google Scholar] [CrossRef]
- Tunçtürk, M.; Rezaee Danesh, Y.; Tunçtürk, R.; Oral, E.; Najafi, S.; Nohutçu, L.; Jalal, A.; da Silva Oliveira, C.E.; Filho, M.C.M.T. Safflower (Carthamus tinctorius L.) response to cadmium stress: Morpho-physiological traits and mineral concentrations. Life 2023, 13, 135. [Google Scholar] [CrossRef]
- Ullah, R.; Muhammad, S. Heavy metals contamination in soils and plants along with the mafic-ultramafic complex (Ophiolites), Baluchistan, Pakistan: Evaluation for risk and phytoremediation potential. Environ. Technol. Innov. 2020, 19, 100931. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, F.; Meng, Y.; Chandrasekaran, U.; Luo, X.; Yang, W.; Shu, K. Plant waterlogging/flooding stress responses: From seed germination to maturation. Plant Physiol. Biochem. 2020, 148, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, L.R.; Souza, V.T.; Campos, R.A.; Rüther, R. Extreme solar overirradiance events: Occurrence and impacts on utility-scale photovoltaic power plants in Brazil. Solar Energy 2019, 186, 370–381. [Google Scholar] [CrossRef]
- Gobbett, D.L.; Nidumolu, U.; Crimp, S. Modelling frost generates insights for managing risk of minimum temperature extremes. Weather Clim. Extrem. 2020, 27, 100176. [Google Scholar] [CrossRef]
- Tavakol, A.; Rahmani, V.; Harrington Junior, J. Evaluation of hot temperature extremes and heat waves in the Mississippi River Basin. Atmos. Res. 2020, 239, 104907. [Google Scholar] [CrossRef]
- Balestrini, R.; Chitarra, W.; Fotopoulos, V.; Ruocco, M. Potential role of beneficial soil microorganisms in plant tolerance to abiotic stress factors. In Soil Biological Communities and Ecosystem Resilience; Springer: Cham, Switzerland, 2017; pp. 191–207. [Google Scholar]
- Ma, Y.; Vosátka, M.; Freitas, H. Editorial: Beneficial Microbes Alleviate Climatic Stresses in Plants. Front. Plant Sci. 2019, 10, 595. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Swain, C.K. Role of Microbes in Climate Smart Agriculture. In Microorganisms for Green Revolution. Microorganisms for Sustainability; Panpatte, D., Jhala, Y., Shelat, H., Vyas, R., Eds.; Springer: Singapore, 2018; p. 7. [Google Scholar] [CrossRef]
- Saghafi, D.; Delangiz, N.; Lajayer, B.A.; Ghorbanpour, M. An overview on improvement of crop productivity in saline soils by halotolerant and halophilic PGPRs. J. Biotechnol. 2019, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef]
- Arif, I.; Batool, M.; Schenk, P.M. Plant Microbiome Engineering: Expected Benefits for Improved Crop Growth and Resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef]
- Dogra, N.; Yadav, R.; Kaur, M.; Adhikary, A.; Kumar, S.; Ramakrishna, W. Nutrient enhancement of chickpea grown with plant growth promoting bacteria in local soil of Bathinda, Northwestern India. Physiol. Mol. Biol. Plants 2019, 25, 1251–1259. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Jalal, A.; da Silva Oliveira, C.E.; Galindo, F.S.; Rosa, P.A.L.; Gato, I.M.B.; de Lima, B.H.; Teixeira Filho, M.C.M. Regulatory Mechanisms of Plant Growth-Promoting Rhizobacteria and Plant Nutrition against Abiotic Stresses in Brassicaceae Family. Life 2023, 13, 211. [Google Scholar] [CrossRef]
- Greetatorn, T.; Hashimoto, S.; Sarapat, S.; Tittabutr, P.; Boonkerd, N.; Uchiumi, T.; Teaumroong, N. Empowering rice seedling growth by endophytic Bradyrhizobium sp. SUTN 9-2. Lett. Appl. Microbiol. 2019, 68, 258–266. [Google Scholar] [CrossRef]
- Da Silveira, A.P.D.; Iório, R.D.P.F.; Marcos, F.C.C.; Fernandes, A.O.; de Souza, S.A.C.D.; Kuramae, E.E.; Cipriano, M.A.P. Exploitation of new endophytic bacteria and their ability to promote sugarcane growth and nitrogen nutrition. Antonie Van Leeuwenhoek 2019, 112, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, P.; Kashyap, P.L.; Kuppusamy, P.; Srivastava, A.K.; Tiwari, R.K. Functional characterization of endophytic bacilli from pearl millet (Pennisetum glaucum) and their possible role in multiple stress tolerance. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2019, 154, 503–514. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Ullah, I.; Ali, S.; Kang, S.M.; Lee, I.J. Alleviation of salt stress response in soybean plants with the endophytic bacterial isolate Curtobacterium sp. SAK1. Ann. Microbiol. 2019, 69, 797–808. [Google Scholar] [CrossRef]
- Konkolewska, A.; Piechalak, A.; Ciszewska, L.; Antos-Krzemińska, N.; Skrzypczak, T.; Hanć, A.; Sitko, K.; Małkowski, E.; Barałkiewicz, D.; Małecka, A. Combined use of companion planting and PGPR for the assisted phytoextraction of trace metals (Zn, Pb, Cd). Environ. Sci. Pollut. Res. 2020, 27, 13809–13825. [Google Scholar] [CrossRef] [PubMed]
- Costa-Gutierrez, S.B.; Lami, M.J.; Santo, M.C.C.D.; Zenoff, A.M.; Vincent, P.A.; Molina-Henares, M.A.; Espinosa-Urgel, M.; Cristóbal, R.E. Plant growth promotion by Pseudomonas putida KT2440 under saline stress: Role of eptA. Appl. Microbiol. Biotechnol. 2020, 104, 4577–4592. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Kumar, A.; Maurya, B.R.; Raghuwanshi, R.; Meena, V.S.; Islam, M.T. Co-inoculation with Enterobacter and Rhizobacteria on yield and nutrient uptake by wheat (Triticum aestivum L.) in the alluvial soil under indo-gangetic plain of India. J. Plant Growth Regul. 2017, 36, 608–617. [Google Scholar] [CrossRef]
- Bokhari, A.; Essack, M.; Lafi, F.F.; Barrao, C.A.; Jalal, R.; Alamoudi, S.; Razali, R.; Alzubaidy, H.; Shah, K.H.; Siddique, S.; et al. Bioprospecting desert plant Bacillus endophytic strains for their potential to enhance plant stress tolerance. Sci. Rep. 2019, 9, 18154. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A. Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environ. Exp. Bot. 2019, 159, 55–65. [Google Scholar] [CrossRef]
- Alen’kina, A.S.; Nikitina, V.E. Effect of Azospirillum Lectins on the Ascorbate Peroxidase Activity and Ascorbic Acid Content in Wheat Seedling Roots Exposed to Abiotic Stresses. Appl. Biochem. Microbiol. 2020, 56, 211–218. [Google Scholar] [CrossRef]
- Leite, R.D.C.; dos Santos, J.G.; Silva, E.L.; Alves, C.R.; Hungria, M.; Leite, R.D.C.; dos Santos, A.C. Productivity increase, reduction of nitrogen fertiliser use and drought-stress mitigation by inoculation of Marandu grass (Urochloa brizantha) with Azospirillum brasilense. Crop Pasture Sci. 2018, 70, 61–67. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Amelioration of drought tolerance in wheat by the interaction of plant growth-promoting rhizobacteria. Plant Biol. 2016, 18, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.; Tripathi, P.; Chandra, A. Isolation and molecular characterization of plant growth-promoting Bacillus spp. and their impact on sugarcane (Saccharum spp. hybrids) growth and tolerance towards drought stress. Acta Physiol. Plant 2018, 40, 199. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef] [PubMed]
- Jochum, M.; McWilliams, K.M.; Borrego, E.; Kolomiets, M.; Niu, G.; Pierson, E.; Jo, Y.K. Bioprospecting plant growth-promoting rhizobacteria that mitigate drought stress in grasses. Front. Microbiol. 2019, 10, 2106. [Google Scholar] [CrossRef]
- Selim, S.; Hassan, Y.M.; Saleh, A.M.; Habeeb, T.H.; AbdElgawad, H. Actinobacterium isolated from a semi-arid environment improves the drought tolerance in maize (Zea mays L.). Plant Physiol. Biochem. 2019, 142, 15–21. [Google Scholar] [CrossRef]
- Ullah, A.; Akbar, A.; Luo, Q.; Khan, A.H.; Manghwar, H.; Shaban, M.; Yang, X. Microbiome diversity in cotton rhizosphere under normal and drought conditions. Microb. Ecol. 2019, 77, 429–439. [Google Scholar] [CrossRef]
- Silva, E.R.; Zoz, J.; Oliveira, C.E.S.; Zuffo, A.M.; Steiner, F.; Zoz, T.; Vendruscolo, E.P. Can co-inoculation of Bradyrhizobium and Azospirillum alleviate adverse effects of drought stress on soybean (Glycine max L. Merrill.)? Arch. Microbiol. 2019, 201, 325–335. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Sheikh, I.; Kumar, V.; Dhaliwal, H.S.; Saxena, A.K. Amelioration of drought stress in Foxtail millet (Setaria italica L.) by P-solubilizing drought-tolerant microbes with multifarious plant growth promoting attributes. Environ. Sustain. 2020, 3, 23–34. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, L.; Hao, R.; Bai, X.; Wang, Y.; Yu, X. Drought-tolerant plant growth-promoting rhizobacteria isolated from jujube (Ziziphus jujuba) and their potential to enhance drought tolerance. Plant Soil 2020, 452, 423–440. [Google Scholar] [CrossRef]
- Steiner, F.; Oliveira, C.E.S.; Zoz, T.; Zuffo, A.M.; Freitas, R.S. Co-Inoculation of Common Bean with Rhizobium and Azospirillum Enhance the Drought Tolerance. Russ. J. Plant Physiol. 2020, 67, 923–932. [Google Scholar] [CrossRef]
- Viscardi, S.; Ventorino, V.; Duran, P.; Maggio, A.; De Pascale, S.; Mora, M.L.; Pepe, O. Assessment of plant growth promoting activities and abiotic stress tolerance of Azotobacter chroococcum strains for a potential use in sustainable agriculture. J. Soil Sci. Plant Nutr. 2016, 16, 848–863. [Google Scholar] [CrossRef]
- Hahm, M.S.; Son, J.S.; Hwang, Y.J.; Kwon, D.K.; Ghim, S.Y. Alleviation of Salt Stress in Pepper (Capsicum annum L.) Plants by Plant Growth-Promoting Rhizobacteria. J. Microbiol. Biotechnol. 2017, 27, 1790–1797. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, Z.; He, Y.; Huang, Y.; Li, X.; Ye, B.C. Plant growth promotion and alleviation of salinity stress in Capsicum annuum L. by Bacillus isolated from saline soil in Xinjiang. Ecotoxicol. Environ. Saf. 2018, 164, 520–529. [Google Scholar] [CrossRef]
- Ansari, M.; Shekari, F.; Mohammadi, M.H.; Juhos, K.; Végvári, G.; Biró, B. Salt-tolerant plant growth-promoting bacteria enhanced salinity tolerance of salt-tolerant alfalfa (Medicago sativa L.) cultivars at high salinity. Acta Physiol. Plant 2019, 41, 195. [Google Scholar] [CrossRef]
- Bhise, K.K.; Dandge, P.B. Alleviation of salinity stress in rice plant by encapsulated salt tolerant plant growth promoting bacteria Pantoea agglomerans strain KL and its root colonization ability. Arch. Agron. Soil Sci. 2019, 79, 1955–1968. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.M.; Lee, I.J. Halotolerant rhizobacterial strains mitigate the adverse effects of NaCl stress in soybean seedlings. BioMed Res. Int. 2019, 2019, 9530963. [Google Scholar] [CrossRef]
- Yoo, S.J.; Weon, H.Y.; Song, J.; Sang, M.K. Induced tolerance to salinity stress by halotolerant Bacteria Bacillus aryabhattai H19-1 and B. mesonae H20-5 in tomato plants. J. Microbiol. Biotechnol. 2019, 29, 1124–1136. [Google Scholar] [CrossRef]
- Chu, T.N.; Tran, B.T.H.; Van Bui, L.; Hoang, M.T.T. Plant growth-promoting rhizobacterium Pseudomonas PS01 induces salt tolerance in Arabidopsis thaliana. BMC Res. Notes 2019, 12, 11. [Google Scholar] [CrossRef]
- Azadikhah, M.; Jamali, F.; Nooryazdan, H.R.; Bayat, F. Growth promotion and yield enhancement of barley cultivars using ACC deaminase producing Pseudomonas fluorescens strains under salt stress. Span. J. Agric. Res. 2019, 17, e0801. [Google Scholar] [CrossRef]
- Safdarian, M.; Askari, H.; Shariati, J.V.; Nematzadeh, G. Transcriptional responses of wheat roots inoculated with Arthrobacter nitroguajacolicus to salt stress. Sci. Rep. 2019, 9, 1792. [Google Scholar] [CrossRef] [PubMed]
- Hemida, K.A.; Reyad, A.M.M. Improvement salt tolerance of safflower plants by endophytic bacteria. J. Hortic. Plant Res. 2019, 5, 38–56. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Tavasolee, A.; Ghassemi-Golezani, K.; Torabian, S.; Monirifar, H.; Rahmani, H.A. Growth-promoting bacteria and natural regulators mitigate salt toxicity and improve rapeseed plant performance. Protoplasma 2020, 257, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, U.B.; Travedi, M.; Sahu, P.K.; Paul, S.; Paul, D.; Saxena, A.K. Seed biopriming with salt-tolerant endophytic Pseudomonas geniculata-modulated biochemical responses provide ecological fitness in maize (Zea mays L.) grown in saline sodic soil. Int. J. Environ. Res. Public Health 2020, 17, 253. [Google Scholar] [CrossRef]
- El-Akhdar, I.; Elsakhawy, T.; Abo-Koura, H.A. Alleviation of Salt Stress on Wheat (Triticum aestivum L.) by Plant Growth Promoting Bacteria strains Bacillus halotolerans MSR-H4 and Lelliottia amnigena MSR-M49. J. Adv. Microbiol. 2020, 20, 44–58. [Google Scholar] [CrossRef]
- Amiri, R.; Nikbakht, A.; Etemadi, N. Alleviation of drought stress on rose geranium [Pelargonium graveolens (L.) Herit.] in terms of antioxidant activity and secondary metabolites by mycorrhizal inoculation. Sci. Hortic. 2015, 197, 373–380. [Google Scholar] [CrossRef]
- Guler, N.S.; Pehlivan, N.; Karaoglu, S.A.; Guzel, S.; Bozdeveci, A. Trichoderma atroviride ID20G inoculation ameliorates drought stress-induced damages by improving antioxidant defence in maize seedlings. Acta Physiol. Plant. 2016, 38, 132. [Google Scholar] [CrossRef]
- Ban, Y.; Zhouying, X.U.; Yang, Y.; Zhang, H.; Chen, H.; Tang, M. Effect of dark septate endophytic fungus Gaeumannomyces cylindrosporus on plant growth, photosynthesis and Pb tolerance of maize (Zea mays L.). Pedosphere 2017, 27, 283–292. [Google Scholar] [CrossRef]
- Yooyongwech, S.; Samphumphuang, T.; Tisarum, R.; Theerawitaya, C.; Cha-Um, S. Arbuscular mycorrhizal fungi (AMF) improved water deficit tolerance in two different sweet potato genotypes involves osmotic adjustments via soluble sugar and free proline. Sci. Hortic. 2016, 198, 107–117. [Google Scholar] [CrossRef]
- Huang, Y.M.; Zou, Y.N.; Wu, Q.S. Alleviation of drought stress by mycorrhizas is related to increased root H2O2 efflux in trifoliate orange. Sci. Rep. 2017, 7, 42335. [Google Scholar] [CrossRef]
- Mona, S.A.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Soliman, D.W.K.; Wirth, S.; Egamberdieva, D. Increased resistance of drought by Trichoderma harzianum fungal treatment correlates with increased secondary metabolites and proline content. J. Integr. Agric. 2017, 16, 1751–1757. [Google Scholar] [CrossRef]
- Mathur, S.; Tomar, R.S.; Jajoo, A. Arbuscular mycorrhizal fungi (AMF) protects photosynthetic apparatus of wheat under drought stress. Photosynth. Res. 2019, 139, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Meng, B.; Chai, H.; Yang, X.; Song, W.; Li, S.; Lu, A.; Zhang, T.; Sun, W. Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. Front. Plant Sci. 2019, 10, 499. [Google Scholar] [CrossRef] [PubMed]
- Bashyal, B.M.; Zaidi, N.W.; Singh, U.S.; Aggarwal, R. Effect of fungal biocontrol agents on enhancement of drought stress tolerance in rice (Oryza sativa L.). Indian J. Biochem. Biophys. (IJBB) 2020, 57, 101–108. [Google Scholar]
- Hashem, A.; Adb-Allah, E.F.; Alqarawi, A.A.; Aldubise, A.; Egamberdieva, D. Arbuscular mycorrhizal fungi enhances salinity tolerance of Panicum turgidum Forssk by altering photosynthetic and antioxidant pathways. J. Plant Interact. 2015, 10, 230–242. [Google Scholar] [CrossRef]
- Ahmad, P.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; John, R.; Egamberdieva, D.; Gucel, S. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L.) through antioxidative defense system. Front. Plant Sci. 2015, 14, 868. [Google Scholar] [CrossRef]
- Rubio, M.B.; Hermosa, R.; Vicente, R.; Gómez-Acosta, F.A.; Morcuende, R.; Monte, E.; Bettiol, W. The combination of Trichoderma harzianum and chemical fertilization leads to the deregulation of phytohormone networking, preventing the adaptive responses of tomato plants to salt stress. Front. Plant Sci. 2017, 8, 294. [Google Scholar] [CrossRef]
- Yasmeen, R.; Siddiqui, Z.S. Ameliorative effects of Trichoderma harzianum on monocot crops under hydroponic saline environment. Acta Physiol. Plantae 2018, 40, 4. [Google Scholar] [CrossRef]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa). Microbiol. Res. 2018, 206, 25–32. [Google Scholar] [CrossRef]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.B.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd_Allah, E.F. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef]
- Molina-Montenegro, M.A.; Acuña-Rodríguez, I.S.; Torres-Díaz, C.; Gundel, P.R. Root endophytes improve physiological performance and yield in crops under salt stress by up-regulating the foliar sodium concentration. bioRxiv 2018. [Google Scholar] [CrossRef]
- Armada, E.; Probanza, A.; Roldán, A.; Azcón, R. Native plant growth promoting bacteria Bacillus thuringiensis and mixed or individual mycorrhizal species improved drought tolerance and oxidative metabolism in Lavandula dentata plants. J. Plant Physiol. 2016, 192, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, M.; Naghdi, R.; Kartoolinejad, D. Milkweed seedlings tolerance against water stress: Comparison of inoculations with Rhizophagus irregularis and Pseudomonas putida. Environ. Technol. Innov. 2018, 10, 111–121. [Google Scholar] [CrossRef]
- Ghanbarzadeh, Z.; Mohsenzadeh, S.; Rowshan, V.; Zarei, M. Mitigation of water deficit stress in Dracocephalum moldavica by symbiotic association with soil microorganisms. Sci. Hortic. 2020, 272, 109549. [Google Scholar] [CrossRef]
- Aalipour, H.; Nikbakht, A.; Etemadi, N.; Rejali, F.; Soleimani, M. Biochemical response and interactions between arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria during establishment and stimulating growth of Arizona cypress (Cupressus arizonica G.) under drought stress. Sci. Hortic. 2020, 261, 108923. [Google Scholar] [CrossRef]
- Younesi, O.; Moradi, A. Effects of plant growth-promoting rhizobacterium (pgpr) and arbuscular mycorrhizal fungus (AMF) on antioxidant enzyme activities in salt-stressed bean (Phaseolus vulgaris L.). Agriculture (Poľnohospodárstvo) 2014, 60, 10–21. [Google Scholar] [CrossRef]
- Lee, Y.; Krishnamoorthy, R.; Selvakumar, G.; Kim, K.; Sa, T. Alleviation of salt stress in maize plant by co-inoculation of arbuscular mycorrhizal fungi and Methylobacterium oryzae CBMB20. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 33–540. [Google Scholar] [CrossRef]
- Hashem, A.; Adb-Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Shah, M.A. Induction of Osmoregulation and Modulation of Salt Stress in Acacia gerrardii Benth by Arbuscular Mycorrhizal Fungi and Bacillus subtilis (BERA 71). Biomed. Res. Int. 2016, 2016, 6294098. [Google Scholar] [CrossRef]
- Silva, L.V.D.; Oliveira, S.B.R.D.; Azevedo, L.A.D.; Rodrigues, A.C.; Bonifacio, A. Coinoculation with Bradyrhizobium and Trichoderma alleviates the effects of salt stress in cowpea. Rev. Caatinga 2019, 32, 336–344. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Asaf, S.; Lee, I.J. What is there in seeds? Vertically transmitted endophytic resources for sustainable improvement in plant growth. Front. Plant Sci. 2018, 9, 24. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Imran, M.; Jan, R.; Kim, K.M.; Lee, I.J. Synergistic association of endophytic fungi enhances Glycine max L. resilience to combined abiotic stresses: Heavy metals, high temperature and drought stress. Ind. Crops Prod. 2020, 143, 111931. [Google Scholar] [CrossRef]
- Bilal, S.; Ali, L.; Khan, A.L.; Shahzad, R.; Asaf, S.; Imran, M.; Kang, S.M.; Kim, S.K.; Lee, I.J. Endophytic fungus Paecilomyces formosus LHL10 produces sester-terpenoid YW3548 and cyclic peptide that inhibit urease and α-glucosidase enzyme activities. Arch. Microbiol. 2018, 200, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, M.R.; Mirzaei, M.; Ghabooli, M.; Khatabi, B.; Wu, Y.; Zabet-Moghaddam, M.; Mohammadi-Nejad, G.; Haynes, P.A.; Hajirezaei, M.R.; Sepehri, M.; et al. Root endophytic fungus Piriformospora indica improves drought stress adaptation in barley by metabolic and proteomic reprogramming. Environ. Exp. Bot. 2019, 157, 197–210. [Google Scholar] [CrossRef]
- Jang, S.; Jang, Y.; Kim, C.W.; Lee, H.; Hong, J.H.; Heo, Y.M.; Lee, Y.M.; Lee, D.W.; Lee, H.B.; Kim, J.J. Five New Records of Soil-Derived Trichoderma in Korea: T. albolutescens, T. asperelloides, T. orientale, T. spirale, and T. tomentosum. Mycobiology 2017, 45, 1–8. [Google Scholar] [CrossRef]
- Ripa, F.A.; Cao, W.D.; Tong, S.; Sun, J.G. Assessment of plant growth promoting and abiotic stress tolerance properties of wheat endophytic fungi. BioMed Res. Int. 2019, 2019, 6105865. [Google Scholar] [CrossRef]
- Hamayun, M.; Hussain, A.; Khan, S.A.; Kim, H.Y.; Khan, A.L.; Waqas, M.; Irshad, M.; Iqbal, A.; Rehman, G.; Jan, S.; et al. Gibberellins producing endophytic fungus Porostereum spadiceum AGH786 rescues growth of salt affected soybean. Front. Microbiol. 2017, 8, 686. [Google Scholar] [CrossRef]
- Enebe, M.C.; Babalola, O.O. The influence of plant growth-promoting rhizobacteria in planttolerance to abiotic stress: A survival strategy. Appl. Microbiol. Biotechnol. 2018, 102, 7821–7835. [Google Scholar] [CrossRef]
- Hussain, S.; Khaliq, A.; Ali, B.; Hussain, H.A.; Qadir, T.; Hussain, S. Temperature extremes: Impact on rice growth and development. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; pp. 153–171. [Google Scholar]
- Mertz-Henning, L.M.; Ferreira, L.C.; Henning, F.A.; Mandarino, J.M.G.; Santos, E.D.; Oliveira, M.C.N.D.; Nepomuceno, A.E.L.; Farias, J.R.B.; Neumaier, N. Effect of water deficit-induced at vegetative and reproductive stages on protein and oil content in soybean grains. Agronomy 2018, 8, 3. [Google Scholar] [CrossRef]
- Kang, Y.; Khan, S.; Ma, X. Climate change impacts on crop yield, crop water productivity and food security—A review. Prog. Nat. Sci. 2009, 19, 1665–1674. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Khan, E.A. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants. Plant Physiol. Biochem. 2019, 135, 295–303. [Google Scholar] [CrossRef]
- Vieira, E.A.; Silva, M.G.; Moro, C.F.; Laura, V.A. Physiological and biochemical changes attenuate the effects of drought on the Cerrado species Vatairea macrocarpa (Benth.) Ducke. Plant Physiol. Biochem. 2017, 115, 472–483. [Google Scholar] [CrossRef] [PubMed]
- Kaushai, M.; Wani, S.P. Rhizobacterial-plant interactions: Strategies ensuring plant growth promotion under drought and salinity stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar] [CrossRef]
- Gray, S.B.; Brady, S.M. Plant developmental responses to climate change. Dev. Biol. 2016, 419, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, D.; Veste, M.; Boldt-Burisch, K.; Fritsch, S.; Koning, L.A.; Freese, D. Carbon allocation, nodulation, and biological nitrogen fixation of black locust (Robinia pseudoacacia L.) under soil water limitation. Ann. For. Sci. 2015, 58, 259–274. [Google Scholar] [CrossRef]
- Latif, S.; Shah, T.; Nawaz, R.; Munsif, F.; Ali, M.; ur Rehman, M.; Khan, H. Programmed Cell Death and Drought Stress Signaling. In Salt and Drought Stress Tolerance in Plants; Springer: Cham, Switzerland, 2020; pp. 211–229. [Google Scholar]
- Munnik, T.; Testerink, C. Plant phospholipid signaling: “in a nutshell”. J. Lipid Res. 2009, 50, 260–265. [Google Scholar] [CrossRef]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef]
- Andleeb, T.; Shah, T.; Nawaz, R.; Munir, I.; Munsif, F.; Jalal, A. QTL Mapping for Drought Stress Tolerance in Plants. In Salt and Drought Stress Tolerance in Plants. Signaling and Communication in Plants; Hasanuzzaman, M., Tanveer, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 383–403. [Google Scholar] [CrossRef]
- Evans, S.E.; Wallenstein, M.D.; Burke, I.C. Is bacterial moisture niche a good predictor of shifts in community composition under long-term drought? Ecology 2014, 95, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Caruso, T. Soil microbial community responses to climate extremes: Resistance, resilience and transitions to alternative states. Philos. Trans. R. Soc. B 2020, 375, 20190112. [Google Scholar] [CrossRef]
- Joshi, S.; Mohapatra, B.; Mishra, J.P.N. Microbial Soil Enzymes: Implications in the Maintenance of Rhizosphere Ecosystem and Soil Health. In Advances in Soil Microbiology: Recent Trends and Future Prospects. Microorganisms for Sustainability; Adhya, T., Lal, B., Mohapatra, B., Paul, D., Das, S., Eds.; Springer: Singapore, 2018; p. 3. [Google Scholar] [CrossRef]
- Bradáčová, K.; Sittinger, M.; Tietz, K.; Neuhäuser, B.; Kandeler, E.; Berger, N.; Ludewig, U.; Neumann, G. Maize inoculation with microbial consortia: Contrasting effects on rhizosphere activities, nutrient acquisition and early growth in different soils. Microorganisms 2019, 7, 329. [Google Scholar] [CrossRef]
- Odoh, C.K.; Sam, K.; Zabbey, N.; Eze, C.N.; Nwankwegu, A.S.; Laku, C.; Dumpe, B.B. Microbial Consortium as Biofertilizers for Crops Growing under the Extreme Habitats. In Plant Microbiomes for Sustainable Agriculture; Springer: Cham, Switzerland, 2020; pp. 381–424. [Google Scholar] [CrossRef]
- Mpanga, I.K.; Dapaah, H.K.; Geistlinger, J.; Ludewig, U.; Neumann, G. Soil type-dependent interactions of P-solubilizing microorganisms with organic and inorganic fertilizers mediate plant growth promotion in tomato. Agronomy 2018, 8, 213. [Google Scholar] [CrossRef]
- Thonar, C.; Lekfeldt, J.D.S.; Cozzolino, V.; Kundel, D.; Kulhánek, M.; Mosimann, C.; Neumann, G.; Piccolo, A.; Rex, M.; Symanczik, S.; et al. Potential of three microbial bio-effectors to promote maize growth and nutrient acquisition from alternative phosphorous fertilizers in contrasting soils. Chem. Biol. Technol. Agric. 2017, 4, 7. [Google Scholar] [CrossRef]
- Curá, J.A.; Franz, D.R.; Filosofía, J.E.; Balestrasse, K.B.; Burgueño, L.E. Inoculation with Azospirillum sp. and Herbaspirillum sp. bacteria increases the tolerance of maize to drought stress. Microorganisms 2017, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Kerry, R.G.; Patra, S.; Gouda, S.; Patra, J.K.; Das, G. Microbes and their role in drought tolerance of agricultural food crops. In Microbial Biotechnology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 253–273. [Google Scholar] [CrossRef]
- Almaroai, Y.A.; Eissa, M.A. Role of marine algae extracts in water stress resistance of onion under semiarid conditions. J. Soil Sci. Plant Nutr. 2020, 44, 1092–1101. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H. Mechanistic insights into arbuscular mycorrhizal fungi-mediated drought stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef]
- Ghorbanpour, A.; Salimi, A.; Ghanbary, M.A.T.; Pirdashti, H.; Dehestani, A. The effect of Trichoderma harzianum in mitigating low temperature stress in tomato (Solanum lycopersicum L.) plants. Sci. Hortic. 2018, 230, 134–141. [Google Scholar] [CrossRef]
- Vaishnav, A.; Shukla, A.K.; Sharma, A.; Kumar, R.; Choudhary, D.K. Endophytic bacteria in plant salt stress tolerance: Current and future prospects. J. Plant Growth Regul. 2019, 38, 650–668. [Google Scholar] [CrossRef]
- Chhabra, R. Soil Salinity and Water Quality; Routledge: London, UK, 2017. [Google Scholar] [CrossRef]
- Qayyum, F.; Saifullah, M.S.; Akhtar, J. Salt-affected soils: Sources genesis and management. In Soil Science Concepts and Applications; Sabir, M., Akhtar, J., Hakeem, K.R., Eds.; University of Agriculture Faisalabad: Faisalabad, Pakistan, 2016; Available online: https://www.researchgate.net/profile/Muhammad-Qayyum-6/publication/320583309_Salt-affected_Soils_Sources_Genesis_and_Management/links/59eec988aca272029ddf802f/Salt-affected-Soils-Sources-Genesis-and-Management.pdf (accessed on 25 February 2023).
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef]
- Lima, G.S.; Santos, J.B.; Soares, L.A.A.; Gheyi, H.R.; Nobre, R.G.; Pereira, R.F. Irrigation with saline water and application of leaf proline in pepper cultivation. ‘All Big’. Comun. Sci. 2016, 7, 513–522. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Safdar, H.; Amin, A.; Shafiq, Y.; Ali, A.; Yasin, R.; Shoukat, A.; Hussan, M.U.; Sarwar, M.I. A review: Impact of salinity on plant growth. Nat. Sci. 2019, 17, 34–40. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Kader, M.A.L.; Lindberg, S. Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal Behav. 2010, 5, 233–238. [Google Scholar] [CrossRef]
- Pérez-Alfocea, F.; Albacete, A.; Ghanem, M.E.; Dodd, I.C. Hormonal regulation of source-sink relations to maintain crop productivity under salinity: A case study of root-to-shoot signalling in tomato. Funct. Plant Biol. 2010, 37, 592–603. [Google Scholar] [CrossRef]
- Ben Salah, I.; Albacete, A.; Messedi, D.; Gandour, M.; Andújar, C.M.; Zribi, K.; Martinez, V.; Abdelly, C. Hormonal responses of nodulated Medicago ciliaris lines differing in salt tolerance. Environ. Exp. Bot. 2013, 86, 35–43. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Albacete, A.; Martínez-Andújar, C.; Acosta, M.; Romero-Aranda, R.; Dodd, I.C.; Lutts, S.; Pérez-Alfocea, F. Hormonal changes during salinity induced leaf senescence in tomato (Solanum lycopersicum L.). J. Exp. Bot. 2008, 59, 3039–3050. [Google Scholar] [CrossRef]
- Orhan, F. Alleviation of salt stress by halotolerant and halophilic plant growth-promoting bacteria in wheat (Triticum aestivum). Braz. J. Microbiol. 2016, 47, 621–627. [Google Scholar] [CrossRef]
- Mishra, J.; Singh, R.; Arora, N.K. Alleviation of heavy metal stress in plants and remediation of soil by rhizosphere microorganisms. Front. Microbiol. 2017, 8, 1706. [Google Scholar] [CrossRef]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; AL-Harrasi, A. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Al-Garni, S.M.S.; Khan, M.D.M.A.; Bahieldin, A. Plant growth-promoting bacteria and silicon fertilizer enhance plant growth and salinity tolerance in Coriandrum sativum. J. Plant Interact. 2019, 14, 386–396. [Google Scholar] [CrossRef]
- Etesami, H.; Glick, B.R. Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environ. Exp. Bot. 2020, 173, 104124. [Google Scholar] [CrossRef]
- Lenoir, I.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Arbuscular mycorrhizal fungal responses to abiotic stresses: A review. Photochemistry 2016, 123, 4–15. [Google Scholar] [CrossRef]
- Guerrero-Galán, C.; Calvo-Polanco, M.; Zimmermann, S.D. Ectomycorrhizal symbiosis helps plants to challenge salt stress conditions. Mycorrhiza 2019, 29, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J. Trichoderma parareesei favors the tolerance of rapeseed (Brassica napus L.) to salinity and drought due to a chorismate mutase. Agronomy 2020, 10, 118. [Google Scholar] [CrossRef]
- Alori, E.T.; Dare, M.O.; Babalola, O.O. Microbial inoculants for soil quality and plant health. In Sustainable Agriculture Reviews; Springer: Cham, Switzerland, 2017; pp. 281–307. [Google Scholar] [CrossRef]
- Teixeira Filho, M.C.M.; Galindo, F.S. Inoculation of bacteria with a focus on biological nitrogen fixation and promotion of plant growth. In Topics in Soil Science—Volume X; Severiano, E.C., Moraes, M.F., Paula, A.M., Eds.; Brazilian Society of Soil Science: Brasilia, Brazil, 2019; Chapter 11; pp. 577–648. [Google Scholar]
- Galindo, F.S.; Teixeira Filho, M.C.M.; Buzetti, S.; Santini, J.M.K.; Alves, C.J.; Nogueira, L.M.; Ludkiewicz, M.G.Z.; Andreotti, M.; Bellotte, J.L.M. Corn yield and foliar diagnosis affected by nitrogen fertilization and inoculation with Azospirillum brasilense. Rev. Bras. De Ciência Do Solo 2016, 40, e0150364. [Google Scholar] [CrossRef]
- Jimtha John, C.; Jishma, P.; Karthika, N.R.; Nidheesh, K.S.; Ray, J.G.; Mathew, J.; Radhakrishnan, E.K. Pseudomonas fluorescens R68 assisted enhancement in growth and fertilizer utilization of Amaranthus tricolor (L.). Biotechnology 2017, 7, 256. [Google Scholar] [CrossRef]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Lajayer, B.A.; Ghorbanpour, M.; Nikabadi, S. Heavy metals in contaminated enviroment: Destiny of secondary metabolite biosynthesis, oxidative status and phytoextraction in medicinal plants. Ecotoxicol. Environ. Saf. 2017, 145, 377–390. [Google Scholar] [CrossRef]
- Dinis, M.D.L.; Fiúza, A. Exposure assessment to heavy metals in the environment: Measures to eliminate or reduce the exposure to critical receptors. Environ. Heavy Met. Pollut. Eff. Child Ment. Dev. 2011, 1, 27–50. [Google Scholar] [CrossRef]
- Shahid, M.; Dumat, C.; Khalid, S.; Schreck, E.; Xiong, T.; Niazi, N.K. Foliar heavy metal uptake, toxicity and detoxification in plants: A comparison of foliar and root metal uptake. J. Hazard. Mater. 2017, 325, 36–58. [Google Scholar] [CrossRef]
- Manoj, S.R.; Karthik, C.; Kadirvelu, K.; Arulselvi, P.I.; Shanmugasundaram, T.; Bruno, B.; Rajkumar, M. Understanding the molecular mechanisms for the enhanced phytoremediation of heavy metals through plant growth promoting rhizobacteria: A review. J. Environ. Manag. 2020, 254, 109779. [Google Scholar] [CrossRef]
- Cambrollé, J.; Mancilla-Leytón, J.M.; Muñoz-Vallés, S.; Luque, T.; Figueroa, M.E. Zinc tolerance and accumulation in the salt-marsh shrub Halimione portulacoides. Chemosphere 2012, 86, 867–874. [Google Scholar] [CrossRef]
- Viehweger, K.; Geipel, G. Uranium accumulation and tolerance in Arabidopsis halleri under native versus hydroponic conditions. Environ. Exp. Bot. 2010, 69, 39–46. [Google Scholar] [CrossRef]
- Viehweger, K. How plants cope with heavy metals. Bot. Stud. 2014, 55, 39–46. [Google Scholar] [CrossRef]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Pires, C.; Franco, A.R.; Pereira, S.I.A.; Henriques, I.; Correia, A.; Magan, N.; Castro, P.M.L. Metal(loid)-contaminated soils as a source of culturable heterotrophic aerobic bacteria for remediation applications. Geomicrobiol. J. 2017, 34, 760–768. [Google Scholar] [CrossRef]
- Hadi, F.; Bano, A. Effect of diazotrophs (Rhizobium and Azobactor) on growth of maize (Zea mays L.) and accumulation of Lead (Pb) in different plant parts. Pak. J. Bot. 2010, 42, 4363–4370. [Google Scholar]
- Ma, Y.; Rajkumar, M.; Luo, Y.; Freitas, H. Inoculation of endophytic bacteria on host and non-host plants-effects on plant growth and Ni uptake. J. Hazard Mater. 2011, 195, 230–237. [Google Scholar] [CrossRef]
- Coelho, L.M.; Rezende, H.C.; Coelho, L.M.; de Sousa, P.A.; Melo, D.F.; Coelho, N.M. Bioremediation of polluted waters using microorganisms. In Advances in Bioremediation of Wastewater and Polluted Soil; Shiomi, N., Ed.; In Tech: Shanghai, China, 2015. [Google Scholar]
- Tiwari, S.; Singh, S.N.; Garg, S.K. Stimulated phytoextraction of metals from fly ash by microbial interventions. Environ. Technol. 2012, 33, 2405–2413. [Google Scholar] [CrossRef]
- Babu, A.G.; Kim, J.D.; Oh, B.T. Enhancement of heavy metal phytoremediation by Alnus firma with endophytic Bacillus thuringiensis GDB-1. J. Hazard Mater. 2013, 250, 477–483. [Google Scholar] [CrossRef]
- Dong, G.; Wang, Y.; Gong, L.; Wang, M.; Wang, H.; He, N.; Zheng, Y.; Li, Q. Formation of soluble Cr (III) end-products and nanoparticles during Cr (VI) reduction by bacillus cereus strain XMCr-6. Biochem. Eng. J. 2013, 70, 166–172. [Google Scholar] [CrossRef]
- Balamurugan, D.; Udayasooriyan, C.; Kamaladevi, B. Chromium (VI) reduction by Pseudomonas putida and Bacillus subtilis isolated from contaminated soils. Int. J. Environ. Sci. 2014, 5, 522. [Google Scholar] [CrossRef]
- Chen, L.; Luo, S.; Li, X.; Wan, Y.; Chen, J.; Liu, C. Interaction of Cd hyperaccumulator Solanum nigrum L. and functional endophyte Pseudomonas sp. Lk9 on soil heavy metals uptake. Soil Biol. Biochem. 2014, 68, 300–308. [Google Scholar] [CrossRef]
- Jing, Y.X.; Yan, J.L.; He, H.D.; Yang, D.J.; Xiao, L.; Zhong, T.; Yuan, M.; Cai, X.D.; Li, S.B. Characterization of bacteria in the rhizosphere soils of Polygonum pubescens and their potential in promoting growth and Cd Pb, Zn uptake by Brassica napus. Int. J. Phytoremediat. 2014, 16, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Oliviera, R.S.; Nai, F.; Rajkumar, M.; Luo, Y.; Rocha, I.; Freitas, H. The hyperaccumulator Sedum plumbizincicola harbors metal-resistant endophytic bacteria that improve its phytoextraction capacity in multi-metal contaminated soil. J. Environ. Manag. 2015, 156, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Nahar, N.; Nawani, N.N.; Jass, J.; Hossain, K.; Saud, Z.A.; Saha, A.K.; Ghosh, S.; Olsson, B.; Mandal, A. Bioremediation of hexavalent chromium (VI) by a soil-borne bacterium, Enterobacter cloacae b2-dha. J. Environ. Sci. Health Part A 2015, 50, 1136–1147. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Modulation of phytoremediation and plant growth by the treatment with PGPR, Ag nanoparticle and untreated municipal wastewater. Int. J. Phytoremediat. 2016, 18, 1258–1269. [Google Scholar] [CrossRef]
- Tamoor, U.H.; Bano, A.; Naz, I. Alleviation of heavy metals toxicity by the application of plant growth promoting rhizobacteria and effects on wheat grown in saline sodic field. Int. J. Phytoremediat. 2017, 19, 522–529. [Google Scholar] [CrossRef]
- Seyed, M.M.; Babak, M.; Hossein, M.H.; Hoseinali, A.; Ali, A.Z. Root-induced changes of Zn and Pb dynamics in the rhizosphere of sunflower with different plant growth promoting treatments in a heavily contaminated soil. Ecotoxicol. Environ. Saf. 2018, 147, 206–216. [Google Scholar] [CrossRef]
- Das, D.; Das, N.; Mathew, L. Kinetics, equilibrium and thermodynamic studies on biosorption of AG(I) from aqueous solution by macrofungus Pleurotus platypus. J. Hazard Mater. 2010, 184, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Sukumar, M. Reduction of hexavalent chromium by Rhizopus oryzae. Afr. J. Environ. Sci. Technol. 2010, 4, 412–418. [Google Scholar]
- Kumar Ramasamy, R.; Congeevaram, S.; Thamaraiselvi, K. Evaluation of isolated fungal strain from e-waste recycling facility for effective sorption of toxic heavy metal Pb (II) ions and fungal protein molecular characterization—A mycoremediation approach. Asian J. Exp Biol. Sci. 2011, 2, 342–347. [Google Scholar]
- Fu, Y.Q.; Li, S.; Zhu, H.Y.; Jiang, R.; Yin, L.F. Biosorption of copper (II) from aqueous solution by mycelial pellets of Rhizopus oryzae. Afr. J. Biotechnol. 2012, 11, 1403–1411. [Google Scholar] [CrossRef]
- Mane, P.; Bhosle, A. Bioremoval of some metals by living algae Spirogyra sp. and Spirullina sp. from aqueous solution. Int. J. Environ. Res. 2012, 6, 571–576. [Google Scholar] [CrossRef]
- Yalçın, S.; Sezer, S.; Apak, R. Characterization and lead(II), cadmium(II), nickel(II) biosorption of dried marine brown macro algae Cystoseira barbata. Environ. Sci. Pollut. Res. 2012, 19, 3118–3125. [Google Scholar] [CrossRef]
- Srivastava, S.; Agrawal, S.; Mondal, M. A review on progress of heavy metal removal using adsorbents of microbial and plant origin. Environ. Sci. Pollut. Res. 2015, 22, 15386–15415. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Miao, C.; Hanel, M.; Borthwick, A.G.; Duan, Q.; Ji, D.; Li, H. Global heat stress on health, wildfires, and agricultural crops under different levels of climate warming. Environ. Int. 2019, 128, 125–136. [Google Scholar] [CrossRef]
- Deryng, D.; Conway, D.; Ramankutty, N.; Price, J.; Warren, R. Global crop yield response to extreme heat stress under multiple climate change futures. Environ. Res. Lett. 2014, 9, 034011. [Google Scholar] [CrossRef]
- Abd El-Daim, I.A.; Bejai, S.; Meijer, J. Improved heat stress tolerance of wheat seedlings by bacterial seed treatment. Plant Soil 2014, 379, 337–350. [Google Scholar] [CrossRef]
- Liu, B.; Liu, L.; Tian, L.; Cao, W.; Zhu, Y.; Asseng, S. Post-heading heat stress and yield impact in winter wheat of China. Glob. Chang. Biol. 2014, 20, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Uchida, S. Quality Matters more than quantity: Asymmetric temperature effects on crop yield and quality grade. Am. J. Agric. Econ. 2016, 98, 1195–1209. [Google Scholar] [CrossRef]
- Osman, R.; Zhu, Y.; Ma, W.; Zhang, D.; Ding, Z.; Liu, L.; Tang, L.; Liu, B.; Cao, W. Comparison of wheat simulation models for impacts of extreme temperature stress on grain quality. Agric. For. Meteorol. 2020, 288, 107995. [Google Scholar] [CrossRef]
- Wang, E.; Martre, P.; Zhao, Z.; Ewert, F.; Maiorano, A.; Rötter, R.P.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. The uncertainty of crop yield projections is reduced by improved temperature response functions. Nat. Plants 2017, 3, 17102. [Google Scholar] [CrossRef]
- Webber, H.; Martre, P.; Asseng, S.; Kimball, B.; White, J.; Ottman, M.; Wall, G.W.; De Sanctis, G.; Doltra, J.; Grant, R.; et al. Canopy temperature for simulation of heat stress in irrigated wheat in a semi-arid environment: A multi-model comparison. Field Crops Res. 2017, 202, 21–35. [Google Scholar] [CrossRef]
- Kong, L.; Ashraf, U.; Cheng, S.; Rao, G.; Mo, Z.; Tian, H.; Pan, S.; Tang, X. Short-term water management at early filling stage improves early-season rice performance under high temperature stress in South China. Eur. J. Agron. 2017, 90, 117–126. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Perumal, R.; Jagadish, S.V.K.; Ciampitti, I.A.; Welti, R.; Prasad, P.V.V. Sensitivity of sorghum pollen and pistil to high-temperature stress. Plant Cell Environ. 2018, 41, 1065–1082. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Perumal, R.; Ciampitti, I.A.; Gupta, S.K.; Prasad, P.V.V. Quantifying pearl millet response to high temperature stress: Thresholds, sensitive stages, genetic variability and relative sensitivity of pollen and pistil. Plant Cell Environ. 2018, 41, 993–1007. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Narayanan, S.; Erdayani, E.; Prasad, P.V. Effects of high temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol. 2020, 20, 268. [Google Scholar] [CrossRef]
- Zhang, S.; He, Y.; Sen, B.; Wang, G. Reactive oxygen species and their applications toward enhanced lipid accumulation in oleaginous microorganisms. Bioresour. Technol. 2020, 307, 123234. [Google Scholar] [CrossRef]
- Bhattacharjee, S. ROS and oxidative stress: Origin and implication. In Reactive Oxygen Species in Plant Biology; Springer: New Delhi, India, 2019. [Google Scholar] [CrossRef]
- Tripathy, B.C.; Oelmuller, R. Reactive oxygen species generation and signalling in plants. Plant Signal. Behaviour. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Yalcinkaya, T.; Uzilday, B.; Ozgur, R.; Turkan, I.; Mano, J. Lipid peroxidation-derived reactive carbonyl species (RCS): Their interaction with ROS and cellular redox during environmental stresses. Environ. Exp Bot. 2019, 165, 139–149. [Google Scholar] [CrossRef]
- Zhou, R.; Kong, L.; Yu, X.; Ottosen, C.O.; Zhao, T.; Jiang, F.; Wu, Z. Oxidative damage and antioxidant mechanism in tomatoes responding to drought and heat stress. Acta Physiol. Plant 2019, 41, 20. [Google Scholar] [CrossRef]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Kumar, M.; Kour, D.; Yadav, A.N.; Saxena, R.; Rai, P.K.; Jyoti, A.; Tomar, R.S. Biodiversity of methylotrophic microbial communities and their potential role in mitigation of abiotic stresses in plants. Biologia 2019, 74, 287–308. [Google Scholar] [CrossRef]
- Amin, A.; Ahmed, I.; Salam, N.; Kim, B.-Y.; Singh, D.; Zhi, X.-Y.; Xiao, M.; Li, W.-J. Diversity and distribution of thermophilic bacteria in hot springs of Pakistan. Microbiol. Ecol. 2017, 74, 116–127. [Google Scholar] [CrossRef]
- Chanratana, M.; Han, G.H.; Roy Choudhury, A.; Sundaram, S.; Halim, M.A.; Krishnamoorthy, R.; Kang, Y.; Sa, T. Assessment of Methylobacterium oryzae CBMB20 aggregates for salt tolerance and plant growth promoting characteristics for bio-inoculant development. AMB Express 2017, 7, 208. [Google Scholar] [CrossRef]
- Cassán, F.; Maiale, S.; Masciarelli, O.; Vidal, A.; Luna, V.; Ruiz, O. Cadaverine production by Azospirillum brasilense and its possible role in plant growth promotion and osmotic stress mitigation. Eur. J. Soil Biol. 2009, 45, 12–19. [Google Scholar] [CrossRef]
- Puente, M.; Li, C.; Bashan, Y. Microbial populations and activities in the rhizoplane of rock-weathering desert plants. II. Growth promotion of cactus seedlings. Plant Biol. 2004, 6, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Rosa, P.A.L.; Mortinho, E.S.; Jalal, A.; Galindo, F.S.; Buzetti, S.; Fernandes, G.C.; Barco Neto, M.; Pavinato, P.S.; Teixeira Filho, M.C.M. Inoculation with growth-promoting bacteria associated with the reduction of phosphate fertilization in sugarcane. Front. Environ. Sci. 2020, 8, 32. [Google Scholar] [CrossRef]
- Rosa, P.A.L.; Galindo, F.S.; Oliveira, C.E.D.S.; Jalal, A.; Mortinho, E.S.; Fernandes, G.C.; Marega, E.M.R.; Buzetti, S.; Teixeira Filho, M.C.M. Inoculation with plant growth-promoting bacteria to reduce phosphate fertilization requirement and enhance technological quality and yield of sugarcane. Microorganisms 2022, 10, 192. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, Z.M.; Ahmada, M.; Jamil, M.; Hussain, T. Zinc solubilizing Bacillus spp. potential candidates for biofortification in maize. Microbiol. Res. 2017, 202, 51–60. [Google Scholar] [CrossRef]
- Jalal, A.; da Silva Oliveira, C.E.; Freitas, L.A.; Galindo, F.S.; Lima, B.H.; Boleta, E.H.M.; Da Silva, E.C.; do Nascimento, V.; Nogueira, T.A.R.; Buzetti, S.; et al. Agronomic biofortification and productivity of wheat with soil zinc and diazotrophic bacteria in tropical savannah. Crop Pasture Sci. 2022, 73, 817–830. [Google Scholar] [CrossRef]
- Jalal, A.; Oliveira, C.E.D.S.; Fernandes, H.B.; Galindo, F.S.; Silva, E.C.D.; Fernandes, G.C.; Nogueira, T.A.R.; De Carvalho, P.H.G.; Balbino, V.R.; Lima, B.H.D.; et al. Diazotrophic bacteria is an alternative strategy for increasing grain biofortification, yield and zinc use efficiency of maize. Plants 2022, 11, 1125. [Google Scholar] [CrossRef]
- Jalal, A.; Oliveira, C.E.D.S.; Bastos, A.; Fernandes, G.; de Lima, B.; de Carvalho, P.; Galindo, F.; Gato, I.; Teixeira Filho, M. Nanozinc and plant growth-promoting bacteria improve biochemical and metabolic attributes of maize in tropical Cerrado. Front. Plant Sci. 2022, 13, 1046642. [Google Scholar] [CrossRef]
- Fukami, J.; Ollero, F.J.; Megías, M.; Hungria, M. Phytohormones and induction of plant stress tolerance and defense genes by seed and foliar inoculation with Azospirillum brasilense cells and metabolites promote maize growth. AMB Express 2017, 7, 153. [Google Scholar] [CrossRef]
- Passari, A.K.; Mishra, V.K.; Singh, G.; Singh, P.; Kumar, B.; Gupta, V.K.; Sarma, R.K.; Saikia, R.; Donovan, A.O.; Singh, B.P. Insights into the functionality of endophytic actinobacteria with a focus on their biosynthetic potential and secondary metabolites production. Sci. Rep. 2017, 7, 11809. [Google Scholar] [CrossRef]
- Tiwari, S.; Prasad, V.; Chauhan, P.S.; Lata, C. Bacillus amyloliquefaciens confers tolerance to various abiotic stresses and modulates plant response to phytohormones through osmoprotection and gene expression regulation in rice. Front. Plant Sci. 2017, 8, 1510. [Google Scholar] [CrossRef]
- Bruno, L.B.; Karthik, C.; Ma, Y.; Kadirvelu, K.; Freitas, H.; Rajkumar, M. Amelioration of chromium and heat stresses in Sorghum bicolor by Cr6+ reducing-thermotolerant plant growth promoting bacteria. Chemosphere 2020, 244, 125521. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Wu, X.; Zhang, J.; Huang, C. High-temperature induced changes of extracellular metabolites in Pleurotus ostreatus and their positive effects on the growth of Trichoderma asperellum. Front. Microbiol. 2018, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Daim, I.A.; Bejai, S.; Fridborg, I.; Meijer, J. Identifying potential molecular factors involved in Bacillus amyloliquefaciens 5113 mediated abiotic stress tolerance in wheat. Plant Biol. J. 2018, 20, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Janda, M.; Lamparova, L.; Zubijova, A.; Burketova, L.; Martinec, J.; Krckova, Z. Temporary heat stress suppresses PAMP-triggered immunity and resistance to bacteria in Arabidopsis thaliana. Mol. Plant Pathol. 2019, 20, 1005–1012. [Google Scholar] [CrossRef]
- Shekhawat, K.; Sheikh, A.; Mariappan, K.; Jalal, R.; Hirt, H. Enterobacter sp. SA187 mediates plant thermotolerance by chromatin modification of heat stress genes. bioRxiv 2019. [Google Scholar] [CrossRef]
- Abd El-Daim, I.A.; Bejai, S.; Meijer, J. Bacillus velezensis 5113 induced metabolic and molecular reprogramming during abiotic stress tolerance in wheat. Sci. Rep. 2019, 9, 16282. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.-M.; Kim, K.-M.; Lee, I.J. Extending thermotolerance to tomato seedlings by inoculation with SA1 isolate of Bacillus cereus and comparison with exogenous humic acid application. PLoS ONE 2020, 15, e0232228. [Google Scholar] [CrossRef]
- Mukhtar, T.; Regman, S.U.; Smith, D.; Sultan, T.; Seleiman, M.F.; Alsadon, A.A.; Amna; Ali, S.; Chaudhary, H.J.; Solieman, T.H.I.; et al. Mitigation of Heat Stress in Solanum lycopersicum L. by ACC-deaminase and Exopolysaccharide Producing Bacillus cereus: Effects on Biochemical Profiling. Sustainability 2020, 12, 2159. [Google Scholar] [CrossRef]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Panjiar, N.; Kumar, S.; Saxena, A.K.; Suman, A. Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Ann. Microbiol. 2015, 65, 1885–1899. [Google Scholar] [CrossRef]
- Subramanian, P.; Kim, K.; Krishnamoorthy, R.; Mageswari, A.; Selvakumar, G.; Sa, T. Cold stress tolerance in psychrotolerant soil bacteria and their conferred chilling resistance in tomato (Solanum lycopersicum Mill.) under low temperatures. PLoS ONE 2016, 11, e0161592. [Google Scholar] [CrossRef]
- Zubair, M.; Hanif, A.; Farzand, A.; Sheikh, T.M.M.; Khan, A.R.; Suleman, M.; Ayaz, M.; Gao, X. Genetic screening and expression analysis of psychrophilic Bacillus spp. reveal their potential to alleviate cold stress and modulate phytohormones in wheat. Microorganisms 2019, 7, 337. [Google Scholar] [CrossRef]
- Tiryaki, D.; Aydın, İ.; Atıcı, Ö. Psychrotolerant bacteria isolated from the leaf apoplast of cold-adapted wild plants improve the cold resistance of bean (Phaseolus vulgaris L.) under low temperature. Cryobiology 2019, 86, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, A.; Rehman, S.; Salavati, A.; Komatsu, S. A comparative proteomics analysis in roots of soybean to compatible symbiotic bacteria under flooding stress. Amino Acids 2012, 43, 2513–2525. [Google Scholar] [CrossRef] [PubMed]
- Barnawal, D.; Bharti, N.; Maji, D.; Chanotiya, C.S.; Kalra, A. 1-Aminocyclopropane-1-carboxylic acid (ACC) deaminase-containing rhizobacteria protect Ocimum sanctum plants during waterlogging stress via reduced ethylene generation. Plant Physiol. Biochar 2012, 58, 227–235. [Google Scholar] [CrossRef]
- Li, J.; McConkey, B.J.; Cheng, Z.; Guo, S.; Glick, B.R. Identification of plant growth-promoting bacteria-responsive proteins in cucumber roots under hypoxic stress using a proteomic approach. J. Proteom. 2013, 84, 119–131. [Google Scholar] [CrossRef]
- Ferrando, L.; Fernández Scavino, A. Strong shift in the diazotrophic endophytic bacterial community inhabiting rice (Oryza sativa) plants after flooding. FEMS Microbiol. Ecol. 2015, 91, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Ravanbakhsh, M.; Sasidharan, R.; Voesenek, L.A.C.J.; Kowalchuk, G.A.; Jousset, A. ACC deaminase-producing rhizosphere bacteria modulate plant responses to flooding. J. Ecol. 2015, 105, 979–986. [Google Scholar] [CrossRef]
- Duc, N.H.; Csintalan, Z.; Posta, K. Arbuscular mycorrhizal fungi mitigate negative effects of combined drought and heat stress on tomato plants. Plant Physiol. Biochem. 2018, 132, 297–307. [Google Scholar] [CrossRef]
- Ismail, H.M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I.J. Endophytic fungus Aspergillus japonicus mediates host plant growth under normal and heat stress conditions. BioMed Res. Int. 2018, 2018, 7696831. [Google Scholar] [CrossRef]
- Ali, A.H.; Abdelrahman, M.; Radwan, U.; El-Zayat, S.; El-Sayed, M.A. Effect of Thermomyces fungal endophyte isolated from extreme hot desert-adapted plant on heat stress tolerance of cucumber. Appl. Soil Ecol. 2018, 124, 155–162. [Google Scholar] [CrossRef]
- Ali, A.H.; Radwan, U.; El-Zayat, S.; El-Sayed, M.A. The role of the endophytic fungus, Thermomyces lanuginosus, on mitigation of heat stress to its host desert plant Cullen plicata. Biol. Futur. 2019, 70, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.T.; Fu, J.J.; Sun, Y.F.; Xu, Y.M.; Miao, Y.J.; Xu, Y.F.; Hu, T.M. Effect of arbuscular mycorrhizal fungi inoculation on cold stress-induced oxidative damage in leaves of Elymus nutans Griseb. S. Afr. J. Bot. 2016, 104, 21–29. [Google Scholar] [CrossRef]
- Hajiboland, R.; Joudmand, A.; Aliasgharzad, N.; Tolrá, R.; Poschenrieder, C. Arbuscular mycorrhizal fungi alleviate low-temperature stress and increase freezing resistance as a substitute for acclimation treatment in barley. Crop Pasture Sci. 2019, 70, 218–233. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Molina, S.; Zamarreño, A.M.; García-Mina, J.M.; Aroca, R. The symbiosis with the arbuscular mycorrhizal fungus Rhizophagus irregularis drives root water transport in flooded tomato plants. Plant Cell Physiol. 2014, 55, 1017–1029. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.M.; Wang, P.T.; Wang, S.G. Effect of Flooding Time Length on Mycorrhizal Colonization of Three AM Fungi in Two Wetland Plants. Huan Jing Ke Xue 2014, 35, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Khadka, R.B.; Uphoff, N. Effects of Trichoderma seedling treatment with system of rice intensification management and with conventional management of transplanted rice. PeerJ 2019, 7, e5877. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Gao, Y.; Liu, C.; Liu, S. A styrene antioxidant NFA from riparian endophytic fungi enhances flooding tolerance in Arabidopsis. J. Plant Interact. 2020, 15, 111–116. [Google Scholar] [CrossRef]
- Meena, R.S.; Vijayakumar, V.; Yadav, G.S.; Mitran, T. Response and interaction of Bradyrhizobium japonicum and arbuscular mycorrhizal fungi in the soybean rhizosphere. Plant Growth Regul. 2018, 84, 207–223. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Cotton, J.; Gardner, T.; Moore-Kucera, J.; Zak, J.; Wester, D.; Cox, S. Predominant bacterial and fungal assemblages in agricultural soils during a record drought/heat wave and linkages to enzyme activities of biogeochemical cycling. Appl. Soil Ecol. 2014, 84, 69–82. [Google Scholar] [CrossRef]
- Sati, P.; Dhakar, K.; Pandey, A. Microbial diversity in soil under potato cultivation from cold desert Himalaya, India. Int. Sch. Res. Not. 2013, 2013, 767453. [Google Scholar] [CrossRef]
- Caradonia, F.; Francia, E.; Morcia, C.; Ghizzoni, R.; Moulin, L.; Terzi, V.; Ronga, D. Arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria avoid processing tomato leaf damage during chilling stress. Agronomy 2013, 9, 299. [Google Scholar] [CrossRef]
- Walker, L.R. The Biology of Disturbed Habitats; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- González Macé, O.; Steinauer, K.; Jousset, A.; Eisenhauer, N.; Scheu, S. Flood-induced changes in soil microbial functions as modified by plant diversity. PLoS ONE 2016, 11, e0166349. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Ma, C.; Chen, H.; Chen, G.; White, J.C.; Xing, B. Copper stress in flooded soil: Impact on enzyme activities, microbial community composition and diversity in the rhizosphere of Salix integra. Sci. Total Environ. 2020, 704, 135350. [Google Scholar] [CrossRef] [PubMed]
- Hartman, K.; Tringe, S.G. Interactions between plants and soil shaping the root microbiome under abiotic stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [PubMed]
- Colmer, T.D. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 2003, 26, 17–36. [Google Scholar] [CrossRef]
- Thomas, A.L.; Guerreiro, S.M.C.; Sodek, L. Aerenchyma formation and recovery from hypoxia of the flooded root system of nodulated soybean. Ann. Bot. 2005, 96, 1191–1198. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.A.; Rainbolt, C.R.; Morris, D.R.; Bennett, A.C. Morphological responses of sugarcane to long-term flooding. Agron. J. 2005, 99, 1622–1628. [Google Scholar] [CrossRef]
- Yamauchi, T.; Shimamura, S.; Nakazono, M.; Mochizuki, T. Aerenchyma formation in crop species: A review. Field Crop Res. 2013, 152, 8–16. [Google Scholar] [CrossRef]
- López-Tarazón, J.A.; Bronstert, A.; Thieken, A.H.; Petrow, T. The effects of global change on floods, fluvial geomorphology and related hazards in mountainous rivers. Sci. Total Environ. 2019, 669, 7–10. [Google Scholar] [CrossRef]
- Mentzer, J.L.; Goodman, R.M.; Balser, T.C. Microbial response over time to hydrologic and fertilization treatments in a simulated wet prairie. Plant Soil 2006, 284, 85–100. [Google Scholar] [CrossRef]
- Unger, I.M.; Kennedy, A.C.; Muzika, R.M. Flooding effects on soil microbial communities. Appl. Soil Ecol. 2009, 42, 1–8. [Google Scholar] [CrossRef]
- Gamalero, E.; Glick, B.R. Bacterial modulation of plant ethylene levels. Plant Physiol. 2015, 169, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Grichko, V.P.; Glick, B.R. Amelioration of flooding stress by ACC deaminase-containing plant growth-promoting bacteria. Plant Physiol. Biochnol. 2001, 39, 11–17. [Google Scholar] [CrossRef]
- Farwell, A.J.; Vesely, S.; Nero, V.; Rodriguez, H.; McCormack, K.; Shah, S.; Dixon, D.G.; Glick, B.R. Tolerance of transgenic canola plants (Brassica napus) amended with plant growth-promoting bacteria to flooding stress at a metal-contaminated field site. Environ. Pollut. 2007, 147, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Müller-Xing, R.; Xing, Q.; Goodrich, J. Footprints of the sun: Memory of UV and light stress in plants. Front Plant Sci. 2014, 5, 474. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.; Nieuwendijk, N.M.; Pantazopoulou, C.K.; Pierik, R. Light signalling shapes plant–plant interactions in dense canopies. Plant Cell Environ. 2021, 44, 1014–1029. [Google Scholar] [CrossRef]
- Takahashi, S.; Badger, M.R. Photoprotection in plants: A new light on photosystem II damage. Trends Plant Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef]
- Praeg, N.; Illmer, P. Microbial community composition in the rhizosphere of Larix decidua under different light regimes with additional focus on methane cycling microorganisms. Sci. Rep. 2020, 10, 22324. [Google Scholar] [CrossRef]
- Stefan, M.; Munteanu, N.; Stoleru, V.; Mihasan, M.; Hritcu, L. Seed inoculation with plant growth promoting rhizobacteria enhances photosynthesis and yield of runner bean (Phaseolus coccineus L.). Sci. Hortic. 2014, 151, 22–29. [Google Scholar] [CrossRef]
- Suzuki, W.; Sugawara, M.; Miwa, K.; Morikawa, M. Plant growth-promoting bacterium Acinetobacter calcoaceticus P23 increases the chlorophyll content of the monocot Lemna minor (duckweed) and the dicot Lactuca sativa (lettuce). J. Biosci. Bioeng. 2014, 118, 41–44. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Microorganism | Stress | Plant Species | References |
|---|---|---|---|
| Bacteria | |||
| Azospirillum brasilense | Drought | Marandu grass (Urochloa brizantha) | [34] |
| PGPRs strain IG 3, Enterobacter ludwigii, and Flavobacterium sp. | Drought | Wheat (Triticum aestivum) | [35] |
| Bacillus sp. | Drought | Sugarcane (Saccharum spp.) | [36] |
| Bacillus megaterium, B. subtilis, and Bacillus thuringiensis | Drought | Wheat (Triticum aestivum L.) and chickpea (CicerArietinum) | [37] |
| Bacillus sp. (12D6) and Enterobacter sp. (16i) | Drought | Wheat (Triticum aestivum) and maize (Zea mays) | [38] |
| Actinobacterium | Drought | Maize (Zea mays L.) | [39] |
| Proteobacteria, Actinobacteria, Gemmatimonadetes, Chloroflexi, Cyanobacteria, and Acidobacteria | Drought | Cotton (Gossypium hirsutum) | [40] |
| Bradyrhizobium japonicum and Azospirillum brasilense | Drought | Soybean (Glycine max) | [41] |
| Acinetobacter calcoaceticus EU- LRNA-72 and Penicillium sp. EU-FTF-6 | Drought | Foxtail millet (Setaria italica L.) | [42] |
| Pseudomonas lini, Bacillus, and Serratia plymuthica | Drought | Jujube (Ziziphus jujuba) | [43] |
| Rhizobium tropici and Azospirillum brasilense | Drought | Common bean (Phaseolus vulgaris) | [44] |
| Azotobacter chroococcum | Salt | Tomato (Solanum lycopersicum) | [45] |
| Microbacterium oleivorans, Brevibacterium iodinum, and Rhizobium massiliae | Salt | Pepper (Capsicum annuum) | [46] |
| Bacillus spp. | Salt | Pepper (Capsicum annuum) | [47] |
| Pseudomonas sp. and Hartmannibacter diazotrophicus | Salt | Alfalfa (Medicago sativa) | [48] |
| Pantoea agglomerans | Salt | Rice (Oryza sativa) | [49] |
| Arthrobacter aurescens, A. woluwensis, Microbacterium oxydans, Bacillus megaterium, and B. aryabhattai | Salt | Soybean (Glycine max) | [50] |
| Bacillus aryabhattai and B. mesonae | Salt | Tomato (Solanum lycopersicum) | [51] |
| Pseudomonas sp. | Salt | Arabidopsis thaliana | [52] |
| Pseudomonas fluorescens | Salt | Barley (Hordeum vulgare) | [53] |
| Arthrobacter nitroguajacolicus | Salt | Wheat (Triticum aestivum) | [54] |
| Bacillus cereus and B. aerius | Salt | Safflower (Carthamus tinctorius) | [55] |
| Pseudomonas and Azospirillum brasilense | Salt | Rapeseed (Brassica napus) | [56] |
| Pseudomonas geniculate | Salt | Maize (Zea mays) | [57] |
| Bacillus halotolerans and Lelliottia amnigena, | Salt | Wheat (Triticum aestivum) | [58] |
| Fungi | |||
| Glomus mosseae and Glomus intraradices | Drought | Rose geranium (Pelargonium graveolens L.) | [59] |
| Trichoderma atroviride strain (TaID20G) | Drought | Maize (Zea mays L.) | [60] |
| Gaeumannomyces cylindrosporus | Drought | Maize (Zea mays) | [61] |
| Arbuscular mycorhizal fungi (AMF) | Drought | Sweet potato (Ipomoea batatas (L.) Lam.) | [62] |
| AM fungus Funneliformis mosseae | Drought | Trifoliate orange [Poncirus trifoliata (L.) Raf.] | [63] |
| Trichoderma harzianum | Drought | Tomato (Solanum lycopersicum) | [64] |
| Rhizophagus intraradices, Funneliformis mosseae, F. geosporum | Drought | Wheat (Triticum aestivum | [65] |
| Arbuscular mycorrhizal fungi | Drought | Chinese lyme grass (Leymus chinensis) and limpograss (Hemarthria altissima) | [66] |
| Trichoderma harzinum 1, Trichoderma harzianum 2, Chaetomium globosum, and Talaromyces flavus | Drought | Rice (Oryza sativa L.) | [67] |
| Funneliformis mosseae, Glomus mosseae, G. intraradices, and G. etunicatum | Salt | Desert grass (Panicum turgidum) | [68] |
| Trichoderma harzianum | Salt | Indian mustard (Brassica juncea) | [69] |
| Trichoderma harzianum | Salt | Tomato (Solanum lycopersicum) | [70] |
| Trichoderma harzianum | Salt | Rice (Oryza sativa) and maize (Zea mays) | [71] |
| Klebsiella sp. | Salt | Oat (Avena sativa) | [72] |
| Glomus etunicatum, G. intraradices, and G. mosseae | Salt | Cucumber (Cucumis sativus) | [73] |
| Colobanthus quitensis and Deschampsia antarctica | Salt | Lettuce (Lactuca sativa) and tomato (Solanum lycopersicum) | [74] |
| Bacteria + Fungi | |||
| Bacillus thuringiensis + Arbuscular mycorrhizal fungus | Drought | French lavender (Lavandula dentata) | [75] |
| Pseudomonas putida + Rhizophagus irregularis | Drought | Calotrope (Calotropis procera Ait.) | [76] |
| Micrococcus yunnanensis + Claroideoglomus etunicatum | Drought | Moldavian balm (Dracocephalum moldavica L.) | [77] |
| Pseudomonas fluorescens + Rhizophagus irregularis or Funneliformis mosseae | Drought | Arizona cypress (Cupressus arizonica Green) | [78] |
| Pseudomonas fluorescence + Glomus mosseae | Salt | Bean (Phaseolus vulgaris) | [79] |
| Methylobacterium oryzae + Glomus etunicatum | Salt | Rice (Oryza sativa) | [80] |
| Bacillus subitilis + Glomus. etunicatum, G. intraradices, and G. mosseae | Salt | Acacia (Acacia gerrardii) | [81] |
| Bradyrhizobium sp. + Trichoderma asperelloides | Salt | Cowpea (Vigna unguiculate) | [82] |
| Microorganism | Heavy Metal | Reference |
|---|---|---|
| Bacteria | ||
| Azotobacter chroococum and Rhizobium leguminosarum | Pb | [152] |
| Pseudomonas sp. SRI2, Psychrobacter sp. SRS8, and Bacillus sp. SN9 | Ni | [153] |
| Sporosarcina ginsengisoli | As (III) | [154] |
| Bacillus cereus | Cr (VI) | [154] |
| P. macerans NBRFT5, B. endophyticus NBRFT4, B. pumilus NBRFT9 | Cu, Ni, and Zn | [155] |
| Bacillus thuringiensis GDB-1 | As | [156] |
| Bacillus cereus strain XMCr-6 | Cr (VI) | [157] |
| Bacillus subtilis | Cr (VI) | [158] |
| Pseudomonas putida | Cr (VI) | [158] |
| Pseudomonas sp. LK9 | Cd, Cu, and Zn | [159] |
| Enterobacter sp. And Klebsiella sp. | Cd, Pb, and Zn | [160] |
| Kocuria flava | Cu | [154] |
| Pseudomonas veronii | Cd, Cu, and Zn | [154] |
| Bacillus pumilus E2S2 and Bacillus sp. E1S2 | Cd and Zn | [161] |
| Enterobacter cloacae B2-DHA | Cr (VI) | [162] |
| Planomicrobium chinense, B. cereus, P. fluorescens | Co, Mn, Ni, and Pb | [163] |
| B. cereus, P. moraviensis | Mn and Cd | [164] |
| B. safensis FO-036b (T) and P. fluorescens | Pb and Zn | [165] |
| Fungi | ||
| Pleurotus platypus | Ag | [166] |
| Rhizopus oryzae (MPRO) | Cr (VI) | [167] |
| Aspergillus versicolor | Cu and Ni | [154] |
| Aspergillus fumigatus | Pb | [168] |
| Rhizopus oryzae | Cu | [169] |
| Algae | ||
| Spirogyra spp. and Cladophora spp. | Cu (II) and Pb (II) | [154] |
| Spirogyra spp. and Spirullina spp. | Cr Cu, Fe, Mn, and Zn | [154,170] |
| Cystoseira barbata | Cd, Ni, and Pb | [171] |
| Hydrodictylon, Oedogonium, and Rhizoclonium spp. | As | [172] |
| Microorganism | Stress | Plant Species | Reference |
|---|---|---|---|
| Bacteria | |||
| Azospirillum brasilense and Bacillus amyloliquefaciens | High temperature | Wheat (Triticum aestivum) | [175] |
| Bacillus amyloliquefaciens | High temperature | Rice (Oryza sativa) | [205] |
| Bacillus amyloliquefaciens | High temperature | Wheat (Triticum aestivum) | [208] |
| Pseudomonas syringae | High temperature | Arabidopsis thaliana | [209] |
| Enterobacter sp. | High temperature | Arabidopsis thaliana | [210] |
| Bacillus velezensis | High temperature | Wheat (Triticum aestivum) | [211] |
| Bacillus cereus | High temperature | Tomato (Solanum lycopersicum) | [212] |
| Bacillus cereus | High temperature | Tomato (Solanum lycopersicum) | [213] |
| Pseudomonas, Bacillus, Stenotrophomonas, Methylobacterium, Arthrobacter, Pantoea, Achromobacter, Acinetobacter, Exiguobacterium and Staphylococcus, Enterobacter, Providencia, Klebsiella and Leclercia, Brevundimonas, Flavobacterium, Kocuria, Kluyvera, and Planococcus | Cold temperature | Wheat (Triticum aestivum) | [214] |
| Arthrobacter, Flavimonas, Flavobacterium, Massilia, Pedobacter, and Pseudomonas | Cold temperature | Tomato (Solanum lycopersicum) | [215] |
| Rhizobacterial isolates of Bacillus genera, Gu2 and 127b | Cold temperature | Wheat (Triticum aestivum) | [216] |
| Pseudomonas fragi, P. chloropaphis, P. fluorescens, P. proteolytica, and Brevibacterium frigoritolerans | Cold temperature | Bean (Phaseolus vulgaris L.) | [217] |
| Bradyrhizobium japonicum | Flooding | Soybean (Glycine max) | [218] |
| Achromobacter xylosoxidans, Serratia ureilytica, Herbaspirillum seropedicae, and Ochrobactrum rhizosphaerae | Flooding | Tulsi (Ocimum sanctum) | [219] |
| Pseudomonas putida | Flooding | Cucumber (Cucumis sativus) | [220] |
| Azospira oryzae, Pelomonas saccharophila, and Methylosinus sp. | Flooding | Rice (Oryza sativa) | [221] |
| Pseudomonas putida | Flooding | Rumex palustris | [222] |
| Fungi | |||
| Glomus deserticola and Glomus constrictum | High temperature | Tomato (Solanum lycopersicum) | [223] |
| Aspergillus japonicas | High temperature | Soybean (Glycine max) and sunflower (Helianthus annuus) | [224] |
| Thermomyces sp. | High temperature | Cucumber (Cucumis sativus) | [225] |
| Thermomyces lanuginosus | High temperature | Cullen plicata | [226] |
| Glomus mosseae | Cold | Elymus nutans Griseb | [227] |
| Trichoderma harzianum | Cold | Tomato (Solanum lycopersicum L.) | [115] |
| Glomus versiforme and Rhizophagus irregularis | Cold | Barley (Hordeum vulgare L.) | [228] |
| Rhizophagus irregularis | Cold | Cucumber (Cucumis sativus L.) | [15] |
| Rhizophagus irregularis | Flooding | Tomato (Solanum lycopersicum) | [229] |
| Glomus intraradices, G. versiforme, and G. etunicatum | Flooding | Cattail (Typha orientalis) and rice (Oryza sativa) | [230] |
| Trichoderma | Flooding | Rice (Oryza sativa) | [231] |
| Aspergillus fumigatus | Flooding | Arabidopsis sp. | [232] |
| Bacteria and fungi | |||
| Bradyrhizobium + arbuscular mycorrhizal fungi | High temperature | Soybean (Glycine max L.) | [233] |
| Proteobacteria, Actinobacteria, Chloroflexi, and Nitrospirae + Dothideomycetes, Sordariomycetes, and Ascomycota | High temperature and drought | Sorghum (Sorghum bicolor L.) and foxtail millet (Setaria italica L.) | [234] |
| Bacillus and Pseudomonas + Penicillium | Cold temperature | Potato (Solanum tuberosum) | [235] |
| Paraburkholderia graminis C4D1M and Funneliformis mosseae | Cold temperature | Tomato (Solanum lycopersicum L.) | [236] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalal, A.; Oliveira, C.E.d.S.; Rosa, P.A.L.; Galindo, F.S.; Teixeira Filho, M.C.M. Beneficial Microorganisms Improve Agricultural Sustainability under Climatic Extremes. Life 2023, 13, 1102. https://doi.org/10.3390/life13051102
Jalal A, Oliveira CEdS, Rosa PAL, Galindo FS, Teixeira Filho MCM. Beneficial Microorganisms Improve Agricultural Sustainability under Climatic Extremes. Life. 2023; 13(5):1102. https://doi.org/10.3390/life13051102
Chicago/Turabian StyleJalal, Arshad, Carlos Eduardo da Silva Oliveira, Poliana Aparecida Leonel Rosa, Fernando Shintate Galindo, and Marcelo Carvalho Minhoto Teixeira Filho. 2023. "Beneficial Microorganisms Improve Agricultural Sustainability under Climatic Extremes" Life 13, no. 5: 1102. https://doi.org/10.3390/life13051102