
Identifying mRNAs Residing in Myelinating Oligodendrocyte Processes as a Basis for Understanding Internode Autonomy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Subcellular Fractionation
2.3. RT-qPCR Analysis
2.4. Analysis of MBRS Data
2.5. Analyses of Expression Patterns for mRNAs in AMBA and AMSCA Images
2.6. Statistical Analysis
3. Results
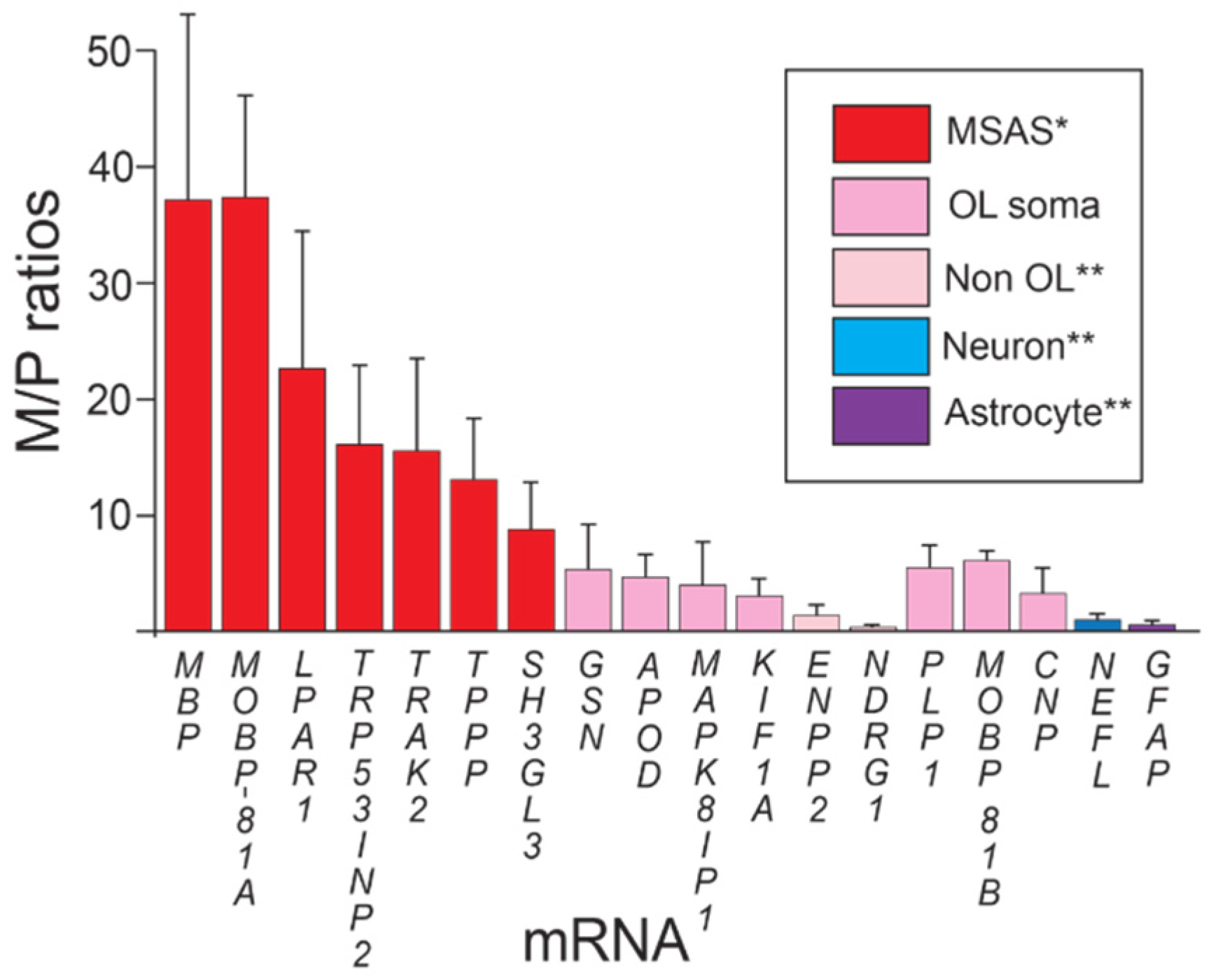
3.1. Enrichments of MSAS mRNAs in Rat Brain Myelin Based on RT-qPCR

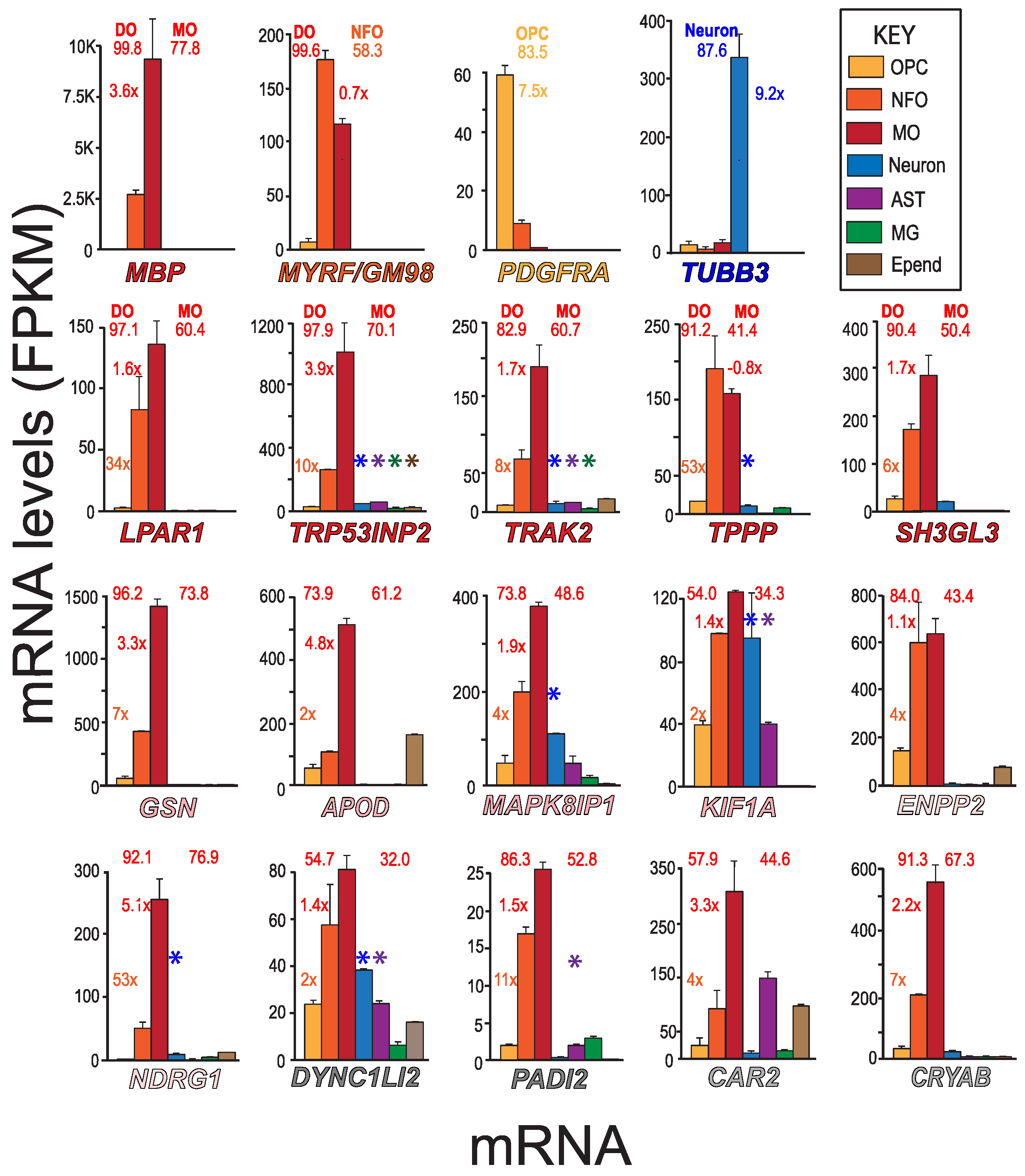
3.2. Identifying Neural Cell-Types Expressing MSAS mRNAs Using MBRS Data
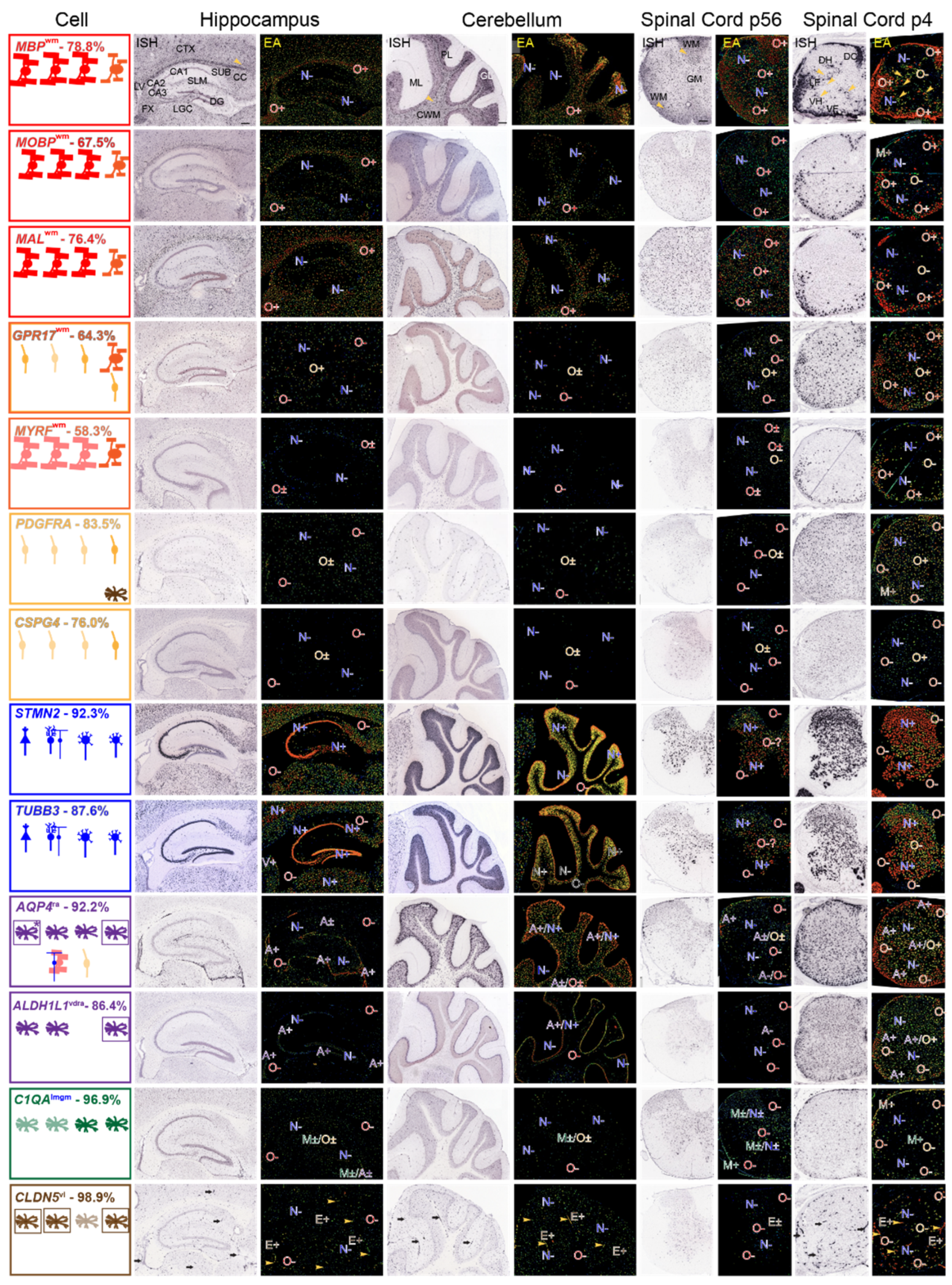
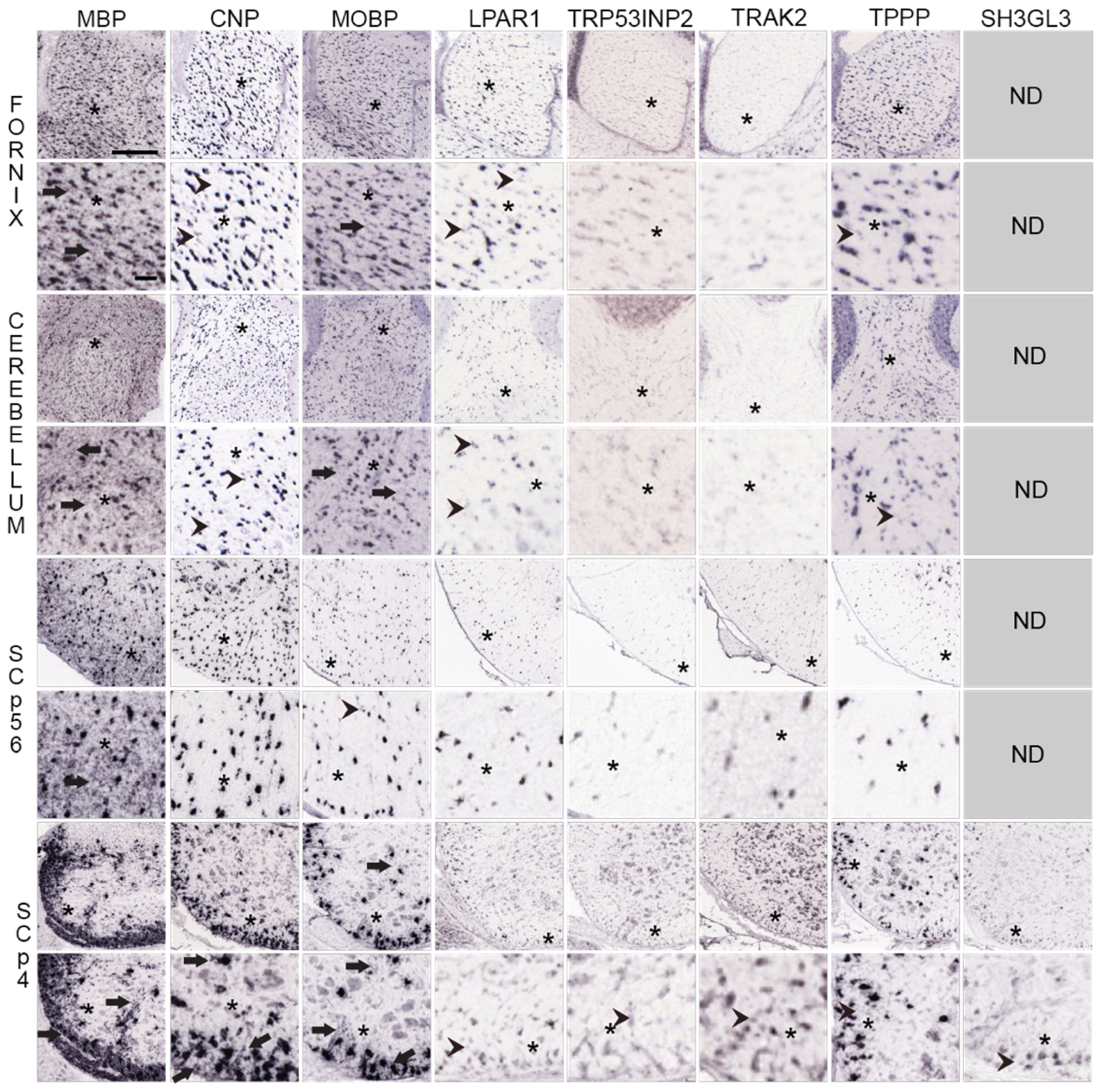
3.3. Recognizing Similarities and Differences among Neural Cell-Type Specific mRNA Expression Patterns in AMBA and AMSCA Images
3.3.1. Regions Occupied by Oligodendrocyte Lineage Cells—AMBA and AMSCA Data

 , Figure 3). In P4 spinal cord, MO mRNA-expressing NFOs are largely restricted to white-matter-rich ventral and lateral funiculi and dorsal columns (O+, Figure 3), as depicted in the leftmost panels (
, Figure 3). In P4 spinal cord, MO mRNA-expressing NFOs are largely restricted to white-matter-rich ventral and lateral funiculi and dorsal columns (O+, Figure 3), as depicted in the leftmost panels ( , Figure 3). Neither OPCs nor neurons in P4 spinal cord gray matter express these mRNAs, (N−, O−, Figure 3), as evidenced by comparisons with expression patterns of OPC and neuronal mRNAs (see below). For MBP, though not for MOBP or MAL, mRNAs, surrounding the few gray matter MO somata (red spots depicting high expression in the cells, EA images, Figure 3) are oligodendrocyte-processes/MSAS expressing MBP mRNA (green and yellow strands, EA images, arrowheads, Figure 3).). OPCs spread throughout P4 spinal cord gray matter express GPR17 mRNA (O−, Figure 3), whereas MOs in corpus callosum and fornix express MYRF mRNA (O+, Figure 3). Brain and spinal cord neurons express neither GPR17 nor MYRF mRNAs (N−, Figure 3).
, Figure 3). Neither OPCs nor neurons in P4 spinal cord gray matter express these mRNAs, (N−, O−, Figure 3), as evidenced by comparisons with expression patterns of OPC and neuronal mRNAs (see below). For MBP, though not for MOBP or MAL, mRNAs, surrounding the few gray matter MO somata (red spots depicting high expression in the cells, EA images, Figure 3) are oligodendrocyte-processes/MSAS expressing MBP mRNA (green and yellow strands, EA images, arrowheads, Figure 3).). OPCs spread throughout P4 spinal cord gray matter express GPR17 mRNA (O−, Figure 3), whereas MOs in corpus callosum and fornix express MYRF mRNA (O+, Figure 3). Brain and spinal cord neurons express neither GPR17 nor MYRF mRNAs (N−, Figure 3). ) and by adult OPCs distributed at low density [54,55] throughout P56 brain and spinal cord (O±, Figure 3, in leftmost panels,
) and by adult OPCs distributed at low density [54,55] throughout P56 brain and spinal cord (O±, Figure 3, in leftmost panels,  ). Neither MOs (O−, Figure 3), NFOs (O−, Figure 3) nor neurons (N−, Figure 3) express PDGFRA nor CSPG4 mRNAs.
). Neither MOs (O−, Figure 3), NFOs (O−, Figure 3) nor neurons (N−, Figure 3) express PDGFRA nor CSPG4 mRNAs.3.3.2. Regions Occupied by Non-Oligodendrocyte Lineage Cells—AMBA and AMSCA Data
 , Figure 3) and to neurons spread throughout the cortex, lateral geniculate complex and subiculum. In cerebellum, Purkinje (
, Figure 3) and to neurons spread throughout the cortex, lateral geniculate complex and subiculum. In cerebellum, Purkinje ( ) and granule cell (
) and granule cell ( ) neurons express both STMN2 and TUBB3 mRNAs (N+, Figure 3), though neurons in the molecular layer do not (N−, Figure 3). In P56 and P4 spinal cord, neurons expressing STMN2 and TUBB3 mRNAs (N+, Figure 3) are distributed throughout gray matter, including in large profile ventral horn motoneurons (
) neurons express both STMN2 and TUBB3 mRNAs (N+, Figure 3), though neurons in the molecular layer do not (N−, Figure 3). In P56 and P4 spinal cord, neurons expressing STMN2 and TUBB3 mRNAs (N+, Figure 3) are distributed throughout gray matter, including in large profile ventral horn motoneurons ( , Figure 3, leftmost panel). Importantly, neither STMN2 or TUBB3 mRNAs are expressed in P56 mouse brain and spinal cord white matter where MOs reside (O−, Figure 3), nor in P4 spinal cord white matter (O−, Figure 3) where NFOs reside, allowing clear distinctions between mRNAs expressed by MOs and NFOs, from mRNAs expressed by neurons.
, Figure 3, leftmost panel). Importantly, neither STMN2 or TUBB3 mRNAs are expressed in P56 mouse brain and spinal cord white matter where MOs reside (O−, Figure 3), nor in P4 spinal cord white matter (O−, Figure 3) where NFOs reside, allowing clear distinctions between mRNAs expressed by MOs and NFOs, from mRNAs expressed by neurons.3.4. Regions in AMBA and AMSCA Images Where MSAS mRNAs Are Expressed
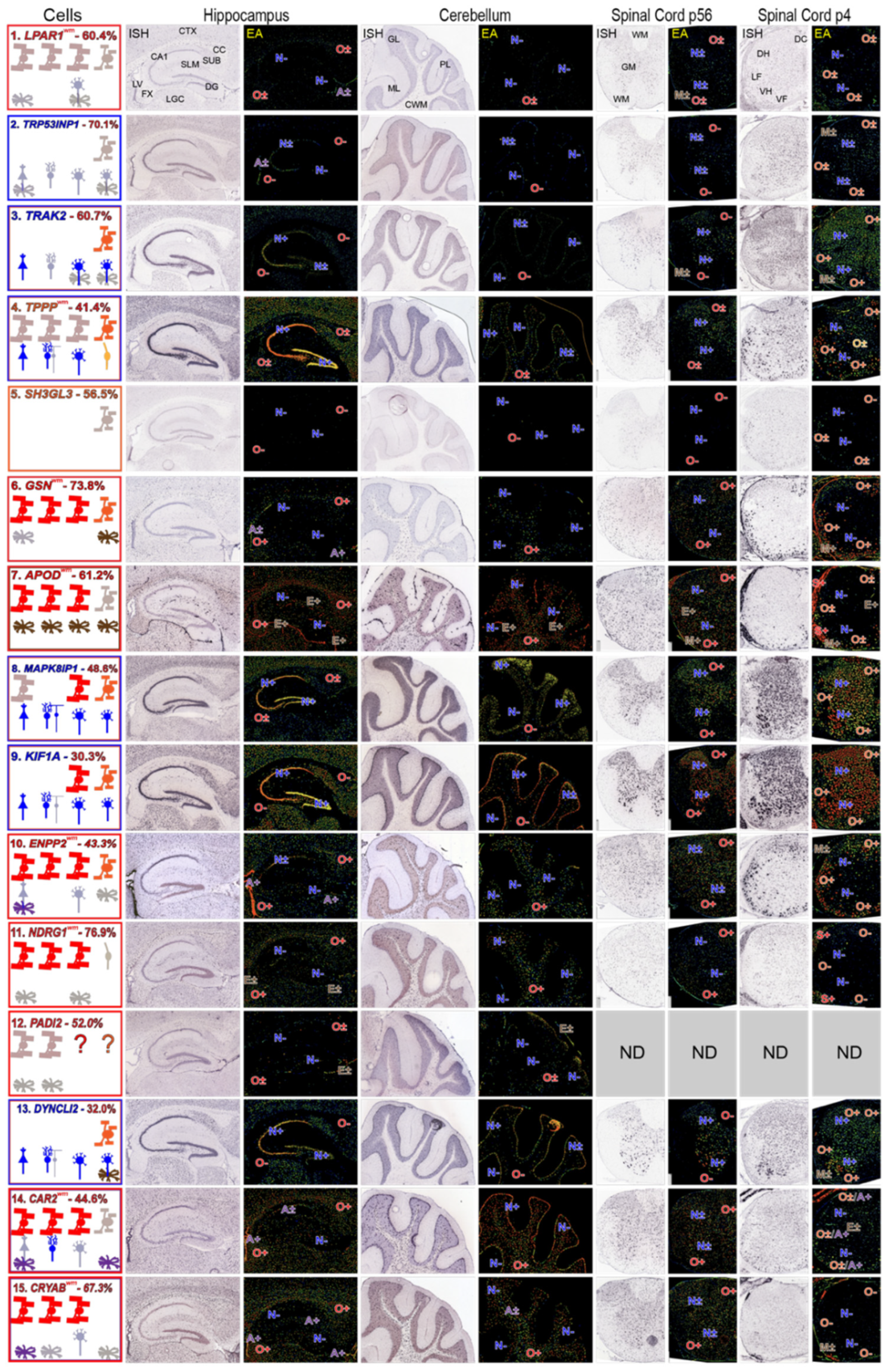
3.5. Using AMBA and AMSCA Images as an Attempt to Identify MSAS mRNAs
4. Discussion
4.1. Effectiveness of the Subcellular Fractionation Approach in Identifying MSAS mRNAs
4.2. An Independent Approach to Identify MSAS mRNAs
4.3. Roles Which Identified MSAS mRNAs and Encoded Proteins Might Play in Internodal Myelination
4.4. Future Efforts to Identify MSAS mRNAs
4.4.1. Locating Intracellular Sites Where MSAS mRNAs Reside Using ISH
4.4.2. Differential Gene Expression Analysis
4.4.3. Final Thoughts
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Das, S.; Vera, M.; Gandin, V.; Singer, R.H.; Tutucci, E. Intracellular mRNA transport and localized translation. Nat. Rev. Mol. Cell Biol. 2021, 22, 483–504. [Google Scholar] [CrossRef] [PubMed]
- Biever, A.; Donlin-Asp, P.G.; Schuman, E.M. Local translation in neuronal processes. Curr. Opin. Neurobiol. 2019, 57, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Cioni, J.M.; Koppers, M.; Holt, C.E. Molecular control of local translation in axon development and maintenance. Curr. Opin. Neurobiol. 2018, 51, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Turner-Bridger, B.; Caterino, C.; Cioni, J.M. Molecular mechanisms behind mRNA localization in axons. Open Biol. 2020, 10, 200177. [Google Scholar] [CrossRef] [PubMed]
- Brophy, P.J.; Boccaccio, G.L.; Colman, D.R. The distribution of myelin basic protein mRNAs within myelinating oligodendrocytes. Trends Neurol. Sci. 1993, 16, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Smith, R. Moving Molecules: mRNA Trafficking in Mammalian Oligodendrocytes and Neurons. Neuroscientist 2004, 10, 495–500. [Google Scholar] [CrossRef]
- White, R.; Kramer-Albers, E.M. Axon-glia interaction and membrane traffic in myelin formation. Front. Cell. Neurosci. 2014, 7, 284. [Google Scholar] [CrossRef]
- Carson, J.H.; Gao, Y.; Tatavarty, V.; Levin, M.K.; Korza, G.; Francone, V.P.; Kosturko, L.D.; Maggipinto, M.J.; Barbarese, E. Multiplexed RNA trafficking in oligodendrocytes and neurons. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2008, 1779, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Torvund-Jensen, J.; Steengaard, J.; Reimer, L.; Fihl, L.B.; Laursen, L.S. Transport and translation of MBP mRNA is regulated differently by distinct hnRNP proteins. J. Cell Sci. 2014, 127, 1550–1564. [Google Scholar] [CrossRef] [Green Version]
- Yergert, K.M.; Doll, C.A.; O’Rouke, R.; Hines, J.H.; Appel, B. Identification of 3’ UTR motifs required for mRNA localization to myelin sheaths in vivo. PLoS Biol. 2021, 19, e3001053. [Google Scholar] [CrossRef]
- Meservey, L.M.; Topkar, V.V.; Fu, M.M. mRNA Transport and Local Translation in Glia. Trends Cell Biol. 2021, 31, 419–423. [Google Scholar] [CrossRef]
- Holz, A.; Schaeren-Wiemers, N.; Schaefer, C.; Pott, U.; Colello, R.J.; Schwab, M.E. Molecular and developmental characterization of novel cDNAs of the myelin-associated oligodendrocytic basic protein. J. Neurosci. 1996, 16, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Mizuno, R.; Nishimura, T.; Ogawa, Y.; Yoshikawa, H.; Fujimura, H.; Adachi, E.; Kishimoto, T.; Yanagihara, T.; Sakoda, S. Cloning and expression of myelin-associated oligodendrocytic basic protein. A novel basic protein constituting the central nervous system myelin. J. Biol. Chem. 1994, 269, 31725–31730. [Google Scholar] [CrossRef]
- LoPresti, P.; Szuchet, S.; Papasozomenos, S.C.; Zinkowski, R.P.; Binder, L.I. Functional implications for the microtubule-associated protein tau: Localization in oligodendrocytes. Proc. Natl. Acad. Sci. USA 1995, 92, 10369–10373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghandour, M.S.; Skoff, R.P. Double-labeling in situ hybridization analysis of mRNAs for carbonic anhydrase II and myelin basic protein: Expression in developing cultured glial cells. Glia 1991, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Quraishe, S.; Wyttenbach, A.; Matinyarare, N.; Perry, V.H.; Fern, R.; O’Connor, V. Selective and compartmentalized myelin expression of HspB5. Neuroscience 2016, 316, 130–142. [Google Scholar] [CrossRef]
- Gould, R.; Freund, C.; Palmer, F.; Knapp, P.E.; Huang, J.; Morrison, H.; Feinstein, D.L. Messenger RNAs for kinesin and dynein are located in neural processes. Biol. Bull. 1999, 197, 259–260. [Google Scholar] [CrossRef] [Green Version]
- Gould, R.M.; Freund, C.M.; Palmer, F.; Feinstein, D.L. Messenger RNAs located at sites of myelin assembly. J. Neurochem. 2000, 75, 1834–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, R.M.; Freund, C.M. mRNAs located at sites of myelin sheath assembly. Mol. Biol. Cell 2000, 11, 154a. [Google Scholar]
- Thakurela, S.; Garding, A.; Jung, R.B.; Muller, C.; Goebbels, S.; White, R.; Werner, H.B.; Tiwari, V.K. The transcriptome of mouse central nervous system myelin. Sci. Rep. 2016, 6, 25828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeren-Wiemers, N.; Schaefer, C.; Valenzuela, D.M.; Yancoupoulos, G.D.; Schwab, M.E. Identification of new oligodendrocyte- and myelin-specific genes by a differential screening approach. J. Neurochem. 1995, 65, 10–22. [Google Scholar] [CrossRef]
- Boyles, J.K.; Notterpek, L.M.; Wardell, M.R.; Rall, S.C., Jr. Identification, characterization, and tissue distribution of apolipoprotein D in the rat. J. Lipid Res. 1990, 31, 2243–2256. [Google Scholar] [CrossRef] [PubMed]
- Fuss, B.; Baba, H.; Phan, T.; Tuohy, V.K.; Macklin, W.B. Phosphodiesterase I, a novel adhesion molecule and/or cytokine involved in oligodendrocyte function. J. Neurosci. 1997, 17, 9095–9103. [Google Scholar] [CrossRef] [Green Version]
- Fox, M.A.; Colello, R.J.; Macklin, W.B.; Fuss, B. Phosphodiesterase-I alpha/autotaxin: A counteradhesive protein expressed by oligodendrocytes during onset of myelination. Mol. Cell. Neurosci. 2003, 23, 507–519. [Google Scholar] [CrossRef]
- Tanaka, J.; Sobue, K. Localization and characterization of gelsolin in nervous tissues: Gelsolin is specifically enriched in myelin-forming cells. J. Neurosci. 1994, 14, 1038–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrand, C.; Ferraz, C.; Clavel, M.C.; Rabie, A. Immunocytochemical localization of gelsolin in oligodendroglia of the developing rabbit central nervous system. Dev. Brain Res. 1986, 30, 231–235. [Google Scholar] [CrossRef]
- Stankoff, B.; Barron, S.; Allard, J.; Barbin, G.; Noel, F.; Aigrot, M.S.; Premont, J.; Sokoloff, P.; Zalc, B.; Lubetzki, C. Oligodendroglial expression of Edg-2 receptor: Developmental analysis and pharmacological responses to lysophosphatidic acid. Mol. Cell. Neurosci. 2002, 20, 415–428. [Google Scholar] [CrossRef]
- King, R.H.; Chandler, D.; Lopaticki, S.; Huang, D.; Blake, J.; Muddle, J.R.; Kilpatrick, T.; Nourallah, M.; Miyata, T.; Okuda, T.; et al. Ndrg1 in development and maintenance of the myelin sheath. Neurobiol. Dis. 2011, 42, 368–380. [Google Scholar] [CrossRef]
- Okuda, T.; Kokame, K.; Miyata, T. Differential expression patterns of NDRG family proteins in the central nervous system. J. Histochem. Cytochem. 2008, 56, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marechal, D.; Dansu, D.K.; Castro, K.; Patzig, J.; Magri, L.; Inbar, B.; Gacias, M.; Moyon, S.; Casaccia, P. N-myc downstream regulated family member 1 (NDRG1) is enriched in myelinating oligodendrocytes and impacts myelin degradation in response to demyelination. Glia 2022, 70, 321–336. [Google Scholar] [CrossRef]
- Mustonen, V.; Muruganandam, G.; Loris, R.; Kursula, P.; Ruskamo, S. Crystal and solution structure of NDRG1, a membrane-binding protein linked to myelination and tumour suppression. FEBS J. 2021, 288, 3507–3529. [Google Scholar] [CrossRef]
- Boggs, J.M.; Rangaraj, G.; Koshy, K.M.; Ackerley, C.; Wood, D.D.; Moscarello, M.A. Highly deiminated isoform of myelin basic protein from multiple sclerosis brain causes fragmentation of lipid vesicles. J. Neurosci. Res. 1999, 57, 529–535. [Google Scholar] [CrossRef]
- Finch, P.R.; Wood, D.D.; Moscarello, M.A. The presence of citrulline in a myelin protein fraction. FEBS Lett. 1971, 15, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musse, A.A.; Li, Z.; Ackerley, C.A.; Bienzle, D.; Lei, H.; Poma, R.; Harauz, G.; Moscarello, M.A.; Mastronardi, F.G. Peptidylarginine deiminase 2 (PAD2) overexpression in transgenic mice leads to myelin loss in the central nervous system. Dis. Model. Mech. 2008, 1, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Pritzker, L.B.; Joshi, S.; Gowan, J.J.; Harauz, G.; Moscarello, M.A. Deimination of myelin basic protein. 1. Effect of deimination of arginyl residues of myelin basic protein on its structure and susceptibility to digestion by cathepsin D. Biochemistry 2000, 39, 5374–5381. [Google Scholar] [CrossRef]
- Pritzker, L.B.; Nguyen, T.A.; Moscarello, M.A. The developmental expression and activity of peptidylarginine deiminase in the mouse. Neurosci. Lett. 1999, 266, 161–164. [Google Scholar] [CrossRef]
- Takahashi, M.; Tomizawa, K.; Fujita, S.C.; Sato, K.; Uchida, T.; Imahori, K. A brain-specific protein p25 is localized and associated with oligodendrocytes, neuropil, and fiber-like structures of the CA3 hippocampal region in the rat brain. J. Neurochem. 1993, 60, 228–235. [Google Scholar] [CrossRef]
- Tirian, L.; Hlavanda, E.; Olah, J.; Horvath, I.; Orosz, F.; Szabo, B.; Kovacs, J.; Szabad, J.; Ovadi, J. TPPP/p25 promotes tubulin assemblies and blocks mitotic spindle formation. Proc. Natl. Acad. Sci. USA 2003, 100, 13976–13981. [Google Scholar] [CrossRef] [Green Version]
- Fu, M.M.; McAlear, T.S.; Nguyen, H.; Oses-Prieto, J.A.; Valenzuela, A.; Shi, R.D.; Perrino, J.J.; Huang, T.T.; Burlingame, A.L.; Bechstedt, S.; et al. The Golgi Outpost Protein TPPP Nucleates Microtubules and Is Critical for Myelination. Cell 2019, 179, 132–146.e14. [Google Scholar] [CrossRef]
- Ghandour, M.S.; Langley, O.K.; Vincendon, G.; Gombos, G.; Filippi, D.; Limozin, N.; Dalmasso, D.; Laurent, G. Immunochemical and immunohistochemical study of carbonic anhydrase II in adult rat cerebellum: A marker for oligodendrocytes. Neuroscience 1980, 5, 559–571. [Google Scholar] [CrossRef]
- Iwaki, T.; Wisniewski, T.; Iwaki, A.; Corbin, E.; Tomokane, N.; Tateishi, J.; Goldman, J.E. Accumulation of alpha B-crystallin in central nervous system glia and neurons in pathologic conditions. Am. J. Pathol. 1992, 140, 345–356. [Google Scholar] [PubMed]
- Boggs, J.M. Myelin basic protein: A multifunctional protein. Cell. Mol. Life Sci. CMLS 2006, 63, 1945–1961. [Google Scholar] [CrossRef] [PubMed]
- Montague, P.; Dickinson, P.J.; McCallion, A.S.; Stewart, G.J.; Savioz, A.; Davies, R.W.; Kennedy, P.G.E.; Griffiths, I.R. Developmental expression of the murine Mobp gene. J. Neurosci. Res. 1997, 49, 133–143. [Google Scholar] [CrossRef]
- Gould, R.M.; Freund, C.M.; Engler, J.; Morrison, H.G. Optimization of homogenization conditions used to isolate mRNAs in processes of myelinating oligodendrocytes. Biol. Bull. 2000, 199, 215–217. [Google Scholar] [CrossRef]
- Gould, R.M.; Freund, C.; Barbarese, E. Myelin-associated oligodendrocytic basic protein (MOBP) mRNAs reside at different subcellular locations. J. Neurochem. 1999, 73, 1913–1924. [Google Scholar]
- Trapp, B.D.; Bernier, L.; Andrews, S.B.; Colman, D.R. Cellular and subcellular distribution of 2′,3′-cyclic nucleotide 3′-phosphodiesterase and its mRNA in the rat central nervous system. J. Neurochem. 1988, 51, 859–868. [Google Scholar] [CrossRef]
- Bloom, G.S. The UNC-104/KIF1 family of kinesins. Curr. Opin. Cell Biol. 2001, 13, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, H.; Sekine, Y.; Takemura, R.; Nangaku, M.; Hirokawa, N. Kinesin family in murine central nervous system. J. Cell Biol. 1992, 119, 1287–1296. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-Sequencing Transcriptome and Splicing Database of Glia, Neurons, and Vascular Cells of the Cerebral Cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, A.; Munoz-Manchado, A.B.; Codeluppi, S.; Lonnerberg, P.; La Manno, G.; Jureus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Brain structure. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef]
- Kramer, E.M.; Schardt, A.; Nave, K.A. Membrane traffic in myelinating oligodendrocytes. Microsc. Res. Tech. 2001, 52, 656–671. [Google Scholar] [CrossRef]
- Pringle, N.P.; Richardson, W.D. A singularity of PDGF alpha-receptor expression in the dorsoventral axis of the neural tube may define the origin of the oligodendrocyte lineage. Development 1993, 117, 525–533. [Google Scholar] [CrossRef]
- Stallcup, W.B. The NG2 proteoglycan: Past insights and future prospects. J. Neurocytol. 2002, 31, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Moyon, S.; Dubessy, A.L.; Aigrot, M.S.; Trotter, M.; Huang, J.K.; Dauphinot, L.; Potier, M.C.; Kerninon, C.; Melik Parsadaniantz, S.; Franklin, R.J.; et al. Demyelination causes adult CNS progenitors to revert to an immature state and express immune cues that support their migration. J. Neurosci. 2015, 35, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, M.R.; Polito, A.; Levine, J.M.; Reynolds, R. NG2-expressing glial progenitor cells: An abundant and widespread population of cycling cells in the adult rat CNS. Mol. Cell. Neurosci. 2003, 24, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.; Caceres, A. Expression of the class III beta-tubulin isotype in developing neurons in culture. J. Neurosci. Res. 1992, 32, 516–529. [Google Scholar] [CrossRef]
- Jiang, Y.Q.; Oblinger, M.M. Differential regulation of beta III and other tubulin genes during peripheral and central neuron development. J. Cell Sci. 1992, 103 Pt. 3, 643–651. [Google Scholar] [CrossRef]
- Mori, N.; Morii, H. SCG10-related neuronal growth-associated proteins in neural development, plasticity, degeneration, and aging. J. Neurosci. Res. 2002, 70, 264–273. [Google Scholar] [CrossRef]
- Di Paolo, G.; Pellier, V.; Catsicas, M.; Antonsson, B.; Catsicas, S.; Grenningloh, G. The phosphoprotein stathmin is essential for nerve growth factor-stimulated differentiation. J. Cell Biol. 1996, 133, 1383–1390. [Google Scholar] [CrossRef]
- Nielsen, S.; Nagelhus, E.A.; Amiry-Moghaddam, M.; Bourque, C.; Agre, P.; Ottersen, O.P. Specialized membrane domains for water transport in glial cells: High-resolution immunogold cytochemistry of aquaporin-4 in rat brain. J. Neurosci. 1997, 17, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, M.I.; Chu, S.H.; Hernandez, M.X.; Fang, M.J.; Modarresi, L.; Selvan, P.; MacGregor, G.R.; Tenner, A.J. Cell-specific deletion of C1qa identifies microglia as the dominant source of C1q in mouse brain. J. Neuroinflamm. 2017, 14, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef]
- Nico, B.; Ribatti, D.; Frigeri, A.; Nicchia, G.P.; Corsi, P.; Svelto, M.; Roncali, L. Aquaporin-4 expression during development of the cerebellum. Cerebellum 2002, 1, 207–212. [Google Scholar] [CrossRef]
- Stevens, B.; Allen, N.J.; Vazquez, L.E.; Howell, G.R.; Christopherson, K.S.; Nouri, N.; Micheva, K.D.; Mehalow, A.K.; Huberman, A.D.; Stafford, B.; et al. The classical complement cascade mediates CNS synapse elimination. Cell 2007, 131, 1164–1178. [Google Scholar] [CrossRef] [Green Version]
- Morita, K.; Sasaki, H.; Furuse, M.; Tsukita, S. Endothelial claudin: Claudin-5/TMVCF constitutes tight junction strands in endothelial cells. J. Cell Biol. 1999, 147, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lein, E.S.; Hawrylycz, M.J.; Ao, N.; Ayres, M.; Bensinger, A.; Bernard, A.; Boe, A.F.; Boguski, M.S.; Brockway, K.S.; Byrnes, E.J.; et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 2007, 445, 168–176. [Google Scholar] [CrossRef]
- Landry, C.F.; Watson, J.B.; Kashima, T.; Campagnoni, A.T. Cellular influences on RNA sorting in neurons and glia: An in situ hybridization histochemical study. Mol. Brain Res. 1994, 27, 1–11. [Google Scholar] [CrossRef]
- Chong, S.Y.; Rosenberg, S.S.; Fancy, S.P.; Zhao, C.; Shen, Y.A.; Hahn, A.T.; McGee, A.W.; Xu, X.; Zheng, B.; Zhang, L.I.; et al. Neurite outgrowth inhibitor Nogo-A establishes spatial segregation and extent of oligodendrocyte myelination. Proc. Natl. Acad. Sci. USA 2012, 109, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- Baumann, N.; Pham-Dinh, D. Biology of Oligodendrocyte and Myelin in the Mammalian Central Nervous System. Physiol. Rev. 2001, 81, 871–927. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.E.; Martin, K.C.; Schuman, E.M. Local translation in neurons: Visualization and function. Nat. Struct. Mol. Biol. 2019, 26, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Colman, D.R.; Kreibich, G.; Frey, A.B.; Sabatini, D.D. Synthesis and incorporation of myelin polypeptides into CNS myelin. J. Cell Biol. 1982, 95, 598–608. [Google Scholar] [CrossRef]
- Tushev, G.; Glock, C.; Heumuller, M.; Biever, A.; Jovanovic, M.; Schuman, E.M. Alternative 3′ UTRs Modify the Localization, Regulatory Potential, Stability, and Plasticity of mRNAs in Neuronal Compartments. Neuron 2018, 98, 495–511.e6. [Google Scholar] [CrossRef] [Green Version]
- LoPresti, P. Regulation and differential expression of tau mRNA isoforms as oligodendrocytes mature in vivo: Implications for myelination. Glia 2002, 37, 250–257. [Google Scholar] [CrossRef]
- Pfeiffer, S.E.; Warrington, A.E.; Bansal, R. The oligodendrocyte and its many cellular processes. Trends Cell Biol. 1993, 3, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Waehneldt, T.V. Phylogeny of myelin proteins. Ann. N. Y. Acad.Sci. 1990, 605, 15–28. [Google Scholar] [CrossRef]
- Gould, R. Neuron Glia Biology. Commentary. Neuron Glia Biol. 2008, 4, 57–58. [Google Scholar]
- Gould, R.M.; Dawson, R.M.C. Incorporation of newly formed lecithin into peripheral nerve myelin. J. Cell Biol. 1976, 68, 480–496. [Google Scholar] [CrossRef] [PubMed]
- Gould, R.M.; Sinatra, R.S. Internodal distribution of phosphatidylcholine biosynthetic activity in teased peripheral nerve fibers: An autoradiographic study. J. Neurocytol. 1981, 10, 161–167. [Google Scholar] [CrossRef]
- Gould, R.M.; Connell, F.; Spivack, W. Phospholipid metabolism in mouse sciatic nerve in vivo. J. Neurochem. 1987, 48, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Gould, R.M.; Mattingly, G. Regional localization of RNA, protein and lipid metabolism in Schwann cells in vivo. J. Neurocytol. 1990, 19, 285–301. [Google Scholar] [CrossRef]
- Mahalingam, M. Laser Capture Microdissection: Insights into Methods and Applications. Methods Mol. Biol. 2018, 1723, 1–17. [Google Scholar] [CrossRef]
- Datta, S.; Malhotra, L.; Dickerson, R.; Chaffee, S.; Sen, C.K.; Roy, S. Laser capture microdissection: Big data from small samples. Histol. Histopathol. 2015, 30, 1255–1269. [Google Scholar] [CrossRef]
- Lecuyer, E.; Yoshida, H.; Parthasarathy, N.; Alm, C.; Babak, T.; Cerovina, T.; Hughes, T.R.; Tomancak, P.; Krause, H.M. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell 2007, 131, 174–187. [Google Scholar] [CrossRef] [Green Version]
- Cajigas, I.J.; Tushev, G.; Will, T.J.; tom Dieck, S.; Fuerst, N.; Schuman, E.M. The local transcriptome in the synaptic neuropil revealed by deep sequencing and high-resolution imaging. Neuron 2012, 74, 453–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumy, L.F.; Yeo, G.S.; Tung, Y.C.; Zivraj, K.H.; Willis, D.; Coppola, G.; Lam, B.Y.; Twiss, J.L.; Holt, C.E.; Fawcett, J.W. Transcriptome analysis of embryonic and adult sensory axons reveals changes in mRNA repertoire localization. RNA 2011, 17, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Mazare, N.; Oudart, M.; Cohen-Salmon, M. Local translation in perisynaptic and perivascular astrocytic processes—A means to ensure astrocyte molecular and functional polarity? J. Cell Sci. 2021, 134, jcs251629. [Google Scholar] [CrossRef]
- D’Arcy, B.R.; Silver, D.L. Local gene regulation in radial glia: Lessons from across the nervous system. Traffic 2020, 21, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Harauz, G.; Boggs, J.M. Myelin management by the 18.5-kDa and 21.5-kDa classic myelin basic protein isoforms. J. Neurochem. 2013, 125, 334–361. [Google Scholar] [CrossRef]
- Raasakka, A.; Ruskamo, S.; Kowal, J.; Barker, R.; Baumann, A.; Martel, A.; Tuusa, J.; Myllykoski, M.; Burck, J.; Ulrich, A.S.; et al. Membrane Association Landscape of Myelin Basic Protein Portrays Formation of the Myelin Major Dense Line. Sci. Rep. 2017, 7, 4974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, R.M.; Oakley, T.; Goldstone, J.V.; Dugas, J.C.; Brady, S.T.; Gow, A. Myelin sheaths are formed with proteins that originated in vertebrate lineages. Neuron Glia Biol. 2008, 4, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Lambert, J.D. Clade-specific genes and the evolutionary origin of novelty; new tools in the toolkit. Semin. Cell Dev. Biol. 2022; in press. [Google Scholar] [CrossRef] [PubMed]
- Pahl, N.; Homsi, S.; Morrison, H.G.; Gould, R.M. mRNAs located in Squalus acanthias (spiny dogfish) oligodendrocyte processes. Biol. Bull. 2002, 203, 217–218. [Google Scholar] [CrossRef]
- Raasakka, A.; Kursula, P. Flexible Players within the Sheaths: The Intrinsically Disordered Proteins of Myelin in Health and Disease. Cells 2020, 9, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moscarello, M.A.; Pritzker, L.; Mastronardi, F.G.; Wood, D.D. Peptidylarginine deiminase: A candidate factor in demyelinating disease. J. Neurochem. 2002, 81, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcao, A.M.; Meijer, M.; Scaglione, A.; Rinwa, P.; Agirre, E.; Liang, J.; Larsen, S.C.; Heskol, A.; Frawley, R.; Klingener, M.; et al. PAD2-Mediated Citrullination Contributes to Efficient Oligodendrocyte Differentiation and Myelination. Cell Rep. 2019, 27, 1090–1102.e10. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, S.; Cantuti Castelvetri, L.; Simons, M. Metabolism and functions of lipids in myelin. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 999–1005. [Google Scholar] [CrossRef]
- Vassall, K.A.; Bamm, V.V.; Harauz, G. MyelStones: The executive roles of myelin basic protein in myelin assembly and destabilization in multiple sclerosis. Biochem. J. 2015, 472, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.V.; Rakitina, T.V.; Ziganshin, R.H.; Arapidi, G.P.; Saratov, G.A.; Kudriaeva, A.A.; Belogurov, A.A. Comprehensive Atlas of the Myelin Basic Protein Interaction Landscape. Biomolecules 2021, 11, 1628. [Google Scholar] [CrossRef] [PubMed]
- Larijani, B.; Pytowski, L.; Vaux, D.J. The enigma of phosphoinositides and their derivatives: Their role in regulation of subcellular compartment morphology. Biochim. Biophys. Acta (BBA)-Biomembr. 2022, 1864, 183780. [Google Scholar] [CrossRef]
- Bakhti, M.; Aggarwal, S.; Simons, M. Myelin architecture: Zippering membranes tightly together. Cell. Mol. Life Sci. 2014, 71, 1265–1277. [Google Scholar] [CrossRef]
- Belgrad, J.; De Pace, R.; Fields, R.D. Autophagy in Myelinating Glia. J. Neurosci. 2020, 40, 256–266. [Google Scholar] [CrossRef]
- Baron, W.; Hoekstra, D. On the biogenesis of myelin membranes: Sorting, trafficking and cell polarity. FEBS Lett. 2010, 584, 1760–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterstein, C.; Trotter, J.; Kramer-Albers, E.M. Distinct endocytic recycling of myelin proteins promotes oligodendroglial membrane remodeling. J. Cell Sci. 2008, 121, 834–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucrot, E.; Ferreira, A.P.; Almeida-Souza, L.; Debard, S.; Vallis, Y.; Howard, G.; Bertot, L.; Sauvonnet, N.; McMahon, H.T. Endophilin marks and controls a clathrin-independent endocytic pathway. Nature 2015, 517, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Kjaerulff, O.; Brodin, L.; Jung, A. The structure and function of endophilin proteins. Cell Biochem. Biophys. 2011, 60, 137–154. [Google Scholar] [CrossRef]
- Nowak, J.; Archange, C.; Tardivel-Lacombe, J.; Pontarotti, P.; Pebusque, M.J.; Vaccaro, M.I.; Velasco, G.; Dagorn, J.C.; Iovanna, J.L. The TP53INP2 protein is required for autophagy in mammalian cells. Mol. Biol. Cell 2009, 20, 870–881. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, J.C.; Dantas, T.J.; Vallee, R.B. Distinct roles for dynein light intermediate chains in neurogenesis, migration, and terminal somal translocation. J. Cell Biol. 2019, 218, 808–819. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Kumar, C.; Wasnik, N.; Mylavarapu, S.V.S. Dynein light intermediate chains as pivotal determinants of dynein multifunctionality. J. Cell Sci. 2021, 134, jcs254870. [Google Scholar] [CrossRef]
- Okada, Y.; Yamazaki, Y.; Sekine-Aizawa, Y.; Hirokawa, N. The neuron-specific kinesin superfamily protein KIF1A is a unique monomeric motor for anterograde axonal transport of synaptic vesicle precursors. Cell 1995, 81, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Koushika, S.P. “JIP”ing along the axon: The complex roles of JIPs in axonal transport. BioEssays 2008, 30, 10–14. [Google Scholar] [CrossRef]
- Verhey, K.J.; Meyer, D.; Deehan, R.; Blenis, J.; Schnapp, B.J.; Rapoport, T.A.; Margolis, B. Cargo of kinesin identified as JIP scaffolding proteins and associated signaling molecules. J. Cell Biol. 2001, 152, 959–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brickley, K.; Stephenson, F.A. Trafficking kinesin protein (TRAK)-mediated transport of mitochondria in axons of hippocampal neurons. J. Biol. Chem. 2011, 286, 18079–18092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenton, A.R.; Jongens, T.A.; Holzbaur, E.L.F. Mitochondrial adaptor TRAK2 activates and functionally links opposing kinesin and dynein motors. Nat. Commun. 2021, 12, 4578. [Google Scholar] [CrossRef]
- Meyer, N.; Rinholm, J.E. Mitochondria in Myelinating Oligodendrocytes: Slow and Out of Breath? Metabolites 2021, 11, 359. [Google Scholar] [CrossRef] [PubMed]
- Rinholm, J.E.; Vervaeke, K.; Tadross, M.R.; Tkachuk, A.N.; Kopek, B.G.; Brown, T.A.; Bergersen, L.H.; Clayton, D.A. Movement and structure of mitochondria in oligodendrocytes and their myelin sheaths. Glia 2016, 64, 810–825. [Google Scholar] [CrossRef]
- Nakamura, D.S.; Kennedy, T.E. Netrin-1 marshals mitochondrial movement, morphology, and metabolism in myelin. Neural Regen. Res. 2021, 16, 2399–2400. [Google Scholar] [CrossRef]
- Battefeld, A.; Popovic, M.A.; de Vries, S.I.; Kole, M.H.P. High-Frequency Microdomain Ca2+ Transients and Waves during Early Myelin Internode Remodeling. Cell Rep. 2019, 26, 182–191.e5. [Google Scholar] [CrossRef] [Green Version]
- Muntjes, K.; Devan, S.K.; Reichert, A.S.; Feldbrugge, M. Linking transport and translation of mRNAs with endosomes and mitochondria. EMBO Rep. 2021, 22, e52445. [Google Scholar] [CrossRef]
- Gasparski, A.N.; Mason, D.E.; Moissoglu, K.; Mili, S. Regulation and outcomes of localized RNA translation. Wiley Interdiscip. Rev. RNA 2022, 13, e1721. [Google Scholar] [CrossRef]
- Wheeler, N.A.; Fuss, B. Extracellular cues influencing oligodendrocyte differentiation and (re)myelination. Exp. Neurol. 2016, 283, 512–530. [Google Scholar] [CrossRef] [Green Version]
- Xiao, D.; Su, X.; Gao, H.; Li, X.; Qu, Y. The Roles of Lpar1 in Central Nervous System Disorders and Diseases. Front. Neurosci. 2021, 15, 710473. [Google Scholar] [CrossRef]
- Yung, Y.C.; Stoddard, N.C.; Mirendil, H.; Chun, J. Lysophosphatidic Acid signaling in the nervous system. Neuron 2015, 85, 669–682. [Google Scholar] [CrossRef] [Green Version]
- Nogaroli, L.; Yuelling, L.M.; Dennis, J.; Gorse, K.; Payne, S.G.; Fuss, B. Lysophosphatidic acid can support the formation of membranous structures and an increase in MBP mRNA levels in differentiating oligodendrocytes. Neurochem. Res. 2009, 34, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olah, J.; Lehotzky, A.; Szunyogh, S.; Szenasi, T.; Orosz, F.; Ovadi, J. Microtubule-Associated Proteins with Regulatory Functions by Day and Pathological Potency at Night. Cells 2020, 9, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zotter, A.; Bodor, A.; Olah, J.; Hlavanda, E.; Orosz, F.; Perczel, A.; Ovadi, J. Disordered TPPP/p25 binds GTP and displays Mg2+-dependent GTPase activity. FEBS Lett. 2011, 585, 803–808. [Google Scholar] [CrossRef] [Green Version]
- Vincze, O.; Tokesi, N.; Olah, J.; Hlavanda, E.; Zotter, A.; Horvath, I.; Lehotzky, A.; Tirian, L.; Medzihradszky, K.F.; Kovacs, J.; et al. Tubulin polymerization promoting proteins (TPPPs): Members of a new family with distinct structures and functions. Biochemistry 2006, 45, 13818–13826. [Google Scholar] [CrossRef]
- Kemal, S.; Richardson, H.S.; Dyne, E.D.; Fu, M.M. ER and Golgi trafficking in axons, dendrites, and glial processes. Curr. Opin. Cell Biol. 2022, 78, 102119. [Google Scholar] [CrossRef]
- Chen, M.; Xu, L.; Wu, Y.; Soba, P.; Hu, C. The organization and function of the Golgi apparatus in dendrite development and neurological disorders. Genes Dis. 2022; in press. [Google Scholar] [CrossRef]
- Agrawal, M.; Welshhans, K. Local Translation Across Neural Development: A Focus on Radial Glial Cells, Axons, and Synaptogenesis. Front. Mol. Neurosci. 2021, 14, 717170. [Google Scholar] [CrossRef] [PubMed]
- Trapp, B.D.; Moench, M.; Pulley, E.; Barbosa, E.; Tennekoon, G.I.; Griffin, J. Spatial segregation of mRNA encoding myelin-specific proteins. Proc. Natl. Acad. Sci. USA 1987, 84, 7773–7777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitschopf, H.; Suchanek, G.; Gould, R.M.; Colman, D.R.; Lassmann, H. In situ hybridization with digoxigenin-labeled probes: Sensitive and reliable detection method applied to myelinating rat brain. Acta Neuropathol. 1992, 84, 581–587. [Google Scholar] [CrossRef]
- de Vries, H.; de Jonge, J.C.; Schrage, C.; van der Haar, M.E.; Hoekstra, D. Differential and cell development-dependent localization of myelin mRNAs in oligodendrocytes. J. Neurosci. Res. 1997, 47, 479–488. [Google Scholar] [CrossRef]
- Le, P.; Ahmed, N.; Yeo, G.W. Illuminating RNA biology through imaging. Nat. Cell Biol. 2022, 24, 815–824. [Google Scholar] [CrossRef]
- Pichon, X.; Lagha, M.; Mueller, F.; Bertrand, E. A Growing Toolbox to Image Gene Expression in Single Cells: Sensitive Approaches for Demanding Challenges. Mol. Cell 2018, 71, 468–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safieddine, A.; Coleno, E.; Lionneton, F.; Traboulsi, A.M.; Salloum, S.; Lecellier, C.H.; Gostan, T.; Georget, V.; Hassen-Khodja, C.; Imbert, A.; et al. HT-smFISH: A cost-effective and flexible workflow for high-throughput single-molecule RNA imaging. Nat. Protoc. 2022, 18, 157–187. [Google Scholar] [CrossRef] [PubMed]
- Alon, S.; Goodwin, D.R.; Sinha, A.; Wassie, A.T.; Chen, F.; Daugharthy, E.R.; Bando, Y.; Kajita, A.; Xue, A.G.; Marrett, K.; et al. Expansion sequencing: Spatially precise in situ transcriptomics in intact biological systems. Science 2021, 371, eaax2656. [Google Scholar] [CrossRef] [PubMed]
- Erwig, M.S.; Patzig, J.; Steyer, A.M.; Dibaj, P.; Heilmann, M.; Heilmann, I.; Jung, R.B.; Kusch, K.; Mobius, W.; Jahn, O.; et al. Anillin facilitates septin assembly to prevent pathological outfoldings of central nervous system myelin. eLife 2019, 8, e43888. [Google Scholar] [CrossRef] [PubMed]
- Patzig, J.; Erwig, M.S.; Tenzer, S.; Kusch, K.; Dibaj, P.; Möbius, W.; Goebbels, S.; Schaeren-Wiemers, N.; Nave, K.-A.; Werner, H.B. Septin/anillin filaments scaffold central nervous system myelin to accelerate nerve conduction. eLife 2016, 5, e17119. [Google Scholar] [CrossRef] [PubMed]
- Trapp, B.D.; Quarles, R.H. Presence of the myelin-associated glycoprotein correlates with alterations in the periodicity of peripheral myelin. J. Cell Biol. 1982, 92, 877–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montag, D.; Giese, K.P.; Bartsch, U.; Martini, R.; Lang, Y.; BlÅthmann, H.; Karthigasan, J.; Kirschner, D.A.; Wintergerst, E.S.; Nave, K.A.; et al. Mice deficient for the myelin-associated glycoprotein show subtle abnormalities in myelin. Neuron 1994, 13, 229–246. [Google Scholar] [CrossRef]
- Quarles, R.H. Myelin-associated glycoprotein (MAG): Past, present and beyond. J. Neurochem. 2007, 100, 1431–1448. [Google Scholar] [CrossRef] [PubMed]
- Elazar, N.; Vainshtein, A.; Golan, N.; Vijayaragavan, B.; Schaeren-Wiemers, N.; Eshed-Eisenbach, Y.; Peles, E. Axoglial Adhesion by Cadm4 Regulates CNS Myelination. Neuron 2019, 101, 224–231.e5. [Google Scholar] [CrossRef] [Green Version]
- Moffitt, J.R.; Lundberg, E.; Heyn, H. The emerging landscape of spatial profiling technologies. Nat. Rev. Genet. 2022, 23, 741–759. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Barkley, D.; Franca, G.S.; Yanai, I. Exploring tissue architecture using spatial transcriptomics. Nature 2021, 596, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Longo, S.K.; Guo, M.G.; Ji, A.L.; Khavari, P.A. Integrating single-cell and spatial transcriptomics to elucidate intercellular tissue dynamics. Nat. Rev. Genet. 2021, 22, 627–644. [Google Scholar] [CrossRef]
- Marx, V. Method of the Year: Spatially resolved transcriptomics. Nat. Methods 2021, 18, 9–14. [Google Scholar] [CrossRef]
- Stark, R.; Grzelak, M.; Hadfield, J. RNA sequencing: The teenage years. Nat. Rev. Genet. 2019, 20, 631–656. [Google Scholar] [CrossRef]
- Zhao, W.; Langfelder, P.; Fuller, T.; Dong, J.; Li, A.; Hovarth, S. Weighted gene coexpression network analysis: State of the art. J. Biopharm. Stat. 2010, 20, 281–300. [Google Scholar] [CrossRef]
- Aureli, M.; Grassi, S.; Prioni, S.; Sonnino, S.; Prinetti, A. Lipid membrane domains in the brain. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 1006–1016. [Google Scholar] [CrossRef]
- Cermenati, G.; Mitro, N.; Audano, M.; Melcangi, R.C.; Crestani, M.; De Fabiani, E.; Caruso, D. Lipids in the nervous system: From biochemistry and molecular biology to patho-physiology. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 51–60. [Google Scholar] [CrossRef]
- Marangon, D.; Boccazzi, M.; Lecca, D.; Fumagalli, M. Regulation of Oligodendrocyte Functions: Targeting Lipid Metabolism and Extracellular Matrix for Myelin Repair. J. Clin. Med. 2020, 9, 470. [Google Scholar] [CrossRef] [Green Version]
- Poitelon, Y.; Kopec, A.M.; Belin, S. Myelin Fat Facts: An Overview of Lipids and Fatty Acid Metabolism. Cells 2020, 9, 812. [Google Scholar] [CrossRef] [Green Version]
- Montani, L. Lipids in regulating oligodendrocyte structure and function. Semin. Cell Dev. Biol. 2020, 112, 114–122. [Google Scholar] [CrossRef]
- Wattenberg, B.W. Intra- and intercellular trafficking in sphingolipid metabolism in myelination. Adv. Biol. Regul. 2019, 71, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Barnes-Velez, J.A.; Aksoy Yasar, F.B.; Hu, J. Myelin lipid metabolism and its role in myelination and myelin maintenance. Innovation 2023, 4, 100360. [Google Scholar] [CrossRef] [PubMed]
- Gould, R.M.; Holshek, J.; Silverman, W.; Spivack, W. Localization of phospholipid synthesis to Schwann cells and axons. J. Neurochem. 1987, 48, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Boiron, F.; Spivack, W.D.; Deshmukh, D.S.; Gould, R.M. Basis for phospholipid incorporation into peripheral nerve myelin. J. Neurochem. 1993, 60, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Vos, J.P.; Giudici, M.L.; Van der Bijl, P.; Magni, P.; Marchesini, S.; Van Golde, L.M.G.; Lopes-Cardozo, M. Sphingomyelin is synthesized at the plasma membrane of oligodendrocytes and by purified myelin membranes: A study with fluorescent- and radio-labelled ceramide analogues. FEBS Lett. 1995, 368, 393–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | MSAS mRNAs | Gene Symbol | MSAS |
|---|---|---|---|
| 1 | Apolipoprotein D | APOD | |
| 2 | Dynein light intermediate chain 2 | DYNC1LI2 | ++ |
| 3 | Ectonucleotide pyrophosphatase/phosphodiesterase 2 1 | ENPP2 | |
| 4 | Gelsolin | GSN | |
| 5 | Kinesin family member 1A | KIF1A | ++ |
| 6 | Lysophosphatidic acid receptor 1 2 | LPAR1 | |
| 7 | Mitogen-activated protein kinase 8 interacting protein-1 3 | MAPK8IP1 | |
| 8 | N-myc downstream regulated 1 | NDRG1 | |
| 9 | Peptidylarginine deiminase 2 | PADI2 | ++ |
| 10 | SH3 domain containing GRB2 like 3/endophilin-A3 | SH3GL3 | ++ |
| 11 | Tubulin polymerization promoting protein 4 | TPPP | |
| 12 | Trafficking kinesin protein 2 5 | TRAK2 | ++ |
| 13 | Tumor-promoter p53-inducible nuclear protein-2 6 | TRP53INP2 | ++ |
| 14 | Carbonic anhydrase 2 | CAR2 | |
| 15 | Crystallin alpha B | CRYAB |
| # | mRNA | SSH | %MBP | mmRS | %MBP | SSH/RS |
|---|---|---|---|---|---|---|
| kMSAS1 | MBP | 44,468 ± 20,315 (10) | 100.00 | 1,265,436 ± 290,298 | 100.00 | 1.00 |
| kMSAS2 | MOBP-81A | 1191 ± 225 (5) | 2.68 | 93,503 ± 31,016 * | 7.39 | 0.36 |
| OLS1 | PLP1 | 1096 ± 471 (4) | 2.46 | 88,651 ± 6278 | 7.01 | 0.35 |
| OLS2 | MOBP-81B | 36 ± 17 (6) | 0.08 | -- | -- | -- |
| OLS3 | CNP | 28 ± 12 (4) | 0.06 | 10,985 ± 597 | 0.87 | 0.07 |
| NE1 | NEFL | 8 ± 3 (5) | 0.02 | 5693 ± 255 | 0.45 | 0.04 |
| AST1 | GFAP | 11 ± 4 (3) | 0.02 | 5729 ± 296 | 0.45 | 0.04 |
| MSAS1 | LPAR1 | 156 ± 102 (10) | 0.35 | 6673 ± 984 | 0.53 | 0.66 |
| MSAS2 | NDRG1 | 103 ± 33 (6) | 0.23 | 6334 ± 83 | 0.50 | 0.46 |
| MSAS3 | TRAK2 | 41 ± 25 (11) | 0.09 | 19,253 ± 5371 | 1.52 | 0.06 |
| MSAS4 | APOD | 33 ± 14 (11) | 0.07 | 12,428 ± 1338 | 0.98 | 0.07 |
| MSAS5 | KIF1A | 23 ± 11 (11) | 0.05 | 15,742 ± 2277 | 1.24 | 0.04 |
| MSAS6 | TPPP | 19 ± 15 (9) | 0.04 | 9353 ± 2101 | 0.74 | 0.05 |
| MSAS7 | SH3GL3 | 17 ± 4 (13) | 0.04 | 4343 ± 347 | 0.34 | 0.12 |
| MSAS8 | ENPP2 | 7 ± 2 (11) | 0.02 | 12,016 ± 1371 | 0.95 | 0.02 |
| MSAS9 | GSN | 3 ± 2 (8) | 0.01 | 1992 ± 181 | 0.16 | 0.06 |
| MSAS10 | MAPK81P1 | 2 ± 2 (8) | 0.00 | 5269 ± 205 | 0.42 | <0.01 |
| MSAS11 | TRP53INP2 | 1 ± 1 (7) | 0.00 | 491 ± 43 | 0.04 | <0.01 |
| MSAS12 | PADI2 | -- | -- | 760 ± 71 | 0.06 | -- |
| MSAS13 | DYNC1LI2 | -- | -- | 17,565 ± 4234 | 1.39 | -- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gould, R.; Brady, S. Identifying mRNAs Residing in Myelinating Oligodendrocyte Processes as a Basis for Understanding Internode Autonomy. Life 2023, 13, 945. https://doi.org/10.3390/life13040945
Gould R, Brady S. Identifying mRNAs Residing in Myelinating Oligodendrocyte Processes as a Basis for Understanding Internode Autonomy. Life. 2023; 13(4):945. https://doi.org/10.3390/life13040945
Chicago/Turabian StyleGould, Robert, and Scott Brady. 2023. "Identifying mRNAs Residing in Myelinating Oligodendrocyte Processes as a Basis for Understanding Internode Autonomy" Life 13, no. 4: 945. https://doi.org/10.3390/life13040945