

Impact of Sex and Exercise on Femoral Artery Function: More Favorable Adaptation in Male Rats
 , , , and
, , , and 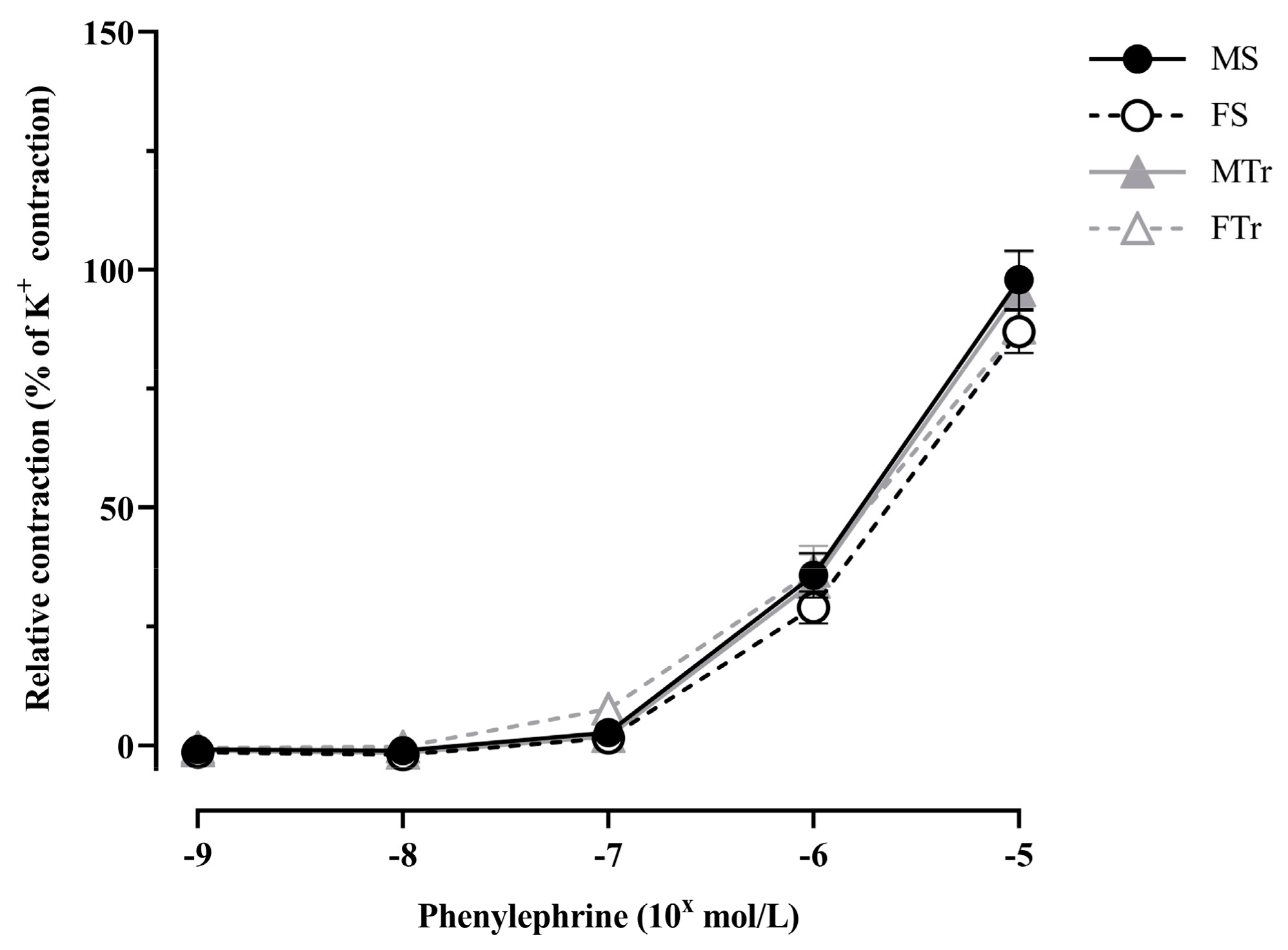{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Ethical Approval and Animals
2.3. Experimental Groups
2.4. Myography
2.5. Histological and Immunochemical Examinations
2.6. Statistical Analysis
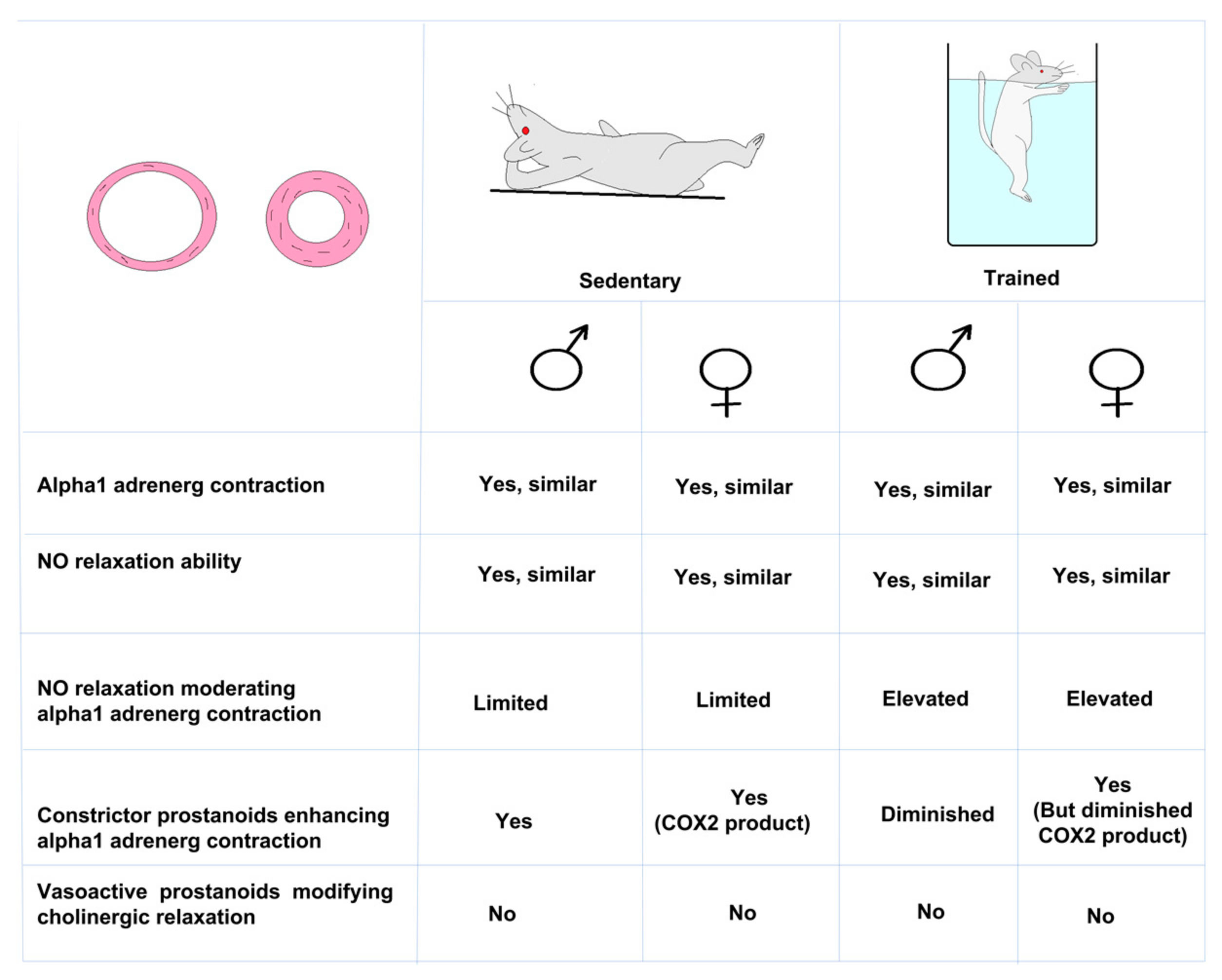
3. Results
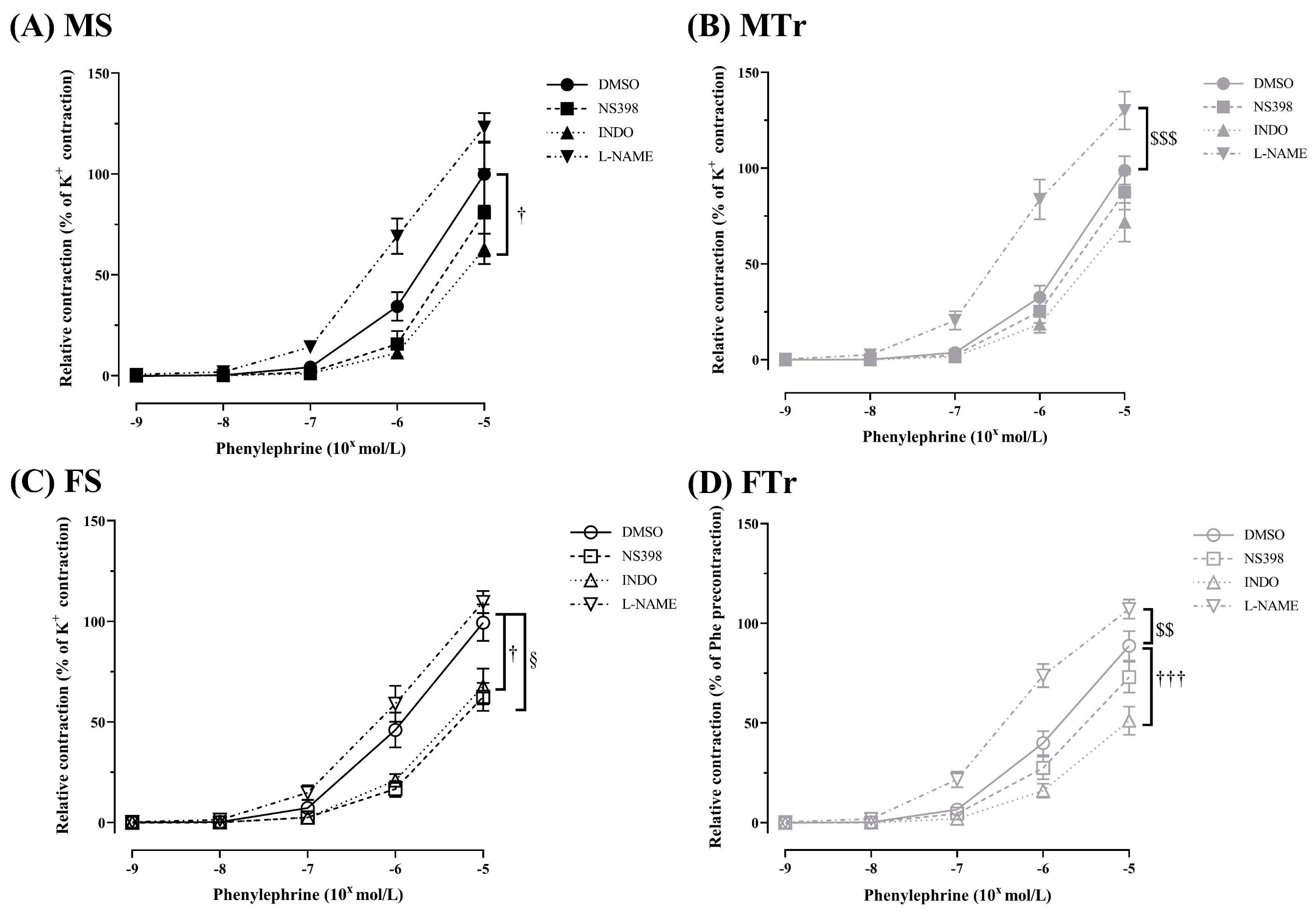
3.1. Contractility of Femoral Arteries
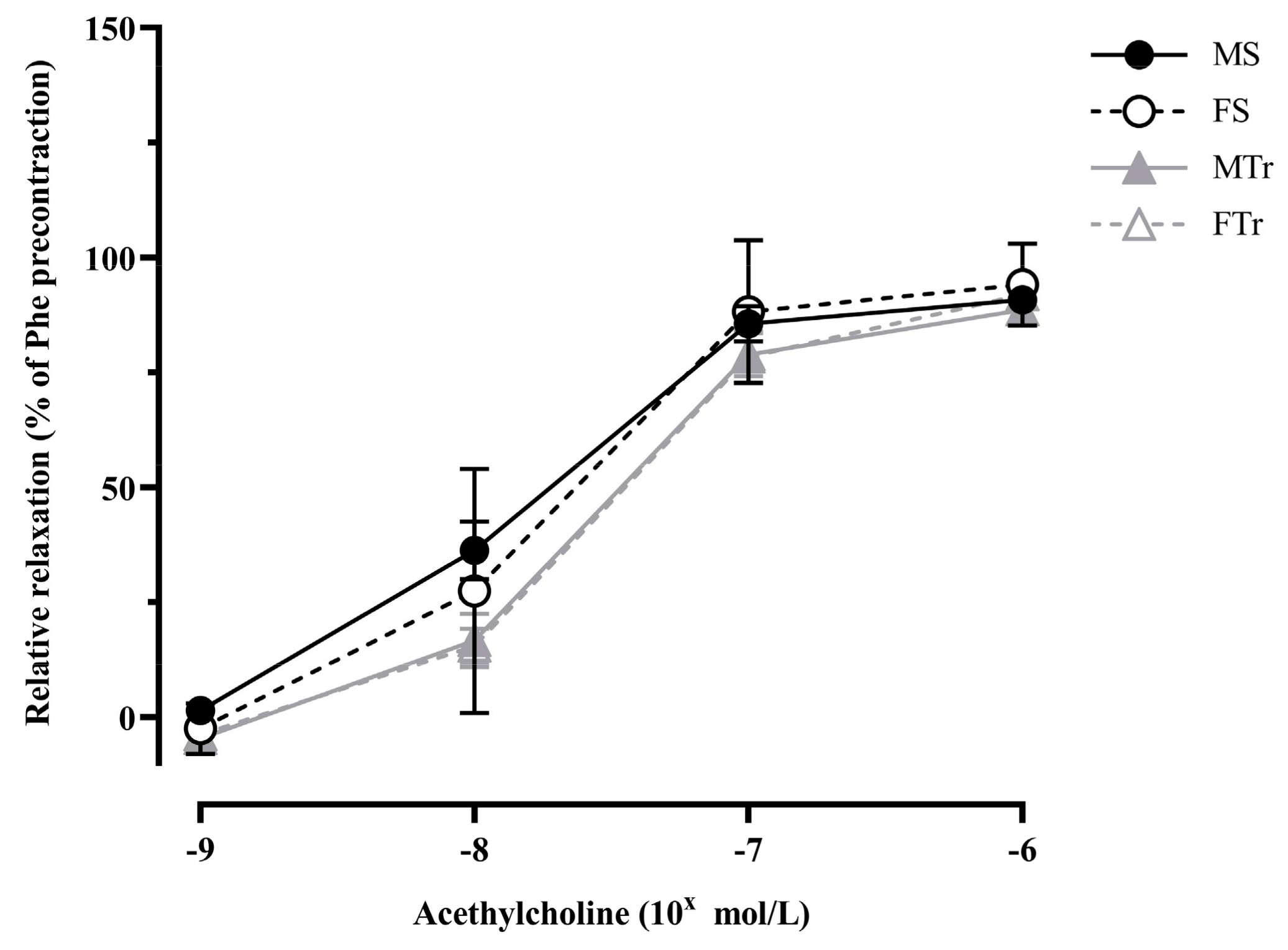
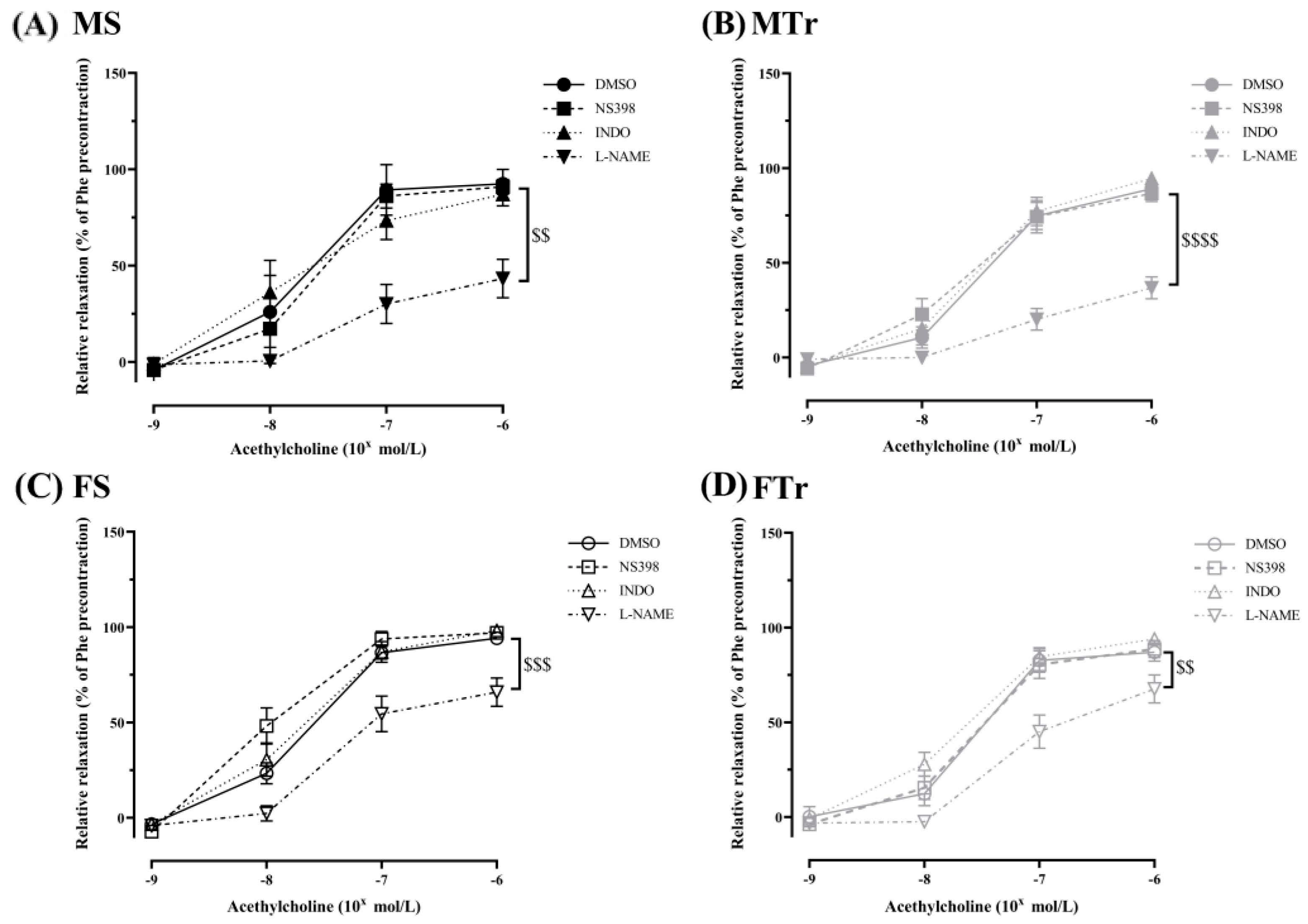
3.2. Relaxation Ability of Femoral Arteries
3.3. Histological Alterations
4. Discussion
Strengths and Limitations of the Paper
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanhees, L.; De Sutter, J.; Gelada, S.N.; Doyle, F.; Prescott, E.; Cornelissen, V.; Kouidi, E.; Dugmore, D.; Vanuzzo, D.; Börjesson, M.; et al. Importance of characteristics and modalities of physical activity and exercise in defining the benefits to cardiovascular health within the general population: Recommendations from the EACPR (Part I). Eur. J. Prev. Cardiol. 2012, 19, 670–686. [Google Scholar] [CrossRef] [PubMed]
- Beets, M.W.; Beighle, A.; Erwin, H.E.; Huberty, J.L. After-school program impact on physical activity and fitness: A meta-analysis. Am. J. Prev. Med. 2009, 36, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A. An Overview of the Beneficial Effects of Exercise on Health and Performance. Adv. Exp. Med. Biol. 2020, 1228, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Heinicke, K.; Wolfarth, B.; Winchenbach, P.; Biermann, B.; Schmid, A.; Huber, G.; Friedmann, B.; Schmidt, W. Blood volume and hemoglobin mass in elite athletes of different disciplines. Int. J. Sports Med. 2001, 22, 504–512. [Google Scholar] [CrossRef]
- Joyner, M.J.; Casey, D.P. Regulation of increased blood flow (hyperemia) to muscles during exercise: A hierarchy of competing physiological needs. Physiol. Rev. 2015, 95, 549–601. [Google Scholar] [CrossRef] [PubMed]
- Prior, B.M.; Yang, H.T.; Terjung, R.L. What makes vessels grow with exercise training? J. Appl. Physiol. 2004, 97, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Hellsten, Y.; Nyberg, M. Cardiovascular Adaptations to Exercise Training. Compr. Physiol. 2015, 6, 1–32. [Google Scholar] [CrossRef]
- Bassett, A.J.; Ahlmen, A.; Rosendorf, J.M.; Romeo, A.A.; Erickson, B.J.; Bishop, M.E. The Biology of Sex and Sport. JBJS Rev. 2020, 8, e0140. [Google Scholar] [CrossRef]
- Thibault, V.; Guillaume, M.; Berthelot, G.; Helou, N.E.; Schaal, K.; Quinquis, L.; Nassif, H.; Tafflet, M.; Escolano, S.; Hermine, O.; et al. Women and Men in Sport Performance: The Gender Gap has not Evolved since 1983. J. Sports Sci. Med. 2010, 9, 214–223. [Google Scholar]
- Török, M.; Horváth, E.M.; Monori-Kiss, A.; PÁl, É.; Gerszi, D.; Merkely, P.; Sayour, A.A.; Mátyás, C.; Oláh, A.; Radovits, T.; et al. Chronic swimming training resulted in more relaxed coronary arterioles in male and enhanced vasoconstrictor ability in female rats. J. Sports Med. Phys. Fitness 2020, 61, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Merkely, P.; Bakos, M.; Bányai, B.; Monori-Kiss, A.; Horváth, E.M.; Bognár, J.; Benkő, R.; Oláh, A.; Radovits, T.; Merkely, B.; et al. Sex Differences in Exercise-Training-Related Functional and Morphological Adaptation of Rat Gracilis Muscle Arterioles. Front. Physiol. 2021, 12, 685664. [Google Scholar] [CrossRef]
- Vezér, M.; Demeter, Á.; Szekeres, M.; Jósvai, A.; Bányai, B.; Oláh, A.; Balogh, F.; Horváth, E.M.; Radovits, T.; Merkely, B.; et al. Sex differences in rat renal arterial responses following exercise training. Am. J. Physiol. Heart Circ. Physiol. 2022, 322, H310–H318. [Google Scholar] [CrossRef]
- Vanhoutte, P.M.; Shimokawa, H.; Feletou, M.; Tang, E.H. Endothelial dysfunction and vascular disease—A 30th anniversary update. Acta Physiol. 2017, 219, 22–96. [Google Scholar] [CrossRef] [PubMed]
- Radovits, T.; Olah, A.; Lux, A.; Nemeth, B.T.; Hidi, L.; Birtalan, E.; Kellermayer, D.; Matyas, C.; Szabo, G.; Merkely, B. Rat model of exercise-induced cardiac hypertrophy: Hemodynamic characterization using left ventricular pressure-volume analysis. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H124–H134. [Google Scholar] [CrossRef] [PubMed]
- Pavlik, G. Effects of physical training and detraining on resting cardiovascular parameters in albino rats. Acta Physiol. Hung. 1985, 66, 27–37. [Google Scholar]
- Horváth, B.; Orsy, P.; Benyó, Z. Endothelial NOS-mediated relaxations of isolated thoracic aorta of the C57BL/6J mouse: A methodological study. J. Cardiovasc. Pharmacol. 2005, 45, 225–231. [Google Scholar] [CrossRef]
- Szekeres, M.; Nádasy, G.L.; Turu, G.; Soltész-Katona, E.; Benyó, Z.; Offermanns, S.; Ruisanchez, É.; Szabó, E.; Takáts, Z.; Bátkai, S.; et al. Endocannabinoid-mediated modulation of Gq/11 protein-coupled receptor signaling-induced vasoconstriction and hypertension. Mol. Cell Endocrinol. 2015, 403, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Koller, A.; Huang, A.; Sun, D.; Kaley, G. Exercise training augments flow-dependent dilation in rat skeletal muscle arterioles. Role of endothelial nitric oxide and prostaglandins. Circ. Res. 1995, 76, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.; Garabito, M.; Jiménez-Altayó, F.; Onetti, Y.; Sabate, M.; Vila, E.; Dantas, A.P. Sex differences in angiotensin II responses contribute to a differential regulation of cox-mediated vascular dysfunction during aging. Exp. Gerontol. 2016, 85, 71–80. [Google Scholar] [CrossRef]
- Fulton, C.T.; Stallone, J.N. Sexual dimorphism in prostanoid-potentiated vascular contraction: Roles of endothelium and ovarian steroids. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H2062–H2073. [Google Scholar] [CrossRef]
- Sullivan, J.C.; Sasser, J.M.; Pollock, D.M.; Pollock, J.S. Sexual dimorphism in renal production of prostanoids in spontaneously hypertensive rats. Hypertension 2005, 45, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Stallone, J.N. Estrogen potentiates vasopressin-induced contraction of female rat aorta by enhancing cyclooxygenase-2 and thromboxane function. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H1542–H1550. [Google Scholar] [CrossRef]
- Deer, R.R.; Stallone, J.N. Effects of age and sex on cerebrovascular function in the rat middle cerebral artery. Biol. Sex Differ. 2014, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Macedo, F.N.; Mesquita, T.R.; Melo, V.U.; Mota, M.M.; Silva, T.L.; Santana, M.N.; Oliveira, L.R.; Santos, R.V.; Miguel Dos Santos, R.; Lauton-Santos, S.; et al. Increased Nitric Oxide Bioavailability and Decreased Sympathetic Modulation Are Involved in Vascular Adjustments Induced by Low-Intensity Resistance Training. Front. Physiol. 2016, 7, 265. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.L.; Tsai, S.J.; Jiang, M.J.; Jen, C.J.; Chen, H.I. Chronic exercise increases both inducible and endothelial nitric oxide synthase gene expression in endothelial cells of rat aorta. J. BioMed. Sci. 2002, 9, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Marchio, P.; Guerra-Ojeda, S.; Vila, J.M.; Aldasoro, M.; Valles, S.L.; Soler, C.; Mauricio, M.D. Chronic exercise impairs nitric oxide pathway in rabbit carotid and femoral arteries. J. Physiol. 2018, 596, 4361–4374. [Google Scholar] [CrossRef]
- McAllister, R.M.; Jasperse, J.L.; Laughlin, M.H. Nonuniform effects of endurance exercise training on vasodilation in rat skeletal muscle. J. Appl. Physiol. 2005, 98, 753–761. [Google Scholar] [CrossRef]
- McAllister, R.M.; Kimani, J.K.; Webster, J.L.; Parker, J.L.; Laughlin, M.H. Effects of exercise training on responses of peripheral and visceral arteries in swine. J. Appl. Physiol. 1996, 80, 216–225. [Google Scholar] [CrossRef]
- Delp, M.D.; Laughlin, M.H. Time course of enhanced endothelium-mediated dilation in aorta of trained rats. Med. Sci. Sports Exerc. 1997, 29, 1454–1461. [Google Scholar] [CrossRef]
- Pabbidi, M.R.; Kuppusamy, M.; Didion, S.P.; Sanapureddy, P.; Reed, J.T.; Sontakke, S.P. Sex differences in the vascular function and related mechanisms: Role of 17β-estradiol. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1499–H1518. [Google Scholar] [CrossRef]
- Laughlin, M.H.; Schrage, W.G.; McAllister, R.M.; Garverick, H.A.; Jones, A.W. Interaction of gender and exercise training: Vasomotor reactivity of porcine skeletal muscle arteries. J. Appl. Physiol. 2001, 90, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, M.G.; Vanetti, C.; Decimo, I.; Di Chio, M.; Martano, G.; Garrone, G.; Bifari, F.; Vicentini, L.M. Sex-specific eNOS activity and function in human endothelial cells. Sci. Rep. 2017, 7, 9612. [Google Scholar] [CrossRef] [PubMed]
- Ansdell, P.; Thomas, K.; Hicks, K.M.; Hunter, S.K.; Howatson, G.; Goodall, S. Physiological sex differences affect the integrative response to exercise: Acute and chronic implications. Exp. Physiol. 2020, 105, 2007–2021. [Google Scholar] [CrossRef] [PubMed]
- Badrov, M.B.; Freeman, S.R.; Zokvic, M.A.; Millar, P.J.; McGowan, C.L. Isometric exercise training lowers resting blood pressure and improves local brachial artery flow-mediated dilation equally in men and women. Eur. J. Appl. Physiol. 2016, 116, 1289–1296. [Google Scholar] [CrossRef]
- Shenouda, N.; Skelly, L.E.; Gibala, M.J.; MacDonald, M.J. Brachial artery endothelial function is unchanged after acute sprint interval exercise in sedentary men and women. Exp. Physiol. 2018, 103, 968–975. [Google Scholar] [CrossRef]
- Pierce, G.L.; Eskurza, I.; Walker, A.E.; Fay, T.N.; Seals, D.R. Sex-specific effects of habitual aerobic exercise on brachial artery flow-mediated dilation in middle-aged and older adults. Clin. Sci. 2011, 120, 13–23. [Google Scholar] [CrossRef]
- Dietz, N.M. Gender and nitric oxide-mediated vasodilation in humans. Lupus 1999, 8, 402–408. [Google Scholar] [CrossRef]
- Nishiyama, S.K.; Wray, D.W.; Richardson, R.S. Sex and limb-specific ischemic reperfusion and vascular reactivity. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1100–H1108. [Google Scholar] [CrossRef]
- Cureton, K.J.; Collins, M.A.; Hill, D.W.; McElhannon, F.M., Jr. Muscle hypertrophy in men and women. Med. Sci. Sports Exerc. 1988, 20, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Török, M.; Monori-Kiss, A.; Pal, E.; Horvath, E.; Josvai, A.; Merkely, P.; Barta, B.A.; Matyas, C.; Olah, A.; Radovits, T.; et al. Long-term exercise results in morphological and biomechanical changes in coronary resistance arterioles in male and female rats. Biol. Sex Differ. 2020, 11, 7. [Google Scholar] [CrossRef]
- Laughlin, M.H.; Pollock, J.S.; Amann, J.F.; Hollis, M.L.; Woodman, C.R.; Price, E.M. Training induces nonuniform increases in eNOS content along the coronary arterial tree. J. Appl. Physiol. 2001, 90, 501. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vezér, M.; Jósvai, A.; Bányai, B.; Ács, N.; Keszthelyi, M.; Soltész-Katona, E.; Szekeres, M.; Oláh, A.; Radovits, T.; Merkely, B.; et al. Impact of Sex and Exercise on Femoral Artery Function: More Favorable Adaptation in Male Rats. Life 2023, 13, 778. https://doi.org/10.3390/life13030778
Vezér M, Jósvai A, Bányai B, Ács N, Keszthelyi M, Soltész-Katona E, Szekeres M, Oláh A, Radovits T, Merkely B, et al. Impact of Sex and Exercise on Femoral Artery Function: More Favorable Adaptation in Male Rats. Life. 2023; 13(3):778. https://doi.org/10.3390/life13030778
Chicago/Turabian StyleVezér, Márton, Attila Jósvai, Bálint Bányai, Nándor Ács, Márton Keszthelyi, Eszter Soltész-Katona, Mária Szekeres, Attila Oláh, Tamás Radovits, Béla Merkely, and et al. 2023. "Impact of Sex and Exercise on Femoral Artery Function: More Favorable Adaptation in Male Rats" Life 13, no. 3: 778. https://doi.org/10.3390/life13030778