
Effect of Fermentation on the Nutritional Quality of the Selected Vegetables and Legumes and Their Health Effects


Abstract
:1. Introduction
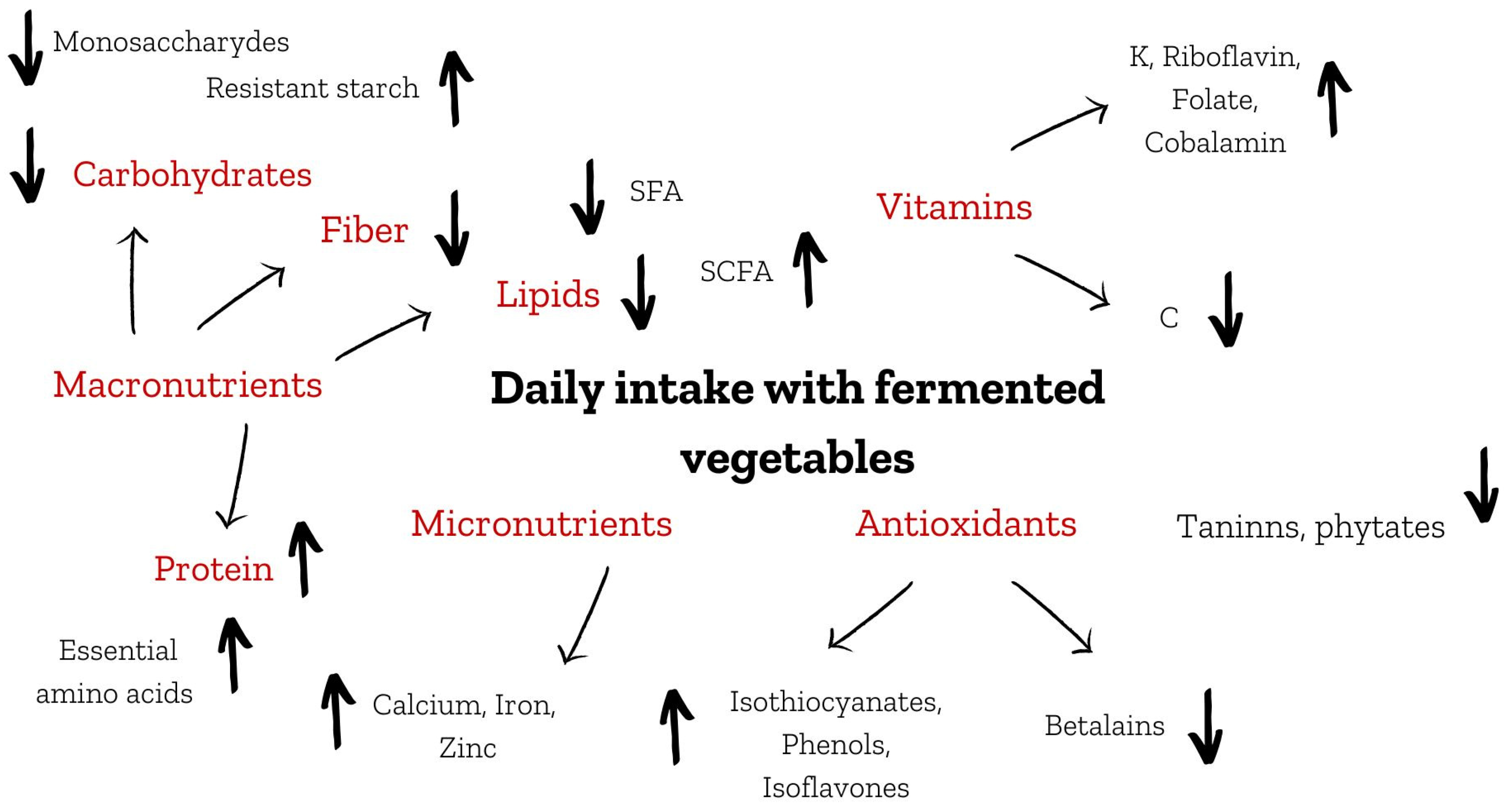
2. Macronutrients
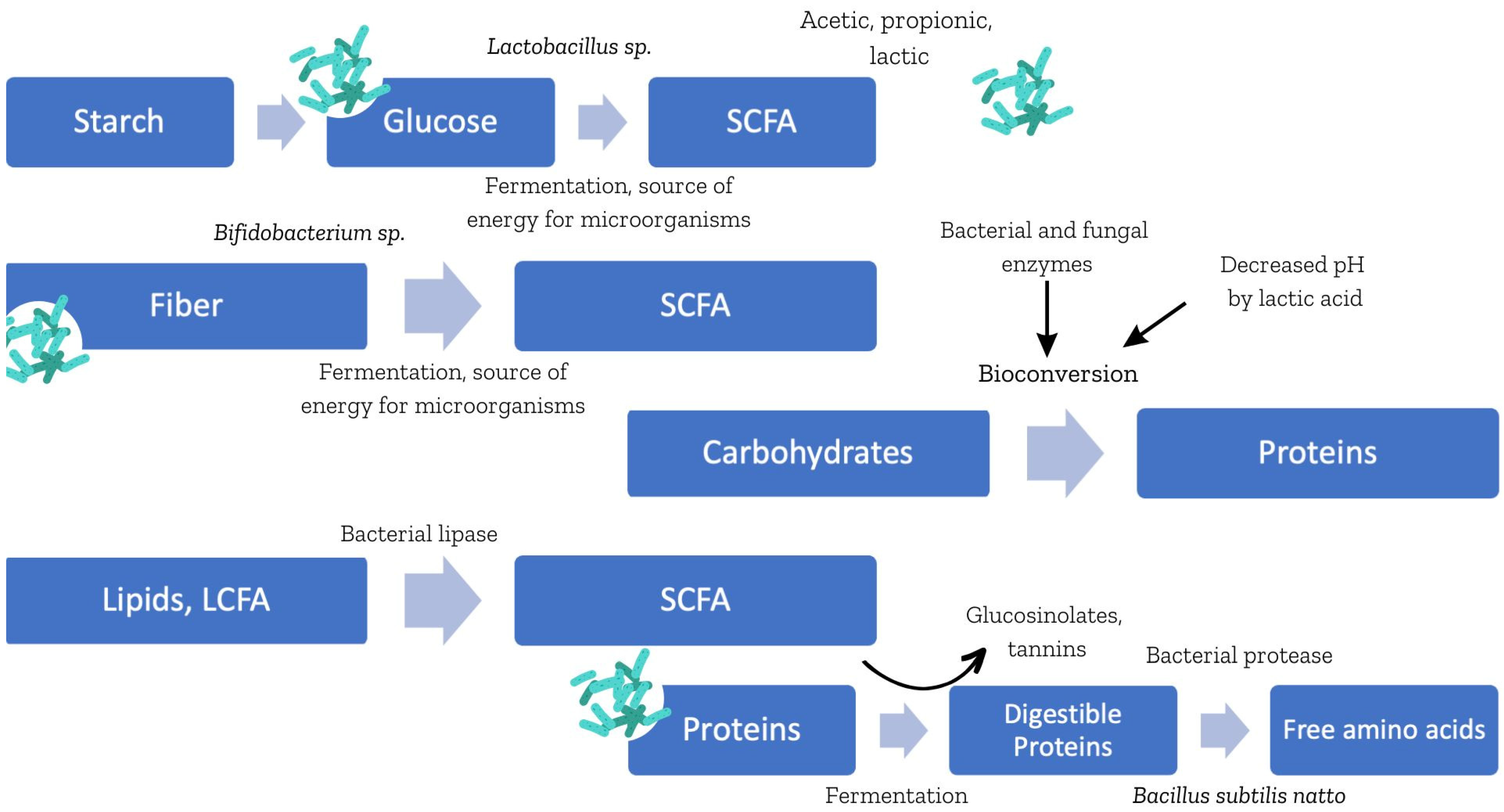
2.1. Carbohydrates and Dietary Fiber
2.2. Proteins
2.3. Lipids
3. Micronutrients vs. Anti-Nutritional Compounds
4. Vitamins
5. Antioxidant Potential
5.1. Antioxidants Content
5.2. Antioxidant Potential

{kind=link}
{kind=link}
{kind=link}
| Content of | ||||||
|---|---|---|---|---|---|---|
| Product | Mineral Components | Anti-Nutritional Compounds | Vitamins | Bioactive Compounds | Bioavailability | Source |
| Fermented bamboo shoots | ↓ Ca, Mg, Mn, K, Zn, Fe | ↓ phytic acid | n.d. | ↑ phenols, flavonoids | ↑ Fe | [31] |
| Sauerkraut | ↓ Ca, Mg, Zn, Fe | ↓ GLS | vit. C ↓ | ↑ indol-3-acetonitrile, indol-3-carbinol, ascorbigen ↑ TPC | ↑ indol-3-acetonitrile, indol-3-carbinol | [19,54,55,56,71] |
| Cucumber, pickled | ↑ Ca ↓ Mg, Zn, Fe | n.d. | ↑ riboflavin ↑ folate, total | ↑ ACE inhibitors ↑ GABA | ↑ FAA (lysine, leucine, isoleucine) | [19,57,70,72] |
| Legumes, fermented, other than soya | ↑ Fe, Ca ↓ Zn | ↓ tannins ↓ phytic acid | ↑ riboflavin ↓ thiamine | ↑ phenols, polyphenols | ↑ Fe, Zn ↑ FAA (lysine) | [29,37,45,73] |
| Soya, fermented | n.d. | ↓ phytic acid | ↑ riboflavin ↑ cobalamin ↑ folate, vit. K | ↑ phenols ↑ GABA ↑ genistein ↑ daidzein ↑ aglicons ↑ SCFA | ↑ isoflavones ↑ phytoestrogens ↑ Zn ↑ protein, FAA | [16,47,49,74,75] |
6. Other Bioactive Compounds
6.1. Inhibitors of Angiotensin-Converting Enzyme
6.2. GABA—γ-Aminobutyric Acid
6.3. Isoflavones and Phytoestrogens
7. Microbiological Composition of Fermented Vegetables
7.1. Fermented Roots
7.2. Sauerkraut
7.3. Kimchi
7.4. Pickled Cucumbers
7.5. Fermented Soy Products
| Vegetable | Microorganisms | Source |
|---|---|---|
| Sauerkraut | Lactiplantibacillus plantarum; Lactobacillus: delbrueckii, curvatus; Staphylococcus epidermidis; Levilactobacillus brevis, Lacticaseibacillus: casei, paracasei HD 1.7; Weissella confuse; Lactococcus lactis Leuconostoc: citreum, mesenteroides | [71,91,92,93] |
| Beetroot, pickled | Lactiplantibacillus: plantarum, pentosus; Lacticaseibacillus: rhamnosus, paracaseil; Lentilactobacillus otakiensis | [86,87] |
| Kimchi | Leuconostoc: mesenteroides lactis, gasicomitatum, gelidum, karnosum, inhae, kimchii, miyukkimchii; Lactobacillus: kimchii, kimchiensis, koreensis; Levilactobacillus brevis; Lactococcus kimchi; Weissella koreensis | [96,106] |
| Cucumber, pickled | Lactiplantibacillus: plantarum, pentosus; Levilactobacillus brevis; Leuconostoc: lactis, holzapfelii, fallax, mesenteroides | [97,98,99,100] |
| Soy sauce | Bacillus spp., Weissella spp., Klebsiella: variicola, pneumonia | [92] |
| Carrot Kanji, pickled | Lactiplantibacillus: paraplantarum, pentosus | [86] |
| Carrot, pickled | Enterobacteriaceae: Erwinia, Serratia; Leuconostac spp.; Lactococcus spp. | [107] |
| Radish, pickled | Enterobacteriaceae: Erwinia, Serratia; Leuconostoc spp.; Lactococcus spp. | [107] |
| Pepper, pickled | Enterobacteriaceae; Lactococcus spp.; Bacillus spp.; Leuconostoc spp.; Levilactobacillus brevis | [107] |
| Tempeh | Rhizopus: oligosporus, oryaze; Enterococcus faecium; Leuconostoc: lactis; Lactobacillus delbruecki spp. Delbrueckii; Alicyclobacillus spp. | [108,109] |
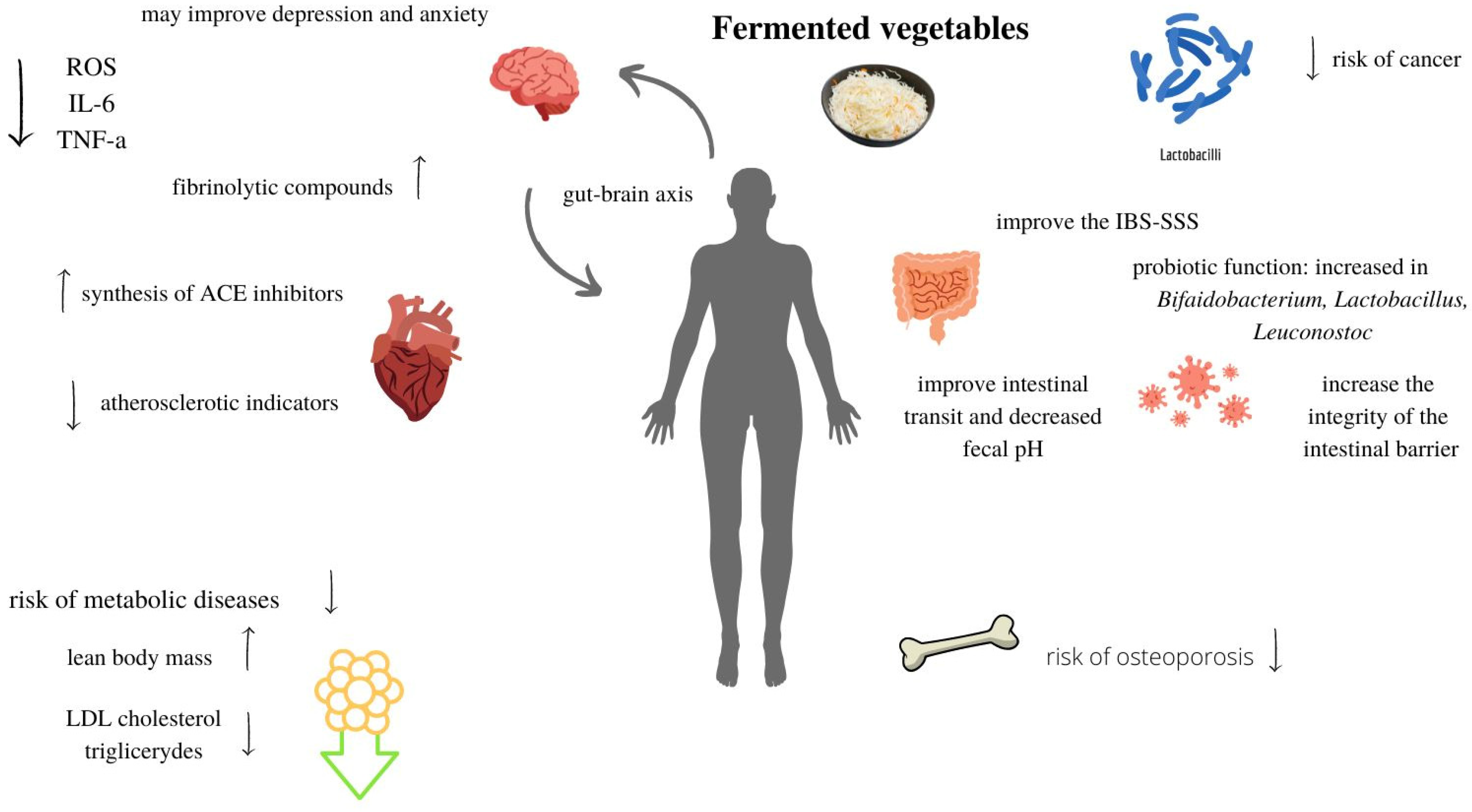
8. Health Effects of Fermented Vegetables
8.1. Probiotic Function
8.2. Immune Function
8.3. Antioxidant Function
8.4. Metabolic Function
8.5. Health Risk Related to High Salinity of Fermented Vegetables
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gasbarrini, G.; Bonvicini, F.; Gramenzi, A. Probiotics History. J. Clin. Gastroenterol. 2016, 50, 116–119. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Puniya, A.K.; Dhewa, T. Enhancing Micronutrients Bioavailability through Fermentation of Plant-Based Foods: A Concise Review. Fermentation 2021, 7, 63. [Google Scholar] [CrossRef]
- Bonatsou, S.; Iliopoulos, V.; Mallouchos, A.; Gogou, E.; Oikonomopoulou, V.; Krokida, M.; Taoukis, P.; Panagou, E.Z. Effect of Osmotic Dehydration of Olives as Pre-Fermentation Treatment and Partial Substitution of Sodium Chloride by Monosodium Glutamate in the Fermentation Profile of Kalamata Natural Black Olives. Food Microbiol. 2017, 63, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Beuchat, L.R.; Perkins, L.M.; Nakayama, T. Fermentation and Sensory Characteristics of Kimchi Containing Potassium Chloride as a Partial Replacement for Sodium Chloride. Int. J. Food Microbiol. 1994, 21, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Lee, Y.C.; Kim, S.S.; do Hong, H.; Kim, K.T. Quality and Antioxidant Activity of Ginseng Seed Processed by Fermentation Strains. J. Ginseng Res. 2015, 39, 178. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.P.; Lee, E.J.; Kim, Y.H.; Ahn, D.U. Effect of Fermentation Temperature on the Volatile Composition of Kimchi. J. Food Sci. 2016, 81, 2623–2629. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Hondrodimou, O.L.; Drosinos, E.H. Development of the Microbial Community during Spontaneous Cauliflower Fermentation. Food Res. Int. 2010, 43, 1098–1103. [Google Scholar] [CrossRef]
- McFeeters, R.F.; Pérez-Díaz, I. Fermentation of Cucumbers Brined with Calcium Chloride Instead of Sodium Chloride. J. Food Sci. 2010, 75, 291–296. [Google Scholar] [CrossRef]
- Ballesteros, C.; Palop, L.; Sánchez, I. Influence of Sodium Chloride Concentration on the Controlled Lactic Acid Fermentation of “Almagro” Eggplants. Int. J. Food Microbiol. 1999, 53, 13–20. [Google Scholar] [CrossRef]
- Ma, T.; Yang, C.; Cai, F.; Cui, L.; Wang, Y. Optimizing Fermentation of Bacillus Amyloliquefaciens 3–5 and Determining Disease Suppression and Growth in Cucumber (Cucumis Sativus). Biol. Control 2022, 176, 105070. [Google Scholar] [CrossRef]
- Pranoto, Y.; Anggrahini, S.; Efendi, Z. Effect of Natural and Lactobacillus Plantarum Fermentation on In-Vitro Protein and Starch Digestibilities of Sorghum Flour. Food Biosci. 2013, 2, 46–52. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant Food Anti-Nutritional Factors and Their Reduction Strategies: An Overview. Food Prod. Process. Nutr. 2020, 2, 6. [Google Scholar] [CrossRef]
- Gupta, R.K.; Gangoliya, S.S.; Singh, N.K. Reduction of Phytic Acid and Enhancement of Bioavailable Micronutrients in Food Grains. J. Food Sci. Technol. 2013, 52, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Takagi, A.; Kano, M.; Kaga, C. Possibility of Breast Cancer Prevention: Use of Soy Isoflavones and Fermented Soy Beverage Produced Using Probiotics. Int. J. Mol. Sci. 2015, 16, 10907–10920. [Google Scholar] [CrossRef] [PubMed]
- Beltrán-Barrientos, L.M.; Hernández-Mendoza, A.; Torres-Llanez, M.J.; González-Córdova, A.F.; Vallejo-Córdoba, B. Invited Review: Fermented Milk as Antihypertensive Functional Food. J. Dairy Sci. 2016, 99, 4099–4110. [Google Scholar] [CrossRef]
- Chen, X.; Lu, Y.; Zhao, A.; Wu, Y.; Zhang, Y.; Yang, X. Quantitative Analyses for Several Nutrients and Volatile Components during Fermentation of Soybean by Bacillus Subtilis Natto. Food Chem. 2022, 374, 131725. [Google Scholar] [CrossRef]
- Hebert, E.M.; de Giori, G.S.; Mozzi, F. Nutraceutics and High Value Metabolites Produced by Lactic Acid Bacteria. In Biotechnology of Lactic Acid Bacteria: Novel Applications, 2nd ed.; Willey Blackwell: Hoboken, NJ, USA, 2015; pp. 297–313. [Google Scholar] [CrossRef]
- Guo, Q.; Chen, P.; Chen, X. Bioactive Peptides Derived from Fermented Foods: Preparation and Biological Activities. J. Funct. Foods 2023, 101, 105422. [Google Scholar] [CrossRef]
- Food Data Central. Available online: https://fdc.nal.usda.gov/ (accessed on 21 June 2022).
- Nilsson, M.; Liljeberg, H.G.M.; Êhl, E.; Molin, G.; Bjo Èrck, I.M.E. On the Effect of Lactic Acid on Blood Glucose and Insulin Responses to Cereal Products: Mechanistic Studies in Healthy Subjects and In Vitro. J. Cereal Sci. 2002, 36, 339–346. [Google Scholar] [CrossRef]
- Asensio-Grau, A.; Calvo-Lerma, J.; Heredia, A.; Andrés, A. Enhancing the Nutritional Profile and Digestibility of Lentil Flour by Solid State Fermentation with Pleurotus ostreatus. Food Funct. 2020, 11, 7905–7912. [Google Scholar] [CrossRef]
- Espinosa-Páez, E.; Alanis-Guzmán, M.G.; Hernández-Luna, C.E.; Báez-González, J.G.; Amaya-Guerra, C.A.; Andrés-Grau, A.M. Increasing Antioxidant Activity and Protein Digestibility in Phaseolus Vulgaris and Avena Sativa by Fermentation with the Pleurotus Ostreatus Fungus. Molecules 2017, 22, 2275. [Google Scholar] [CrossRef]
- Xu, Y.; Hlaing, M.M.; Glagovskaia, O.; Augustin, M.A.; Terefe, N.S. Fermentation by Probiotic Lactobacillus Gasseri Strains Enhances the Carotenoid and Fibre Contents of Carrot Juice. Foods 2020, 9, 1803. [Google Scholar] [CrossRef]
- Dalile, B.; van Oudenhove, L.; Vervliet, B.; Verbeke, K. The Role of Short-Chain Fatty Acids in Microbiota-Gut-Brain Communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.G.; Qoronfleh, M.W. Vegetables. Adv. Neurobiol. 2020, 24, 225–277. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, F.; Gardner, C.D. Dietary Protein and Amino Acids in Vegetarian Diets—A Review. Nutrients 2019, 11, 2661. [Google Scholar] [CrossRef] [PubMed]
- Clarys, P.; Deliens, T.; Huybrechts, I.; Deriemaeker, P.; Vanaelst, B.; de Keyzer, W.; Hebbelinck, M.; Mullie, P. Comparison of Nutritional Quality of the Vegan, Vegetarian, Semi-Vegetarian, Pesco-Vegetarian and Omnivorous Diet. Nutrients 2014, 6, 1318–1332. [Google Scholar] [CrossRef]
- Curiel, J.A.; Coda, R.; Centomani, I.; Summo, C.; Gobbetti, M.; Rizzello, C.G. Exploitation of the Nutritional and Functional Characteristics of Traditional Italian Legumes: The Potential of Sourdough Fermentation. Int. J. Food Microbiol. 2015, 196, 51–61. [Google Scholar] [CrossRef]
- Chinma, C.E.; Azeez, S.O.; Sulayman, H.T.; Alhassan, K.; Alozie, S.N.; Gbadamosi, H.D.; Danbaba, N.; Oboh, H.A.; Anuonye, J.C.; Adebo, O.A. Evaluation of Fermented African Yam Bean Flour Composition and Influence of Substitution Levels on Properties of Wheat Bread. J. Food Sci. 2020, 85, 4281–4289. [Google Scholar] [CrossRef]
- Reyes-Moreno, C.; Cuevas-Rodríguez, E.O.; Milán-Carrillo, J.; Cárdenas-Valenzuela, O.G.; Barrón-Hoyos, J. Solid State Fermentation Process for Producing Chickpea (Cicer Arietinum L) Tempeh Flour. Physicochemical and Nutritional Characteristics of the Product. J. Sci. Food Agric. 2004, 84, 271–278. [Google Scholar] [CrossRef]
- Singhal, P.; Satya, S.; Naik, S.N. Fermented Bamboo Shoots: A Complete Nutritional, Anti-Nutritional and Antioxidant Profile of the Sustainable and Functional Food to Food Security. Food Chem. 2021, 3, 100041. [Google Scholar] [CrossRef]
- Moore, J.F.; DuVivier, R.; Johanningsmeier, S.D. Changes in the Free Amino Acid Profile of Pickling Cucumber during Lactic Acid Fermentation. J. Food Sci. 2022, 87, 599–611. [Google Scholar] [CrossRef]
- Lüttkopf, D.; Ballmer-Weber, B.K.; Wüthrich, B.; Vieths, S. Celery Allergens in Patients with Positive Double-Blind Placebo-Controlled Food Challenge. J. Allergy Clin. Immunol. 2000, 106, 390–399. [Google Scholar] [CrossRef]
- Xia, J.; Zu, Q.; Yang, A.; Wu, Z.; Li, X.; Tong, P.; Yuan, J.; Wu, Y.; Fan, Q.; Chen, H. Allergenicity Reduction and Rheology Property of Lactobacillus-Fermented Soymilk. J. Sci. Food Agric. 2019, 99, 6841–6849. [Google Scholar] [CrossRef]
- Rahim, M.H.A.; Hazrin-Chong, N.H.; Harith, H.H.; Wan-Mohtar, W.A.A.Q.I.; Sukor, R. Roles of Fermented Plant-, Dairy- and Meat-Based Foods in the Modulation of Allergic Responses. Food Sci. Hum. Wellness 2023, 12, 691–701. [Google Scholar] [CrossRef]
- Vici, G.; Belli, L.; Biondi, M.; Polzonetti, V. Gluten Free Diet and Nutrient Deficiencies: A Review. Clin. Nutr. 2016, 35, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Onwurafor, E.U.; Onweluzo, J.C.; Ezeoke, A.M. Effect of Fermentation Methods on Chemical and Microbial Properties of Mung Bean (Vigna Radiata) Flour. Niger. Food J. 2014, 32, 89–96. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Stockmann, R.; Ng, K.; Ajlouni, S. Revisiting Phytate-Element Interactions: Implications for Iron, Zinc and Calcium Bioavailability, with Emphasis on Legumes. Crit. Rev. Food Sci. Nutr. 2022, 62, 1696–1712. [Google Scholar] [CrossRef] [PubMed]
- Chawla, P.; Bhandari, L.; Sadh, P.K.; Kaushik, R. Impact of Solid-State Fermentation (Aspergillus Oryzae) on Functional Properties and Mineral Bioavailability of Black-Eyed Pea (Vigna Unguiculata) Seed Flour. Cereal Chem. 2017, 94, 437–442. [Google Scholar] [CrossRef]
- Song, H.S.; Lee, S.H.; Ahn, S.W.; Kim, J.Y.; Rhee, J.K.; Roh, S.W. Effects of the Main Ingredients of the Fermented Food, Kimchi, on Bacterial Composition and Metabolite Profile. Food Res. Int. 2021, 149, 110668. [Google Scholar] [CrossRef]
- Toydemir, G.; Gultekin Subasi, B.; Hall, R.D.; Beekwilder, J.; Boyacioglu, D.; Capanoglu, E. Effect of Food Processing on Antioxidants, Their Bioavailability and Potential Relevance to Human Health. Food Chem. 2022, 14, 100334. [Google Scholar] [CrossRef]
- Qiao, Y.; Zhang, K.; Zhang, Z.; Zhang, C.; Sun, Y.; Feng, Z. Fermented Soybean Foods: A Review of Their Functional Components, Mechanism of Action and Factors Influencing Their Health Benefits. Food Res. Int. 2022, 158, 111575. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health Benefits of Fruits and Vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Tarvainen, M.; Fabritius, M.; Yang, B. Determination of Vitamin K Composition of Fermented Food. Food Chem. 2019, 275, 515–522. [Google Scholar] [CrossRef]
- Granito, M.; Frias, J.; Doblado, R.; Guerra, M.; Champ, M.; Vidal-Valverde, C. Nutritional Improvement of Beans (Phaseolus Vulgaris) by Natural Fermentation. Eur. Food Res. Technol. 2001, 214, 226–231. [Google Scholar] [CrossRef]
- Doblado, R.; Frias, J.; Muñoz, R.; Vidal-Valverde, C. Fermentation of Vigna Sinensis Var. Carilla Flours by Natural Microflora and Lactobacillus Species. J. Food Prot. 2003, 66, 2313–2320. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Cai, W.X.; Xu, B.J. A Systematic Assesment on Vitamins (B2, B12) and GABA Profiles in Fermented Soy Products Marketed in China. J. Food Process. Preserv. 2017, 41, 13126. [Google Scholar] [CrossRef]
- Granito, M.; Torres, A.; Frias, J.; Guerra, M.; Vidal-Valverde, C. Influence of Fermentation on the Nutritional Value of Two Varieties of Vigna sinensis. Eur. Food Res. Technol. 2005, 220, 176–181. [Google Scholar] [CrossRef]
- Moa, H.; Kariluoto, S.; Piironen, V.; Zhu, Y.; Sanders, M.G.; Vincken, J.P.; Wolkers-Rooijackers, J.; Nout, M.J.R. Effect of Soybean Processing on Content and Bioaccessibility of Folate, Vitamin B12 and Isoflavones in Tofu and Tempeh. Food Chem. 2013, 141, 2418–2425. [Google Scholar] [CrossRef]
- Yanagisawa, Y.; Sumi, H. Natto Bacillus Contains a Large Amount of Water-Soluble Vitamin K (Menaquinone-7). J. Food Biochem. 2005, 29, 267–277. [Google Scholar] [CrossRef]
- Ifesan, B.O.T.; Egbewole, O.O.; Ifesan, B.T. Effect of Fermentation on Nutritional Composition of Selected Commonly Consumed Green Leafy Vegetables in Nigeria. Int. J. Appl. Sci. Biotechnol. 2014, 2, 291–297. [Google Scholar] [CrossRef]
- Frias, J.; Miranda, M.L.; Doblado, R.; Vidal-Valverde, C. Effect of Germination and Fermentation on the Antioxidant Vitamin Content and Antioxidant Capacity of Lupinus Albus L. Var. Multolupa. Food Chem. 2005, 92, 211–220. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health Benefits of Fermented Foods: Microbiota and Beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Palani, K.; Harbaum-Piayda, B.; Meske, D.; Keppler, J.K.; Bockelmann, W.; Heller, K.J.; Schwarz, K. Influence of Fermentation on Glucosinolates and Glucobrassicin Degradation Products in Sauerkraut. Food Chem. 2016, 190, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Peñas, E.; Frias, J.; Sidro, B.; Vidal-Valverde, C. Chemical Evaluation and Sensory Quality of Sauerkrauts Obtained by Natural and Induced Fermentations at Different NaCl Levels from Brassica Oleracea Var. Capitata Cv. Bronco Grown in Eastern Spain. Effect of Storage. J. Agric. Food Chem. 2010, 58, 3549–3557. [Google Scholar] [CrossRef] [PubMed]
- Ciska, E.; Honke, J.; Drabińska, N. Changes in Glucosinolates and Their Breakdown Products during the Fermentation of Cabbage and Prolonged Storage of Sauerkraut: Focus on Sauerkraut Juice. Food Chem. 2021, 365, 130498. [Google Scholar] [CrossRef]
- Gupta, P.; Wright, S.E.; Kim, S.H.; Srivastava, S.K. Phenethyl Isothiocyanate: A Comprehensive Review of Anti-Cancer Mechanisms. Biochem. Biophys. Acta 2014, 1846, 405–424. [Google Scholar] [CrossRef]
- Coscueta, E.R.; Sousa, A.S.; Reis, C.A.; Pintado, M.M. Phenylethyl Isothiocyanate: A Bioactive Agent for Gastrointestinal Health. Molecules 2022, 27, 794. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Peñas, E.; Sidro, B.; Ullate, M.; Frias, J.; Vidal-Valverde, C. White Cabbage Fermentation Improves Ascorbigen Content, Antioxidant and Nitric Oxide Production Inhibitory Activity in LPS-Induced Macrophages. LWT Food Sci. Technol. 2012, 46, 77–83. [Google Scholar] [CrossRef]
- Peñas, E.; Martinez-Villaluenga, C.; Frias, J.; Sánchez-Martínez, M.J.; Pérez-Corona, M.T.; Madrid, Y.; Cámara, C.; Vidal-Valverde, C. Se Improves Indole Glucosinolate Hydrolysis Products Content, Se-Methylselenocysteine Content, Antioxidant Capacity and Potential Anti-Inflammatory Properties of Sauerkraut. Food Chem. 2012, 132, 907–914. [Google Scholar] [CrossRef]
- Chien, H.L.; Huang, H.Y.; Chou, C.C. Transformation of Isoflavone Phytoestrogens during the Fermentation of Soymilk with Lactic Acid Bacteria and Bifidobacteria. Food Microbiol. 2006, 23, 772–778. [Google Scholar] [CrossRef]
- Ricci, A.; Marrella, M.; Hadj Saadoun, J.; Bernini, V.; Godani, F.; Dameno, F.; Neviani, E.; Lazzi, C. Development of Lactic Acid-Fermented Tomato Products. Microorganisms 2020, 8, 1192. [Google Scholar] [CrossRef]
- Dhull, S.B.; Punia, S.; Kidwai, M.K.; Kaur, M.; Chawla, P.; Purewal, S.S.; Sangwan, M.; Palthania, S. Solid-State Fermentation of Lentil (Lens Culinaris L.) with Aspergillus Awamori: Effect on Phenolic Compounds, Mineral Content, and Their Bioavailability. Legume Sci. 2020, 2, 37. [Google Scholar] [CrossRef]
- Sawicki, T.; Wiczkowski, W. The Effects of Boiling and Fermentation on Betalain Profiles and Antioxidant Capacities of Red Beetroot Products. Food Chem. 2018, 259, 292–303. [Google Scholar] [CrossRef]
- Xiao, F.; Xu, T.; Lu, B.; Liu, R. Guidelines for Antioxidant Assays for Food Components. Food Front. 2020, 1, 60–69. [Google Scholar] [CrossRef]
- Negro, E.J.; Sendker, J.; Stark, T.; Lipowicz, B.; Hensel, A. Phytochemical and Functional Analysis of Horseradish (Armoracia Rusticana) Fermented and Non-Fermented Root Extracts. Fitoterapia 2022, 162, 105282. [Google Scholar] [CrossRef]
- Liu, S.; Jia, M.; Chen, J.; Wan, H.; Dong, R.; Nie, S.; Xie, M.; Yu, Q. Removal of Bound Polyphenols and Its Effect on Antioxidant and Prebiotics Properties of Carrot Dietary Fiber. Food Hydrocoll. 2019, 93, 284–292. [Google Scholar] [CrossRef]
- Ye, J.H.; Huang, L.Y.; Terefe, N.S.; Augustin, M.A. Fermentation-Based Biotransformation of Glucosinolates, Phenolics and Sugars in Retorted Broccoli Puree by Lactic Acid Bacteria. Food Chem. 2019, 286, 616–623. [Google Scholar] [CrossRef]
- Thinh Nguyen, V.P.; Stewart, J.; Lopez, M.; Ioannou, I.; Allais, F. Glucosinolates: Natural Occurrence, Biosynthesis, Accessibility, Isolation, Structures, and Biological Activities. Molecules 2020, 25, 4537. [Google Scholar] [CrossRef]
- Fideler, J.; Johanningsmeier, S.D.; Ekelöf, M.; Muddiman, D.C. Discovery and Quantification of Bioactive Peptides in Fermented Cucumber by Direct Analysis IR-MALDESI Mass Spectrometry and LC-QQQ-MS. Food Chem. 2019, 271, 715–723. [Google Scholar] [CrossRef]
- Tlais, A.Z.A.; Junior, W.J.F.L.; Filannino, P.; Campanaro, S.; Gobbetti, M.; Cagno, R. di How Microbiome Composition Correlates with Biochemical Changes during Sauerkraut Fermentation: A Focus on Neglected Bacterial Players and Functionalities. Microbiol. Spectr. 2022, 10, 0016822. [Google Scholar] [CrossRef]
- Xiao, Y.; Sun, M.; Zhang, Q.; Chen, Y.; Miao, J.; Rui, X.; Dong, M. Effects of Cordyceps militaris (L.) Fr. Fermentation on the Nutritional, Physicochemical, Functional Properties and Angiotensin I Converting Enzyme Inhibitory Activity of Red Bean (Phaseolus angularis [Willd.] W.F. Wight.) Flour. J. Food Sci. Technol. 2018, 55, 1244–1255. [Google Scholar] [CrossRef]
- Sozer, N.; Melama, L.; Silbir, S.; Rizzello, C.G.; Flander, L.; Poutanen, K. Lactic Acid Fermentation as a Pre-Treatment Process for Faba Bean Flour and Its Effect on Textural, Structural and Nutritional Properties of Protein-Enriched Gluten-Free Faba Bean Breads. Foods 2019, 8, 431. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, M.; Matsui, T.; Yano, H. Fermentation of Soybean Flour with Aspergillus Usamii Improves Availabilities of Zinc and Iron in Rats. J. Nutr. Sci. Vitaminol. 1998, 44, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Ogawa, Y. In Vitro Protein Digestibility and Biochemical Characteristics of Soaked, Boiled and Fermented Soybeans. Sci. Rep. 2021, 11, 14257. [Google Scholar] [CrossRef] [PubMed]
- Matchar, D.B.; McCrory, D.C.; Orlando, L.A.; Patel, M.R.; Patel, U.D.; Patwardhan, M.B.; Powers, B.; Samsa, G.P.; Gray, R.N. Systematic Review: Comparative Effectiveness of Angiotensin-Converting Enzyme Inhibitors and Angiotensin II Receptor Blockers for Treating Essential Hypertension. Ann. Intern. Med. 2008, 148, 16–29. [Google Scholar] [CrossRef]
- Wang, G.M.; Li, L.J.; Tang, W.L.; Wright, J.M. Renin Inhibitors versus Angiotensin Converting Enzyme (ACE) Inhibitors for Primary Hypertension. Cochrane Database Syst. Rev. 2020, 10, CD012569. [Google Scholar] [CrossRef] [PubMed]
- Momma, K. ACE Inhibitors in Pediatric Patients with Heart Failure. Paediatr. Drugs 2006, 8, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Lee, S.H.; Jung, J.Y.; Choi, E.J.; Jeon, C.O. Microbial Succession and Metabolite Changes during Long-Term Storage of Kimchi. J. Food Sci. 2013, 78, 63–69. [Google Scholar] [CrossRef]
- Lee, M.E.; Jang, J.Y.; Lee, J.H.; Park, H.W.; Choi, H.J.; Kim, T.W. Starter Cultures for Kimchi Fermentation. J. Microbiol. Biotechnol. 2015, 25, 559–568. [Google Scholar] [CrossRef]
- Fideler, M.J. Generation of Bioactive Peptides and γ-Aminobutyric Acid (GABA) During Natural Lactic Acid Fermentation of Cucumber; North Carolina State University ProQuest Dissertations Publishing: Raleigh, NC, USA, 2021. [Google Scholar]
- Möhler, H. The GABA System in Anxiety and Depression and Its Therapeutic Potential. Neuropharmacology 2012, 62, 42–53. [Google Scholar] [CrossRef]
- Ramos, I.M.; Poveda, J.M. Fermented Sheep’s Milk Enriched in Gamma-Amino Butyric Acid (GABA) by the Addition of Lactobacilli Strains Isolated from Different Food Environments. LWT 2022, 163, 113581. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef]
- Donkor, O.N.; Shah, N.P. Production of Beta-Glucosidase and Hydrolysis of Isoflavone Phytoestrogens by Lactobacillus Acidophilus, Bifidobacterium Lactis, and Lactobacillus Casei in Soymilk. J. Food Sci. 2008, 73, 15–20. [Google Scholar] [CrossRef]
- Kingston, J.J.; Radhika, M.; Roshini, P.T.; Raksha, M.A.; Murali, H.S.; Batra, H.V. Molecular Characterization of Lactic Acid Bacteria Recovered from Natural Fermentation of Beet Root and Carrot Kanji. Indian J. Microbiol. 2010, 50, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Kaszab, E.; Laczkó, L.; Bali, K.; Fidrus, E.; Bányai, K.; Kardos, G. Draft Genome Sequences of Lacticaseibacillus Rhamnosus Cek-R1, Lacticaseibacillus Paracasei Cek-R2, and Lentilactobacillus Otakiensis Cek-R3, Isolated from a Beetroot Product. Microbiol. Resour. Announc. 2022, 11, 0092121. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Park, J.Y.; Lee, J.E.; Kim, K.T.; Paik, H.D. Antioxidant Activity and Inhibitory Effect on Nitric Oxide Production of Hydroponic Ginseng Fermented with Lactococcus Lactis KC24. Antioxidants 2021, 10, 1614. [Google Scholar] [CrossRef] [PubMed]
- Ge, B.; Jiang, P.; Han, F.; Saleh, N.K.; Dhiman, N.; Fedorko, D.P.; Nelson, N.A.; Meng, J. Identification and Antimicrobial Susceptibility of Lactic Acid Bacteria from Retail Fermented Foods. J. Food Prot. 2007, 70, 2606–2612. [Google Scholar] [CrossRef]
- Ehrmann, M.; Ludwig, W.; Schleifer, K.H. Reverse Dot Blot Hybridization: A Useful Method for the Direct Identification of Lactic Acid Bacteria in Fermented Food. FEMS Microbiol. Lett. 1994, 117, 143–149. [Google Scholar] [CrossRef]
- Nielsen, E.S.; Garnås, E.; Jensen, K.J.; Hansen, L.H.; Olsen, P.S.; Ritz, C.; Krych, L.; Nielsen, D.S. Lacto-Fermented Sauerkraut Improves Symptoms in IBS Patients Independent of Product Pasteurisation—A Pilot Study. Food Funct. 2018, 9, 5323–5335. [Google Scholar] [CrossRef]
- Yang, Y.; Deng, Y.; Jin, Y.; Liu, Y.; Xia, B.; Sun, Q. Dynamics of Microbial Community during the Extremely Long-Term Fermentation Process of a Traditional Soy Sauce. J. Sci. Food Agric. 2017, 97, 3220–3227. [Google Scholar] [CrossRef]
- Yang, X.; Hu, W.; Xiu, Z.; Jiang, A.; Yang, X.; Saren, G.; Ji, Y.; Guan, Y.; Feng, K. Microbial Community Dynamics and Metabolome Changes During Spontaneous Fermentation of Northeast Sauerkraut from Different Households. Front. Microbiol. 2020, 11, 1878. [Google Scholar] [CrossRef]
- Du, R.; Ge, J.; Zhao, D.; Sun, J.; Ping, W.; Song, G. Bacterial Diversity and Community Structure during Fermentation of Chinese Sauerkraut with Lactobacillus Casei 11MZ-5-1 by Illumina Miseq Sequencing. Lett. Appl. Microbiol. 2018, 66, 55–62. [Google Scholar] [CrossRef]
- Niksic, M.; Niebuhr, S.E.; Dickson, J.S.; Mendonca, A.F.; Koziczkowski, J.J.; Ellingson, J.L.E. Survival of Listeria Monocytogenes and Escherichia Coli O157:H7 during Sauerkraut Fermentation. J. Food Prot. 2005, 68, 1367–1374. [Google Scholar] [CrossRef]
- Lee, S.H.; Whon, T.W.; Roh, S.W.; Jeon, C.O. Unraveling Microbial Fermentation Features in Kimchi: From Classical to Meta-Omics Approaches. Appl. Microbiol. Biotechnol. 2020, 104, 7731–7744. [Google Scholar] [CrossRef] [PubMed]
- Breidt, F.; Caldwell, J.M. Survival of Escherichia Coli O157:H7 in Cucumber Fermentation Brines. J. Food Sci. 2011, 76, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Page, C.A.; Pérez-Díaz, I.M. Whole-Genome Sequencing and Annotation of Selected Lactobacillales Isolated from Commercial Cucumber Fermentation. Microbiol. Resour. Announc. 2021, 10, 0062521. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Pérez-Díaz, I.M. Contribution of Leuconostocaceae to CO2-Mediated Bloater Defect in Cucumber Fermentation. Food Microbiol. 2020, 91, 103536. [Google Scholar] [CrossRef] [PubMed]
- Franco, W.; Pérez-Díaz, I.M. Microbial Interactions Associated with Secondary Cucumber Fermentation. J. Appl. Microbiol. 2013, 114, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.H.; Green-Johnson, J.M.; Buckley, N.D.; Lin, Q.Y. Bioactivity of Soy-Based Fermented Foods: A Review. Biotechnol. Adv. 2019, 37, 223–238. [Google Scholar] [CrossRef]
- Zhao, G.; Li, J.; Zheng, F.; Yao, Y. The Fermentation Properties and Microbial Diversity of Soy Sauce Fermented by Germinated Soybean. J. Sci. Food Agric. 2021, 101, 2920–2929. [Google Scholar] [CrossRef]
- Aljahani, A.H. Microbiological and Physicochemical Quality of Vegetable Pickles. J. Saudi Soc. Agric. Sci. 2020, 19, 415–421. [Google Scholar] [CrossRef]
- Liu, N.; Deng, X.J.; Liang, C.Y.; Cai, H.Y. Fermented Broccoli Residue Reduced Harmful Bacterial Loads and Improved Meat Antioxidation of Free-Range Broilers. J. Appl. Poult. Res. 2018, 27, 590–596. [Google Scholar] [CrossRef]
- Li, Y.; Ten, M.M.Z.; Zwe, Y.H.; Li, D. Lactiplantibacillus Plantarum 299v as Starter Culture Suppresses Enterobacteriaceae More Efficiently than Spontaneous Fermentation of Carrots. Food. Microbiol. 2022, 103, 103952. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Lee, N.K.; Paik, H.D. Health Benefits of Lactic Acid Bacteria Isolated from Kimchi, with Respect to Immunomodulatory Effects. Food Sci. Biotechnol. 2015, 24, 783–789. [Google Scholar] [CrossRef]
- Raghuvanshi, R.; Grayson, A.G.; Schena, I.; Amanze, O.; Suwintono, K.; Quinn, R.A. Microbial Transformations of Organically Fermented Foods. Metabolites 2019, 9, 165. [Google Scholar] [CrossRef]
- Moreno, M.R.F.; Leisner, J.J.; Tee, L.K.; Ley, C.; Radu, S.; Rusul, G.; Vancanneyt, M.; de Vuyst, L. Microbial Analysis of Malaysian Tempeh, and Characterization of Two Bacteriocins Produced by Isolates of Enterococcus Faecium. J. Appl. Microbiol. 2002, 92, 147–157. [Google Scholar] [CrossRef]
- Aoki, H.; Uda, I.; Tagami, K.; Furuya, Y.; Endo, Y.; Fujimoto, K. The Production of a New Tempeh-like Fermented Soybean Containing a High Level of Gamma-Aminobutyric Acid by Anaerobic Incubation with Rhizopus. Biosci. Biotechnol. Biochem. 2003, 67, 1018–1023. [Google Scholar] [CrossRef]
- Kil, J.-H.; Jung, K.-O.; Lee, H.-S.; Hwang, I.-K.; Kim, Y.-J.; Park, K.-Y. Effects of Kimchi on Stomach and Colon Health of Helicobacter Pylori-Infected Volunteers. Prev. Nutr. Food Sci. 2004, 9, 161–166. [Google Scholar] [CrossRef]
- Fujisawa, T.; Shinohara, K.; Kishimoto, Y.; Terada, A. Effect of Miso Soup Containing Natto on the Composition and Metabolic Activity of the Human Faecal Flora. Microb. Ecol. Health Dis. 2009, 18, 79–84. [Google Scholar] [CrossRef]
- Feng, J.; Liu, P.; Yang, X.; Zhao, X. Screening of Immunomodulatory and Adhesive Lactobacillus with Antagonistic Activities against Salmonella from Fermented Vegetables. World J. Microbiol. Biotechnol. 2015, 31, 1947–1954. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, X.; Li, S.; Li, C.; Li, D.; Yang, Z. Evaluation of Probiotic Properties of Lactobacillus Plantarum Strains Isolated from Chinese Sauerkraut. World J. Microbiol. Biotechnol. 2013, 29, 489–498. [Google Scholar] [CrossRef]
- Venema, K.; Verhoeven, J.; Beckman, C.; Keller, D. Survival of a Probiotic-Containing Product Using Capsule-within-Capsule Technology in an in Vitro Model of the Stomach and Small Intestine (TIM-1). Benef. Microbes 2020, 11, 403–409. [Google Scholar] [CrossRef]
- Bernatek, M.; Żukiewicz-Sobczak, W.; Lachowicz-Wiśniewska, S.; Piątek, J. Factors Determining Effective Probiotic Activity: Evaluation of Survival and Antibacterial Activity of Selected Probiotic Products Using an “In Vitro” Study. Nutrients 2022, 14, 3323. [Google Scholar] [CrossRef] [PubMed]
- Schuermann, L.E.; Bergmann, C.B.; Goetzman, H.; Caldwell, C.C.; Satish, L. Burns, 2022; in press. [CrossRef]
- Li, H.; Peng, F.; Lin, J.-X.; Xiong, T.; Huang, T. Preparation of Probiotic Microcapsules Using Gelatin-Xylooligosaccharides Conjugates by Spray Drying: Physicochemical Properties, Survival, Digestion Resistance and Colonization. Food Biosci. 2023, 52, 102462. [Google Scholar] [CrossRef]
- Champagne, C.P.; Guertin, N.; Raymond, Y. Strategies to Improve the Survival of Probiotic Lacticaseibacillus Rhamnosus R0011 during the Production and Storage of Granola Bars. Can. J. Microbiol. 2022, 68, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health Benefits of Fermented Foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef]
- Kwak, S.-H.; Cho, Y.-M.; Noh, G.-M.; Om, A.-S. Cancer Preventive Potential of Kimchi Lactic Acid Bacteria (Weissella Cibaria, Lactobacillus Plantarum). J. Cancer Prev. 2014, 19, 253. [Google Scholar] [CrossRef]
- Kocot, A.M.; Wróblewska, B. Fermented Products and Bioactive Food Compounds as a Tool to Activate Autophagy and Promote the Maintenance of the Intestinal Barrier Function. Trends Food Sci. Technol. 2021, 118, 905–919. [Google Scholar] [CrossRef]
- Schoen, C.; Schulz, A.; Schweikart, J.; Schütt, S.; von Baehr, V. Regulatory Effects of a Fermented Food Concentrate on Immune Function Parameters in Healthy Volunteers. Nutrition 2009, 25, 499–505. [Google Scholar] [CrossRef]
- Lin, Q.; Mathieu, O.; Tompkins, T.A.; Buckley, N.D.; Green-Johnson, J.M. Modulation of the TNFα-Induced Gene Expression Profile of Intestinal Epithelial Cells by Soy Fermented with Lactic Acid Bacteria. J. Funct. Foods 2016, 23, 400–411. [Google Scholar] [CrossRef]
- Won, T.J.; Kim, B.; Song, D.S.; Lim, Y.T.; Oh, E.S.; Lee, D.I.; Park, E.S.; Min, H.; Park, S.Y.; Hwang, K.W. Modulation of Th1/Th2 Balance by Lactobacillus Strains Isolated from Kimchi via Stimulation of Macrophage Cell Line J774A.1 In Vitro. J. Food Sci. 2011, 76, 55–61. [Google Scholar] [CrossRef]
- Oba, M.; Rongduo, W.; Saito, A.; Okabayashi, T.; Yokota, T.; Yasuoka, J.; Sato, Y.; Nishifuji, K.; Wake, H.; Nibu, Y.; et al. Natto Extract, a Japanese Fermented Soybean Food, Directly Inhibits Viral Infections Including SARS-CoV-2 in Vitro. Biochem. Biophys. Res. Commun. 2021, 570, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary Polyphenols and the Prevention of Diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- An, J.M.; Kang, E.A.; Han, Y.M.; Oh, J.Y.; Lee, D.Y.; Choi, S.H.; Kim, D.H.; Hahm, K.B. Dietary Intake of Probiotic Kimchi Ameliorated IL-6-Driven Cancer Cachexia. J. Clin. Biochem. Nutr. 2019, 65, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Albarracin, S.L.; Stab, B.; Casas, Z.; Sutachan, J.J.; Samudio, I.; Gonzalez, J.; Gonzalo, L.; Capani, F.; Morales, L.; Barreto, G.E. Effects of Natural Antioxidants in Neurodegenerative Disease. Nutr. Neurosci. 2012, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Park, K.Y.; Kim, H.Y.; Ahn, S.C.; Cho, E.J. Anti-Aging Effects and Mechanisms of Kimchi during Fermentation under Stress-Induced Premature Senescence Cellular System. Food Sci. Biotechnol. 2011, 20, 643–649. [Google Scholar] [CrossRef]
- Dash, S.; Clarke, G.; Berk, M.; Jacka, F.N. The Gut Microbiome and Diet in Psychiatry: Focus on Depression. Curr. Opin. Psychiatry 2015, 28, 1–6. [Google Scholar] [CrossRef]
- Aslam, H.; Green, J.; Jacka, F.N.; Collier, F.; Berk, M.; Pasco, J.; Dawson, S.L. Fermented Foods, the Gut and Mental Health: A Mechanistic Overview with Implications for Depression and Anxiety. Nutr. Neurosci. 2020, 23, 659–671. [Google Scholar] [CrossRef]
- Seppo, L.; Jauhiainen, T.; Poussa, T.; Korpela, R. A Fermented Milk High in Bioactive Peptides Has a Blood Pressure-Lowering Effect in Hypertensive Subjects. Am. J. Clin. Nutr. 2003, 77, 326–330. [Google Scholar] [CrossRef]
- An, S.Y.; Lee, M.S.; Jeon, J.Y.; Ha, E.S.; Kim, T.H.; Yoon, J.Y.; Ok, C.O.; Lee, H.K.; Hwang, W.S.; Choe, S.J.; et al. Beneficial Effects of Fresh and Fermented Kimchi in Prediabetic Individuals. Ann. Nutr. Metab. 2013, 63, 111–119. [Google Scholar] [CrossRef]
- Hsu, Y.-H.; Chen, L.-G.; Chu, C.; Lamponi, S.; Hariyanto, I.; Hsieh, C.-W.; Hsu, Y.-H.; Chen, L.-G.; Chu, C.; Bor-Chun Weng, B. In Vitro and In Vivo Assessments of Anti-Hyperglycemic Properties of Soybean Residue Fermented with Rhizopus Oligosporus and Lactiplantibacillus Plantarum. 2022, 12, 1716. 12. [CrossRef]
- Huang, Y.C.; Wu, B.H.; Chu, Y.L.; Chang, W.C.; Wu, M.C. Effects of Tempeh Fermentation with Lactobacillus Plantarum and Rhizopus Oligosporus on Streptozotocin-Induced Type II Diabetes Mellitus in Rats. Nutrients 2018, 10, 1143. [Google Scholar] [CrossRef] [PubMed]
- Byun, M.S.; Yu, O.K.; Cha, Y.S.; Park, T.S. Korean Traditional Chungkookjang Improves Body Composition, Lipid Profiles and Atherogenic Indices in Overweight/Obese Subjects: A Double-Blind, Randomized, Crossover, Placebo-Controlled Clinical Trial. Eur. J. Clin. Nutr. 2016, 70, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Kondo, K.; Matsumoto, Y.; Zhao, B.Q.; Otsuguro, K.; Maeda, T.; Tsukamoto, Y.; Urano, T.; Umemura, K. Dietary Supplementation of Fermented Soybean, Natto, Suppresses Intimal Thickening and Modulates the Lysis of Mural Thrombi after Endothelial Injury in Rat Femoral Artery. Life Sci. 2003, 73, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Iki, M.; Morita, A.; Kajita, E.; Kagamimori, S.; Kagawa, Y.; Yoneshima, H. Intake of Fermented Soybeans, Natto, Is Associated with Reduced Bone Loss in Postmenopausal Women: Japanese Population-Based Osteoporosis (JPOS) Study. J. Nutr. 2006, 136, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, D.S.; Kapanoor, S.S.; Girigouda, K.; Kote, N.V.; Mulimani, V.H. Reduction of Flatus-Inducing Factors in Soymilk by Immobilized α-Galactosidase. Biotechnol. Appl. Biochem. 2006, 45, 51–57. [Google Scholar] [CrossRef]
- Mi, T.; Wang, D.; Yao, S.; Yang, H.; Che, Y.; Wu, C. Effects of Salt Concentration on the Quality and Microbial Diversity of Spontaneously Fermented Radish Paocai. Food Res. Int. 2022, 160, 111622. [Google Scholar] [CrossRef]
- Tekol, Y. Irreversible and Reversible Components in the Genesis of Hypertension by Sodium Chloride (Salt). Med. Hypotheses 2008, 70, 255–259. [Google Scholar] [CrossRef]
- Xiang, R.P.; Sun, W.D.; Zhang, K.C.; Li, J.C.; Wang, J.Y.; Wang, X.L. Sodium Chloride-Induced Acute and Chronic Pulmonary Hypertension Syndrome in Broiler Chickens. Poultry Sci. 2004, 83, 732–736. [Google Scholar] [CrossRef]
- World Health Organization. Report of the Formal Meeting of Member States to Conclude the Work on the Comprehensive Global Monitoring Framework, Including Indicators, and a Set of Voluntary Global Targets for the Prevention and Control of Noncommunicable Diseases; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Song, H.J.; Park, S.J.; Jang, D.J.; Kwon, D.Y.; Lee, H.J. High Consumption of Salt-Fermented Vegetables and Hypertension Risk in Adults: A 12-Year Follow-up Study. Asia Pac. J. Clin. Nutr. 2017, 26, 698–707. [Google Scholar] [CrossRef]
- Song, H.J.; Lee, H.-J. Consumption of Kimchi, a Salt Fermented Vegetable, Is Not Associated with Hypertension Prevalence. J. Ethn. Foods 2014, 1, 8–12. [Google Scholar] [CrossRef]
- Diez-Ozaeta, I.; Astiazaran, O.J. Fermented Foods: An Update on Evidence-Based Health Benefits and Future Perspectives. Food Res. Int. 2022, 156, 111133. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.; Wang, Y.N.; Zhang, C.M.; Kang, S.G. In Vitro and in Vivo Antihypertensive and Antioxidant Activities of Fermented Roots of Allium Hookeri. Chin. Herb. Med. 2021, 13, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.M.; Moran, A.E.; Whelton, P.K. Treatment of Hypertension: A Review. JAMA 2022, 328, 1849–1861. [Google Scholar] [CrossRef]
- Greer, R.C.; Marklund, M.; Anderson, C.A.M.; Cobb, L.K.; Dalcin, A.T.; Henry, M.; Appel, L.J. Potassium-Enriched Salt Substitutes as a Means to Lower Blood Pressure: Benefits and Risks. Hypertension 2020, 75, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.E.; Appel, L.J. Potassium-Enriched Salt Substitutes: Benefits, Risks, and a “Trolley Problem” in Public Health. Am. J. Clin. Nutr. 2021, 114, 12–13. [Google Scholar] [CrossRef] [PubMed]
- Wieërs, M.L.A.J.; Mulder, J.; Rotmans, J.I.; Hoorn, E.J. Potassium and the Kidney: A Reciprocal Relationship with Clinical Relevance. Pediatr. Nephrol. 2022, 37, 2245–2254. [Google Scholar] [CrossRef]



| Vegetables | Proteins | Carbohydrates | Fibre | Total Fats | SFA | MUFA | PUFA |
|---|---|---|---|---|---|---|---|
| Beetroot | 1.61 | 9.56 | 2.8 | 0.17 | 0.027 | 0.032 | 0.06 |
| Beetroot, fermented | 0.8 | 16.3 | 0.8 | 0.08 | 0.013 | 0.016 | 0.029 |
| Chickpea, canned, no added fat | 8.19 | 22.4 | 7.3 | 3.24 | 0.339 | 0.739 | 1.47 |
| Soybean, cooked | 16.9 | 7.77 | 5.6 | 14.8 | 2.1 | 4.57 | 7.39 |
| Natto | 19.4 | 12.7 | 5.4 | 11 | 1.59 | 2.43 | 6.21 |
| Red cabbage | 1.43 | 7.37 | 2.1 | 0.16 | 0.021 | 0.012 | 0.08 |
| Red cabbage, pickled | 1.19 | 11 | 1.7 | 0.15 | 0.017 | 0.01 | 0.066 |
| Lentils, cooked | 8.38 | 18.7 | 7.3 | 6.86 | 0.939 | 2.78 | 2.84 |
| Fava beans, cooked | 7.06 | 18.3 | 5 | 6.88 | 0.951 | 2.8 | 2.83 |
| Vegetables | Ca [mg/100 g] | Fe [mg/100 g] | Mg [mg/100 g] | Zn [mg/100 g] | Vitamin C [mg/100 g] | Riboflavin [mg/100 g] | Folates, Total [µg/100 g] |
|---|---|---|---|---|---|---|---|
| Beetroot | 16 | 0.8 | 23 | 0.35 | 4.0 | 0.04 | 109 |
| Beetroot, pickled | 11 | 0,41 | 15 | 0.26 | 2.3 | 0.048 | 27 |
| Green cabbage | 40 | 0.47 | 12 | 0.18 | 36.6 | 0.04 | 43 |
| Sauerkraut, Japanese style | 35 | 0.39 | 10 | 0.15 | 25.8 | 0.032 | 30 |
| Radish | 25 | 0.34 | 10 | 0.28 | 14.8 | 0.039 | 25 |
| Radish, pickled, Hawaiian style | 22 | 0.29 | 8 | 0.23 | 10.4 | 0.032 | 18 |
| Carrot | 33 | 0.3 | 12 | 0.24 | 5.9 | 0.058 | 19 |
| Cucumber | 16 | 0.28 | 13 | 0.2 | 8 | 0.033 | 7 |
| Cucumber, pickled | 54 | 0.23 | 7.1 | 0.11 | 2.1 | 0.057 | 8 |
| Parsley, root | 138 | 6.2 | 27 | 1.07 | 45 | 0.086 | 180 |
| Red cabbage | 45 | 0.8 | 16 | 0.22 | 57 | 0.069 | 18 |
| Red cabbage, pickled | 39 | 0.67 | 13 | 0.18 | 40.2 | 0.055 | 13 |
| Green tomato, raw | 13 | 0.51 | 10 | 0.07 | 23.4 | 0.04 | 9 |
| Green tomato, pickled | 12 | 0.43 | 8 | 0.06 | 18.4 | 0.032 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knez, E.; Kadac-Czapska, K.; Grembecka, M. Effect of Fermentation on the Nutritional Quality of the Selected Vegetables and Legumes and Their Health Effects. Life 2023, 13, 655. https://doi.org/10.3390/life13030655
Knez E, Kadac-Czapska K, Grembecka M. Effect of Fermentation on the Nutritional Quality of the Selected Vegetables and Legumes and Their Health Effects. Life. 2023; 13(3):655. https://doi.org/10.3390/life13030655
Chicago/Turabian StyleKnez, Eliza, Kornelia Kadac-Czapska, and Małgorzata Grembecka. 2023. "Effect of Fermentation on the Nutritional Quality of the Selected Vegetables and Legumes and Their Health Effects" Life 13, no. 3: 655. https://doi.org/10.3390/life13030655