
Effects of Temperature and Humidity on Soil Gross Nitrogen Transformation in a Typical Shrub Ecosystem in Yanshan Mountain and Hilly Region

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
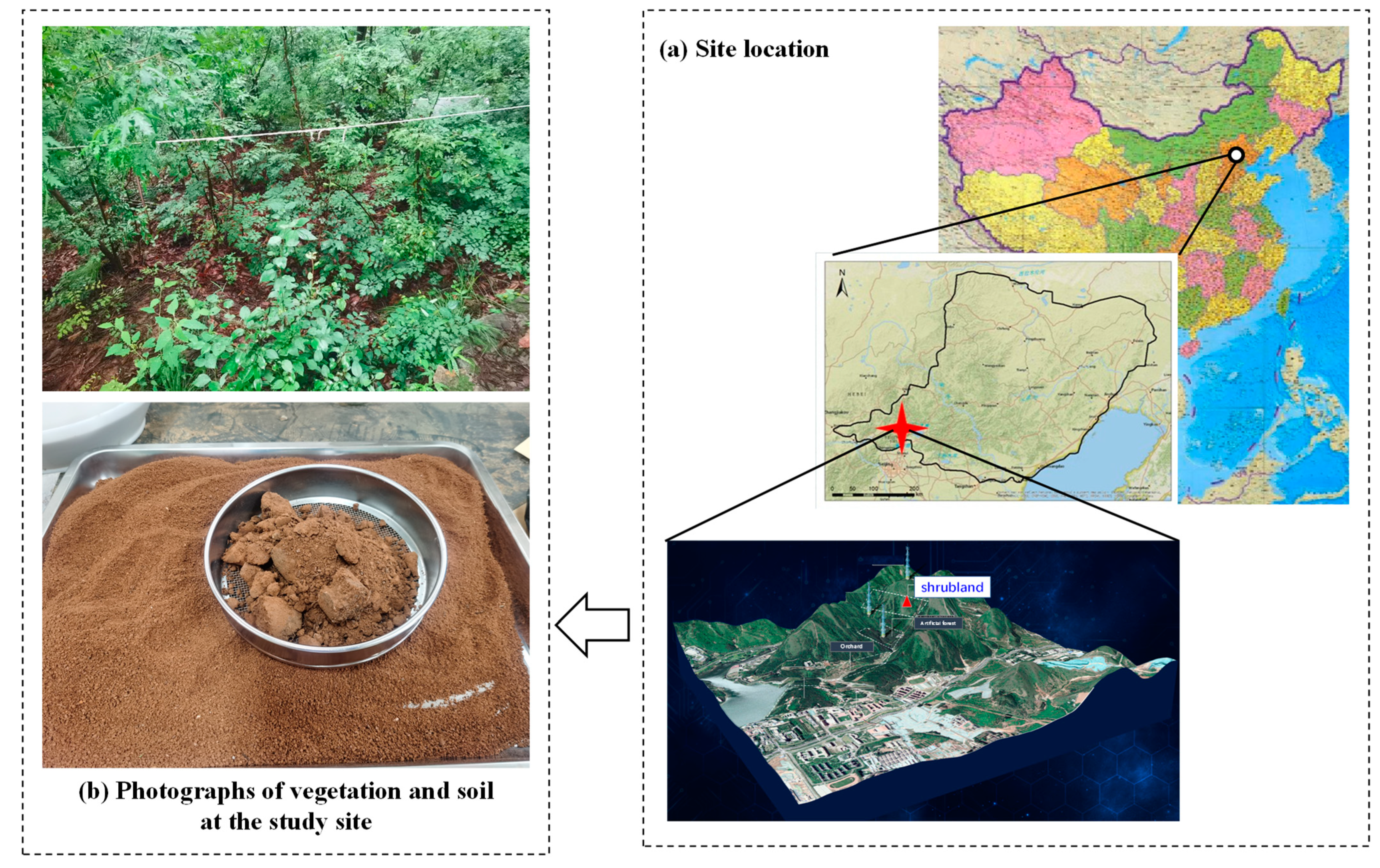
2.1. Study Site Description
2.2. Soil Sampling and Experimental Design
2.3. Measurements of Gross N Transformation Rates
2.4. Measurements of Soil Properties
2.5. Data Processing and Statistical Analysis Methods
3. Results
3.1. Soil Properties
3.2. Changes of 15N Enrichments and Soil Inorganic N Concentrations during the Incubation Period
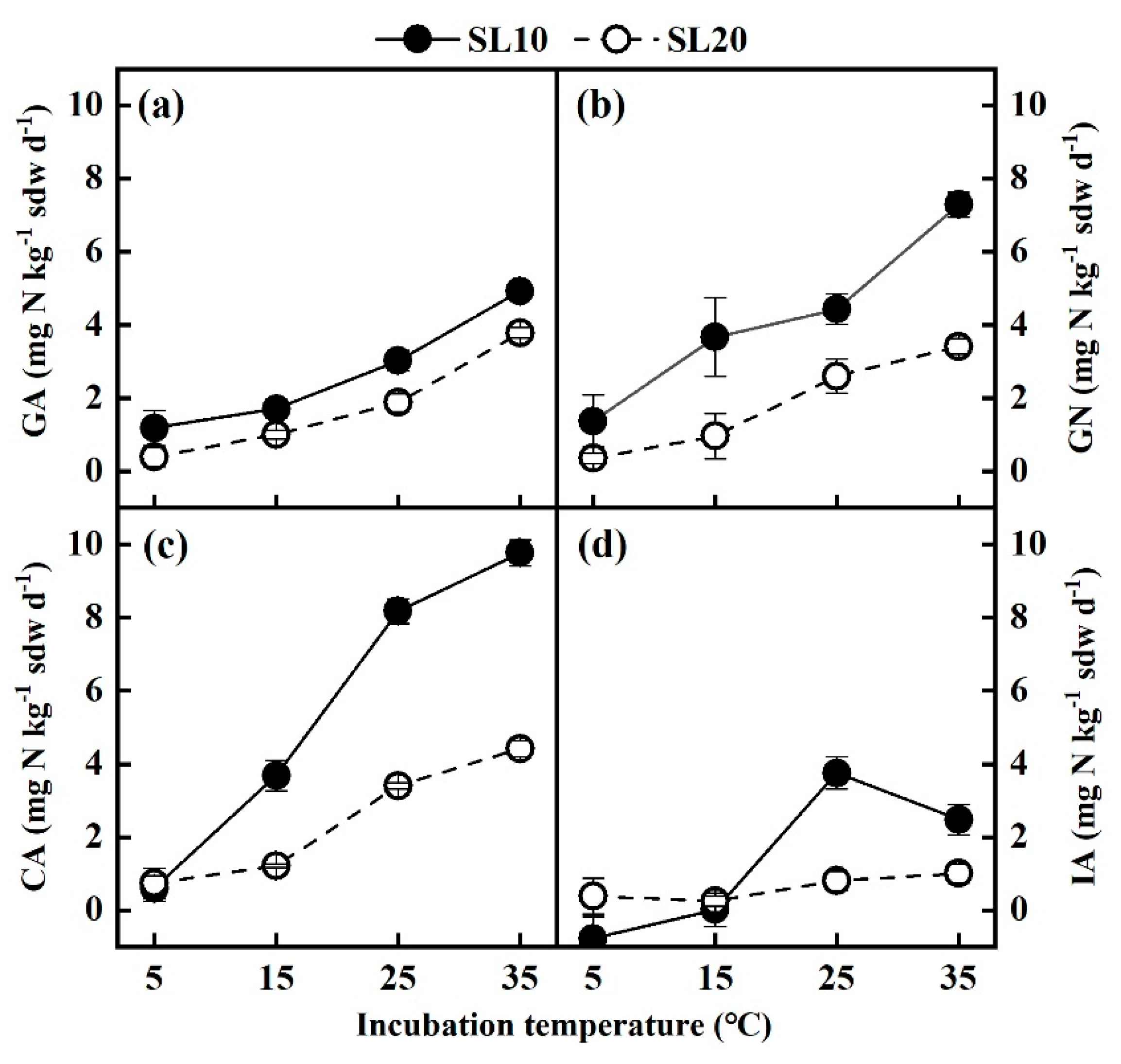
3.3. Gross N Transformation Rates under Different Incubation Temperatures
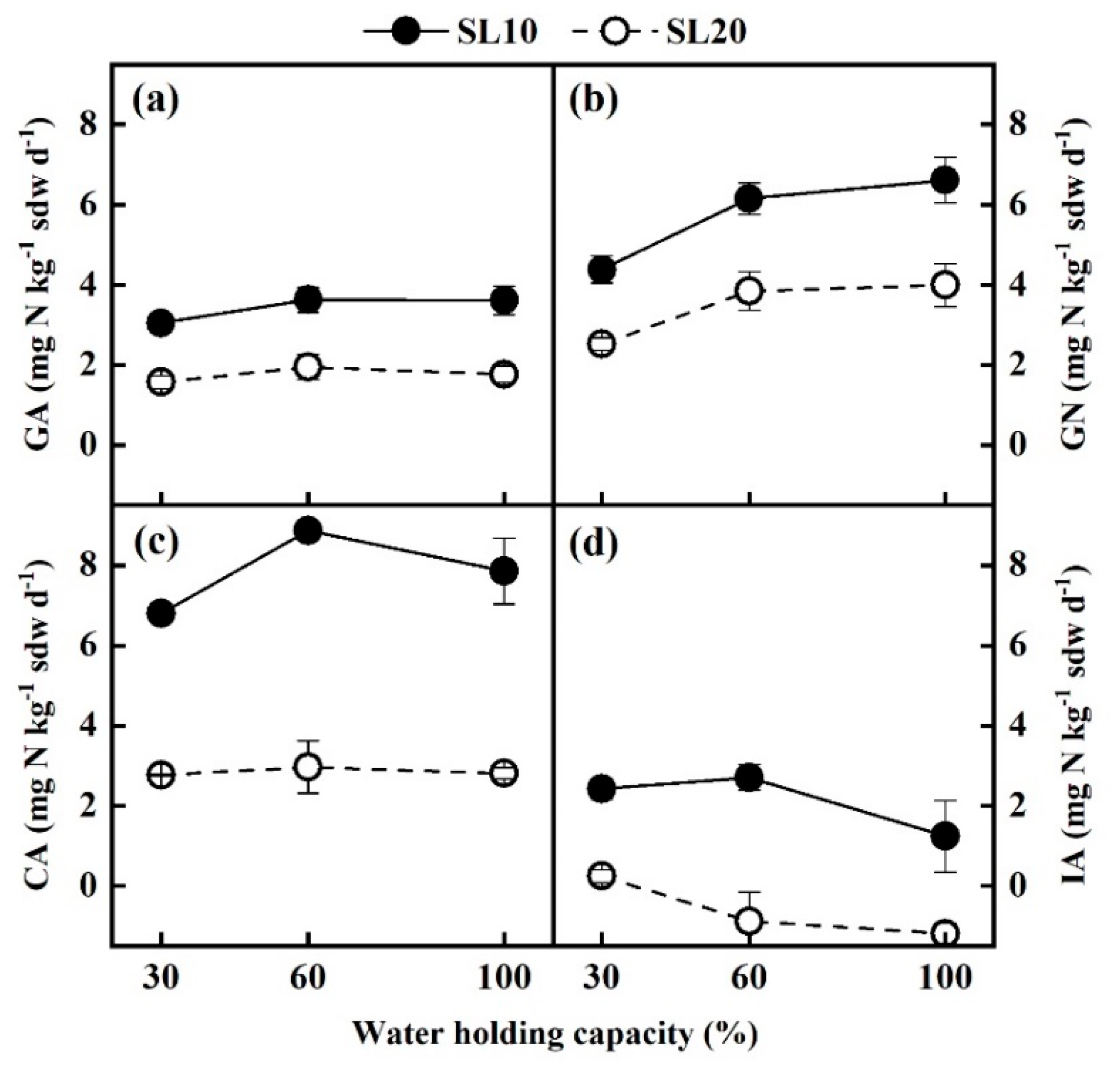
3.4. Gross N Transformation Rates under Different Soil Moisture Conditions
4. Discussion
4.1. Errors of NH4+ Immobilization Rates and a Possibility of Heterotrophic Nitrification
4.2. Effect of Temperature on Soil Gross N Transformation Rates
4.3. Effect of Soil Moisture on Gross N Transformation Rates
4.4. Effect of Soil Depth on Gross N Transformation Rates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Senwo, Z.N.; Tabatabai, M.A. Amino acid composition of soil organic matter. Biol. Fertil. Soils 1998, 26, 235–242. [Google Scholar] [CrossRef]
- Henry, H.A.L.; Jefferies, R.L. Plant amino acid uptake, soluble N turnover and microbial N capture in soils of a grazed Arctic salt marsh. J. Ecol. 2003, 91, 627–636. [Google Scholar] [CrossRef]
- Jones, D.L.; Healey, J.R.; Willett, V.B.; Farrar, J.F.; Hodge, A. Dissolved organic nitrogen uptake by plants—An important N uptake pathway? Soil Biol. Biochem. 2005, 37, 413–423. [Google Scholar] [CrossRef]
- Miller, A.E.; Bowman, W.D. Alpine plants show species-level differences in the uptake of organic and inorganic nitrogen. Plant Soil 2003, 250, 283–292. [Google Scholar] [CrossRef]
- Robertson, G.P.; Groffman, P.M. Nitrogen transformations. In Soil Microbiology, Ecology and Biochemistry, 4th ed.; Paul, E.A., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 421–446. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Hart, S.C.; Nason, G.E.; Myrold, D.D.; Perry, D.A. Dynamics of gross nitrogen transformations in an old-growth forest: The carbon connection. Ecology 1994, 75, 880–891. [Google Scholar] [CrossRef]
- Davidson, E.A.; Hart, S.C.; Firestone, M.K. Internal cycling of nitrate in soils of a mature coniferous forest. Ecology 1992, 73, 1148–1156. [Google Scholar] [CrossRef]
- Cookson, W.R.; Cornforth, I.S.; Rowarth, J.S. Winter soil temperature (2–15 °C) effects on nitrogen transformations in clover green manure amended or unamended soils; a laboratory and field study. Soil Biol. Biochem. 2002, 34, 1401–1415. [Google Scholar] [CrossRef]
- Wu, H.; Dannenmann, M.; Wolf, B.; Han, X.; Zheng, X.; Butterbach-Bahl, K. Seasonality of soil microbial nitrogen turnover in continental steppe soils of Inner Mongolia. Ecosphere 2012, 3, 34. [Google Scholar] [CrossRef]
- Hoyle, F.C.; Murphy, D.V.; Irp, F. Temperature and stubble management influence microbial CO2-C evolution and gross N transformation rates. Soil Biol. Biochem. 2006, 38, 71–80. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Müller, C.; Cai, Z. Temperature sensitivity of gross N transformation rates in an alpine meadow on the Qinghai-Tibetan Plateau. J. Soils Sediments 2017, 17, 423–431. [Google Scholar] [CrossRef]
- Lang, M.; Cai, Z.; Mary, B.; Hao, X.; Chang, S.X. Land-use type and temperature affect gross nitrogen transformation rates in Chinese and Canadian soils. Plant Soil 2010, 334, 377–389. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Wang, S.; Zhang, J.; Cai, Z. Effects of soil moisture on gross N transformations and N2O emission in acid subtropical forest soils. Biol. Fertil. Soils 2014, 50, 1099–1108. [Google Scholar] [CrossRef]
- Jamieson, N.; Monaghan, R.; Barraclough, D. Seasonal trends of gross N mineralization in a natural calcareous grassland. Glob. Chang. Biol. 1999, 5, 423–431. [Google Scholar] [CrossRef]
- Bengtson, P.; Falkengren-Grerup, U.; Bengtsson, G. Relieving substrate limitation-soil moisture and temperature determine gross N transformation rates. Oikos 2005, 111, 81–90. [Google Scholar] [CrossRef]
- Low, A.P.; Stark, J.M.; Dudley, L.M. Effects of soil osmotic potential on nitrification, ammonification, N-assimilation, and nitrous oxide production. Soil Sci. 1997, 162, 16–27. [Google Scholar] [CrossRef]
- Corre, M.D.; Schnabel, R.R.; Stout, W.L. Spatial and seasonal variation of gross nitrogen transformations and microbial biomass in a northeastern us grassland. Soil Biol. Biochem. 2002, 34, 445–457. [Google Scholar] [CrossRef]
- Hu, X.; Liu, C.; Zheng, X.; Dannenmann, M.; Cao, G. Annual dynamics of soil gross nitrogen turnover and nitrous oxide emissions in an alpine shrub meadow. Soil Biol. Biochem. 2019, 138, 107576. [Google Scholar] [CrossRef]
- Guo, Y.; Jiang, M.; Liu, Q.; Xie, Z.; Tang, Z. Climate and vegetation together control the vertical distribution of soil carbon, nitrogen and phosphorus in shrublands in China. Plant Soil 2020, 456, 15–26. [Google Scholar] [CrossRef]
- Guo, Y.; Gheyret, G.; Liu, T.; Zhang, Y.; Kang, M.; Mohhamot, A.; Liu, H.; Ma, W.; Wang, R.; Yu, S.; et al. Distribution patterns and climate limitations of typical shrublands in northern China. Sci. Sin. (Vitae) 2021, 51, 346–361. [Google Scholar] [CrossRef]
- Ross, D.J.; Scott, N.A.; Lambie, S.M.; Trotter, C.M.; Rodda, N.J.; Townsend, J.A. Nitrogen and carbon cycling in a New Zealand pumice soil under a manuka (Leptospermum scoparium) and kanuka (Kunzea ericoides) shrubland. Aust. J. Soil Res. 2009, 47, 725–736. [Google Scholar] [CrossRef]
- Liang, Y.; He, X.; Chen, X.; Hu, Y.; Su, Y. Nitrogen-fixing bacterial communities exhibit strong spatial structure along a slope gradient in Karst shrublands ecosystem. Pol. J. Environ. Stud. 2019, 28, 3369–3376. [Google Scholar] [CrossRef]
- Beier, C.; Emmett, B.A.; Tietema, A.; Schmidt, I.K.; Peñuelas, J.; Láng, E.K.; Duce, P.; De Angelis, P.; Gorissen, A.; Estiarte, M.; et al. Carbon and nitrogen balances for six shrublands across Europe. Glob. Biogeochem. Cycles 2009, 23, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Benvenutto-Vargas, V.P.; Ochoa-Hueso, R. Effects of nitrogen deposition on the spatial pattern of biocrusts and soil microbial activity in a semi-arid Mediterranean shrubland. Funct. Ecol. 2020, 34, 923–937. [Google Scholar] [CrossRef]
- Vourlitis, G.L.; Zorba, G. Nitrogen and carbon mineralization of semi-arid shrubland soil exposed to long-term atmospheric nitrogen deposition. Biol. Fertil. Soils 2007, 43, 611–615. [Google Scholar] [CrossRef]
- Dannenmann, M.; Willibald, G.; Sippel, S.; Butterbach-Bahl, K. Nitrogen dynamics at undisturbed and burned Mediterranean shrublands of Salento Peninsula, Southern Italy. Plant Soil 2011, 343, 5–15. [Google Scholar] [CrossRef]
- Fernández-Fernández, M.; Rütting, T.; González-Prieto, S. Effects of a high-severity wildfire and post-fire straw mulching on gross nitrogen dynamics in Mediterranean shrubland soil. Geoderma 2017, 305, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Dannenmann, M.; Diaz-Pines, E.; Kitzler, B.; Karhu, K.; Tejedor, J.; Ambus, P.; Parra, A.; Sanchez-Martin, L.; Resco, V.; Ramirez, D.A.; et al. Postfire nitrogen balance of Mediterranean shrublands: Direct combustion losses versus gaseous and leaching losses from the postfire soil mineral nitrogen flush. Glob. Chang. Biol. 2018, 24, 4505–4520. [Google Scholar] [CrossRef]
- Song, M.; He, T.; Chen, H.; Wang, K.; Li, D. Dynamics of soil gross nitrogen transformations during post-agricultural succession in a subtropical karst region. Geoderma 2019, 341, 1–9. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022. [Google Scholar]
- Dannenmann, M.; Gasche, R.; Ledebuhr, A.; Papen, H. Effects of forest management on soil N cycling in beech forests stocking on calcareous soils. Plant Soil 2006, 287, 279–300. [Google Scholar] [CrossRef]
- Kirkham, D.; Bartholomew, W.V. Equations for following nutrient transformations in soil, utilizing tracer data1. Soil Sci. Soc. Am. J. 1954, 18, 33–34. [Google Scholar] [CrossRef]
- Davidson, E.A.; Hart, S.C.; Shanks, C.A.; Firestone, M.K. Measuring gross nitrogen mineralization, immobilization, and nitrification by 15N isotopic pool dilution in intact soil cores. J. Soil Sci. 1991, 42, 335–349. [Google Scholar] [CrossRef]
- Verchot, L.V.; Holmes, Z.; Mulon, L.; Groffman, P.M.; Lovett, G.M. Gross vs net rates of N mineralization and nitrification as indicators of functional differences between forest types. Soil Biol. Biochem. 2001, 33, 1889–1901. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Z.; Unteregelsbacher, S.; Lu, H.; Gschwendtner, S.; Gasche, R.; Kolar, A.; Schloter, M.; Kiese, R.; Butterbach-Bahl, K.; et al. Climate change amplifies gross nitrogen turnover in montane grasslands of Central Europe both in summer and winter seasons. Glob. Chang. Biol. 2016, 22, 2963–2978. [Google Scholar] [CrossRef] [Green Version]
- Huygens, D.; Boeckx, P.; Templer, P.H.; Paulino, L.; Van Cleemput, O.; Oyarzun, C.; Muller, C.; Godoy, R. Mechanisms for retention of bioavailable nitrogen in volcanic rainforest soils. Nat. Geosci. 2008, 1, 543–548. [Google Scholar] [CrossRef]
- Liu, R.; Suter, H.; He, J.; Hayden, H.; Chen, D. Influence of temperature and moisture on the relative contributions of heterotrophic and autotrophic nitrification to gross nitrification in an acid cropping soil. J. Soil Sediments 2015, 15, 2304–2309. [Google Scholar] [CrossRef]
- Zhang, J.; Müller, C.; Cai, Z. Heterotrophic nitrification of organic N and its contribution to nitrous oxide emissions in soils. Soil Biol. Biochem. 2015, 84, 199–209. [Google Scholar] [CrossRef]
- Sun, L.; Xia, Z.; Sang, C.; Wang, X.; Peng, B.; Wang, C.; Zhang, J.; Müller, C.; Bai, E. Soil resource status affects the responses of nitrogen processes to changes in temperature and moisture. Biol. Fertil. Soils 2019, 55, 629–641. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Brookes, P.C.; Jenkinson, D.S. Survival of the soil microbial biomass at elevated temperatures. Soil Biol. Biochem. 1990, 22, 1129–1136. [Google Scholar] [CrossRef]
- Mukhtar, H.; Lin, Y.-P.; Lin, C.-M.; Lin, Y.-R. Relative Abundance of Ammonia Oxidizing Archaea and Bacteria Influences Soil Nitrification Responses to Temperature. Microorganisms 2019, 7, 526. [Google Scholar] [CrossRef] [Green Version]
- Bei, S.; Tian, Y.; Zhao, J.; Zhang, H.; Christie, P.; Li, X.; Jia, Z.; Zhang, J. Temperature-dependent changes in active nitrifying communities in response to field fertilization legacy. Biol. Fertil. Soils 2021, 57, 1–14. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Zhang, J.; Wang, S.; Cai, Z. The different temperature sensitivity of gross N transformations between the coniferous and broad-leaved forests in subtropical China. Soil Sci. Plant Nutr. 2015, 61, 506–515. [Google Scholar] [CrossRef] [Green Version]
- Dan, X.; Chen, Z.; Dai, S.; He, X.; Cai, Z.; Zhang, J.; Müller, C. Effects of changing temperature on gross N transformation rates in acidic subtropical forest soils. Forests 2019, 10, 894. [Google Scholar] [CrossRef] [Green Version]
- Zaman, M.; Chang, S.X. Substrate type, temperature, and moisture content affect gross and net N mineralization and nitrification rates in agroforestry systems. Biol Fertil Soils 2004, 39, 269–279. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. (Eds.) IPCC, Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, J. Evaluation of reanalysis datasets against observational soil temperature data over China. Clim. Dyn. 2018, 50, 317–337. [Google Scholar] [CrossRef]
- Zhan, M.; Xia, L.; Zhan, L.; Wang, Y. Recognition of changes in air and soil temperatures at a station typical of China’s subtropical monsoon region (1961–2018). Adv. Meteorol. 2019, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philos. Trans. R. Soc. B 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Zhao, W.; Hu, H.; Feng, X.; Müller, C.; Cai, Z. Soil gross nitrogen transformations along the Northeast China Transect (NECT) and their response to simulated rainfall events. Sci. Rep. 2016, 6, 22830. [Google Scholar] [CrossRef] [Green Version]
- Butterbach-Bahl, K.; Gundersen, P. Nitrogen Processes in Terrestrial Ecosystems; University of Reading: Reading, UK, 2011; Volume 43, pp. 99–125. [Google Scholar] [CrossRef] [Green Version]
- Stark, J.M.; Firestone, M.K. Mechanisms for soil moisture effects on activity of nitrifying bacteria. Appl. Environ. Microbiol. 1995, 61, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Cai, Z.; Zhang, J.; Zhang, J.; Lang, M.; Mary, B.; Chang, S.X. Soil moisture effects on gross nitrification differ between adjacent grassland and forested soils in central Alberta, Canada. Plant Soil 2012, 352, 289–301. [Google Scholar] [CrossRef]
- Osborne, B.B.; Baron, J.S.; Wallenstein, M.D. Moisture and temperature controls on nitrification differ among ammonia oxidizer communities from three alpine soil habitats. Front. Earth Sci. 2016, 10, 1–12. [Google Scholar] [CrossRef]
- Zaman, M.; Di, H.J.; Cameron, K.C.; Frampton, C.M. Gross nitrogen mineralization and nitrification rates and their relationships to enzyme activities and the soil microbial biomass in soils treated with dairy shed effluent and ammonium fertilizer at different water potentials. Biol. Fertil. Soils 1999, 29, 178–186. [Google Scholar] [CrossRef]
- Stange, C.F. A novel approach to combine response functions in ecological process modelling. Ecol. Model. 2007, 204, 547–552. [Google Scholar] [CrossRef]
- Linn, D.M.; Doran, J.W. Effect of water-filled pore space on carbon dioxide and nitrous oxide production in tilled and nontilled soils. Soil Sci. Soc. Am. J. 1984, 48, 647–653. [Google Scholar] [CrossRef] [Green Version]
- Kiese, R.; Hewett, B.; Butterbach-Bahl, K. Seasonal dynamic of gross nitrification and N2O emission at two tropical rainforest sites in Queensland, Australia. Plant Soil 2008, 309, 105–117. [Google Scholar] [CrossRef]
- O’Gorman, P.A. Precipitation extremes under climate change. Curr. Clim. Chang. Rep. 2015, 1, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Shi, P.; Zhang, Q.; Wang, J.; Wu, J.; Chen, D. Evolution of future precipitation extremes: Viewpoint of climate change classification. Int. J. Climatol. 2022, 42, 1220–1230. [Google Scholar] [CrossRef]
- Sharma, N.; Kumar, S. Nitrogen transformation rates in the Himalayan soils at different temperature and elevation conditions. J. Soil Sediments 2021, 21, 13–26. [Google Scholar] [CrossRef]
- Barrett, J.E.; Burke, I.C. Potential nitrogen immobilization in grassland soils across a soil organic matter gradient. Soil Biol. Biochem. 2000, 32, 1707–1716. [Google Scholar] [CrossRef]
- Accoe, F.; Boeckx, P.; Busschaert, J.; Hofman, G.; Van Cleemput, O. Gross N transformation rates and net N mineralisation rates related to the C and N contents of soil organic matter fractions in grassland soils of different age. Soil Biol. Biochem. 2004, 36, 2075–2087. [Google Scholar] [CrossRef]
- Booth, M.S.; Stark, J.M.; Rastetter, E. Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data. Ecol. Monogr. 2005, 75, 139–157. [Google Scholar] [CrossRef] [Green Version]
- Lang, M.; Li, P.; Ti, C.; Zhu, S.; Yan, X.; Chang, S.X. Soil gross nitrogen transformations are related to land-uses in two agroforestry systems. Ecol. Eng. 2019, 127, 431–439. [Google Scholar] [CrossRef]
- Elrys, A.S.; Chen, Z.; Wang, J.; Uwiragiye, Y.; Helmy, A.M.; Desoky, E.M.; Cheng, Y.; Zhang, J.; Cai, Z.; Muller, C. Global patterns of soil gross immobilization of ammonium and nitrate in terrestrial ecosystems. Glob. Chang. Biol. 2022, 28, 4472–4488. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Stark, J.M. Plant species effects and carbon and nitrogen cycling in a sagebrush-crested wheatgrass soil. Soil Biol. Biochem. 2000, 32, 47–57. [Google Scholar] [CrossRef]
- Mack, M.C.; D’Antonio, C.M. Exotic grasses alter controls over soil nitrogen dynamics in a Hawaiian woodland. Ecol. Appl. 2003, 13, 154–166. [Google Scholar] [CrossRef]
- Stiles, K. Quantification of Gross Nitrogen Transformation Rates within a Conventional Potato Rotation Using Stable Isotopes. Master’s Thesis, Dalhousie University, Halifax, NS, USA, 6 December 2012. [Google Scholar]
- Cheng, Y.; Wang, J.; Wang, J.; Chang, S.X.; Wang, S. The quality and quantity of exogenous organic carbon input control microbial NO3− immobilization: A meta-analysis. Soil Biol. Biochem. 2017, 115, 357–363. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soil Depth | SL10 | SL20 |
|---|---|---|
| WHC (%) | 53.1 ± 1.5 a | 49.6 ± 1.5 a |
| pH | 7.4 ± 0.4 a | 7.5 ± 0.2 a |
| Clay (%) | 6.8 ± 1.1 a | 7.6 ± 1.0 a |
| Silt (%) | 28.1 ± 4.0 a | 30.3 ± 4.4 a |
| Sand (%) | 65.1 ± 3.7 a | 62.2 ± 4.5 a |
| SOC (g kg−1) | 19.1 ± 1.5 a | 14.0 ± 1.3 b |
| TN (g kg−1) | 1.81 ± 0.09 a | 1.38 ± 0.08 b |
| WSOC (mg C kg−1 sdw) | 11.7 ± 0.9 a | 9.1 ± 0.3 b |
| NH4+ (mg N kg−1 sdw) | 11.1 ± 0.8 a | 6.0 ± 0.7 b |
| NO3− (mg N kg−1 sdw) | 10.9 ± 0.7 a | 6.7 ± 0.3 b |
| Incubation Temperature/Moisture | SL10 | SL20 | |||
|---|---|---|---|---|---|
| t1 | t2 | t1 | t2 | ||
| 15N enrichment of NH4+ in the 15NH4+ labeled soils (atom%) | 5 °C | 2.35 ± 0.02 | 2.18 ± 0.08 | 2.90 ± 0.07 | 2.79 ± 0.02 |
| 15 °C | 2.13 ± 0.03 | 1.88 ± 0.004 | 2.66 ± 0.04 | 2.39 ± 0.004 | |
| 25 °C | 1.81 ± 0.02 | 1.31 ± 0.04 | 2.21 ± 0.02 | 1.77 ± 0.03 | |
| 35 °C | 1.35 ± 0.02 | 0.92 ± 0.004 | 1.70 ± 0.01 | 1.21 ± 0.01 | |
| 30% WHC | 1.65 ± 0.04 | 1.30 ± 01 | 2.09 ± 0.01 | 1.76 ± 0.02 | |
| 60% WHC | 1.59 ± 0.02 | 1.17 ± 0.02 | 2.10 ± 0.03 | 1.68 ± 0.03 | |
| 100% WHC | 1.60 ± 0.08 | 1.02 ± 0.04 | 2.05 ± 0.05 | 1.60 ± 0.02 | |
| 15N enrichment of NO3− in the 15NO3− labeled soils (atom%) | 5 °C | 2.54 ± 0.01 | 2.34 ± 0.11 | 3.91 ± 0.06 | 3.77 ± 0.11 |
| 15 °C | 2.41 ± 0.06 | 2.02 ± 0.06 | 3.46 ± 0.05 | 3.18 ± 0.12 | |
| 25 °C | 2.05 ± 0.01 | 1.76 ± 0.02 | 3.27 ± 0.03 | 2.72 ± 0.06 | |
| 35 °C | 1.89 ± 0.04 | 1.52 ± 0.02 | 3.02 ± 0.01 | 2.42 ± 0.03 | |
| 30% WHC | 2.04 ± 0.02 | 1.73 ± 0.03 | 2.09 ± 0.01 | 1.76 ± 0.02 | |
| 60% WHC | 1.96 ± 0.02 | 1.61 ± 0.005 | 2.10 ± 0.03 | 1.68 ± 0.03 | |
| 100% WHC | 1.95 ± 0.02 | 1.59 ± 0.04 | 2.05 ± 0.05 | 1.60 ± 0.02 | |
| NH4+ concentrations in the 15NH4+ labeled soils (mg N kg−1 sdw) | 5 °C | 13.0 ± 0.2 | 13.6 ± 0.2 | 8.7 ± 0.04 | 8.3 ± 0.1 |
| 15 °C | 12.3 ± 0.2 | 10.3 ± 0.1 | 8.1 ± 0.1 | 7.8 ± 0.1 | |
| 25 °C | 10.0 ± 0.03 | 4.9 ± 0.1 | 7.7 ± 0.1 | 6.2 ± 0.2 | |
| 35 °C | 11.1 ± 0.4 | 6.3 ± 0.2 | 8.6 ± 0.2 | 7.9 ± 0.2 | |
| 30% WHC | 11.6 ± 0.2 | 7.8 ± 0.3 | 8.1 ± 0.1 | 6.9 ± 0.2 | |
| 60% WHC | 11.4 ± 0.3 | 6.2 ± 0.02 | 7.4 ± 0.1 | 6.4 ± 0.7 | |
| 100% WHC | 8.1 ± 0.5 | 3.8 ± 0.1 | 6.3 ± 0.3 | 5.2 ± 0.1 | |
| NO3− concentrations in the 15NO3− labeled soils (mg N kg−1 sdw) | 5 °C | 12.3 ± 0.1 | 12.2 ± 0.4 | 7.2 ± 0.02 | 7.8 ± 0.1 |
| 15 °C | 11.0 ± 0.4 | 9.3 ± 0.1 | 7.3 ± 0.1 | 7.0 ± 0.2 | |
| 25 °C | 9.0 ± 0.1 | 4.0 ± 0.04 | 7.0 ± 0.1 | 5.4 ± 0.2 | |
| 35 °C | 10.3 ± 0.3 | 5.4 ± 0.2 | 7.7 ± 0.2 | 7.0 ± 0.3 | |
| 30% WHC | 10.8 ± 0.1 | 7.1 ± 0.2 | 7.5 ± 0.2 | 6.4 ± 0.3 | |
| 60% WHC | 9.7 ± 0.05 | 5.3 ± 0.3 | 6.7 ± 0.04 | 5.5 ± 0.2 | |
| 100% WHC | 7.4 ± 0.3 | 3.5 ± 0.5 | 5.8 ± 0.2 | 5.0 ± 0.2 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Zhang, Y.; Wang, D.; Ma, J.; Xue, K.; An, Z.; Luo, W.; Sheng, Y. Effects of Temperature and Humidity on Soil Gross Nitrogen Transformation in a Typical Shrub Ecosystem in Yanshan Mountain and Hilly Region. Life 2023, 13, 643. https://doi.org/10.3390/life13030643
Hu X, Zhang Y, Wang D, Ma J, Xue K, An Z, Luo W, Sheng Y. Effects of Temperature and Humidity on Soil Gross Nitrogen Transformation in a Typical Shrub Ecosystem in Yanshan Mountain and Hilly Region. Life. 2023; 13(3):643. https://doi.org/10.3390/life13030643
Chicago/Turabian StyleHu, Xiaoxia, Yuanxun Zhang, Dong Wang, Jian Ma, Kaibing Xue, Zhaobo An, Wenxing Luo, and Yizhi Sheng. 2023. "Effects of Temperature and Humidity on Soil Gross Nitrogen Transformation in a Typical Shrub Ecosystem in Yanshan Mountain and Hilly Region" Life 13, no. 3: 643. https://doi.org/10.3390/life13030643