

Brachypodium distachyon Seedlings Display Accession-Specific Morphological and Transcriptomic Responses to the Microgravity Environment of the International Space Station

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. A Novel APEX Growth Unit Allows Transportation of Pre-Planted Dry Brachypodium distachyon Seeds to ISS and Subsequent Activation of Germination
2.2. Plant Materials and Growth Conditions
2.3. ISS Experiment
2.4. Ground Control
2.5. RNA Extraction and RNAseq Analysis
2.6. RNA Sequence Mapping and Transcriptional Profiling
2.7. Bioinformatic Analysis, Annotation, and GO Enrichment of Differentially Expressed Genes
2.8. Morphology Measurement
2.9. qRT-PCR Analysis
3. Results
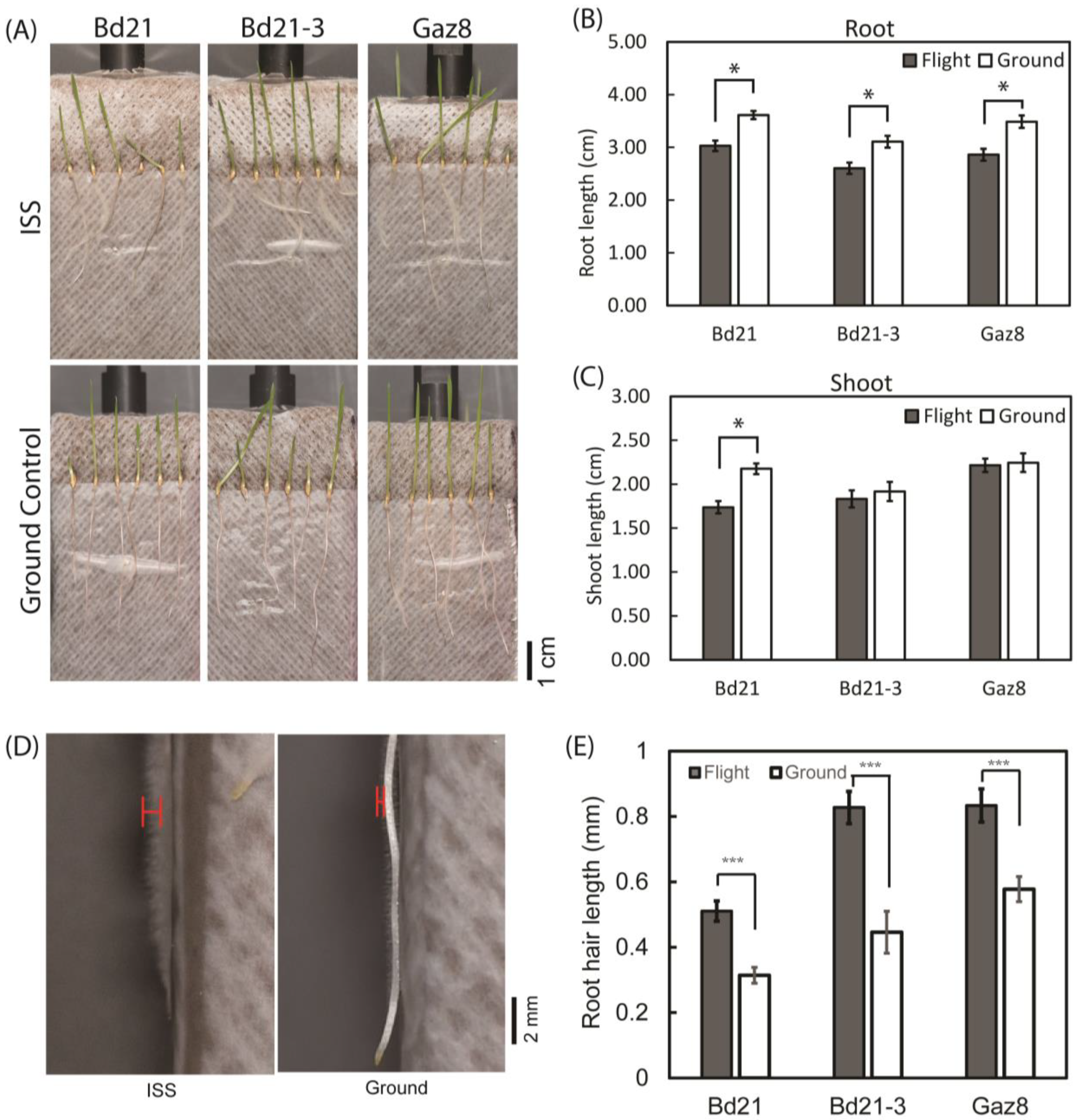
3.1. Microgravity Affects Seedling Growth in an Accession-Specific Manner
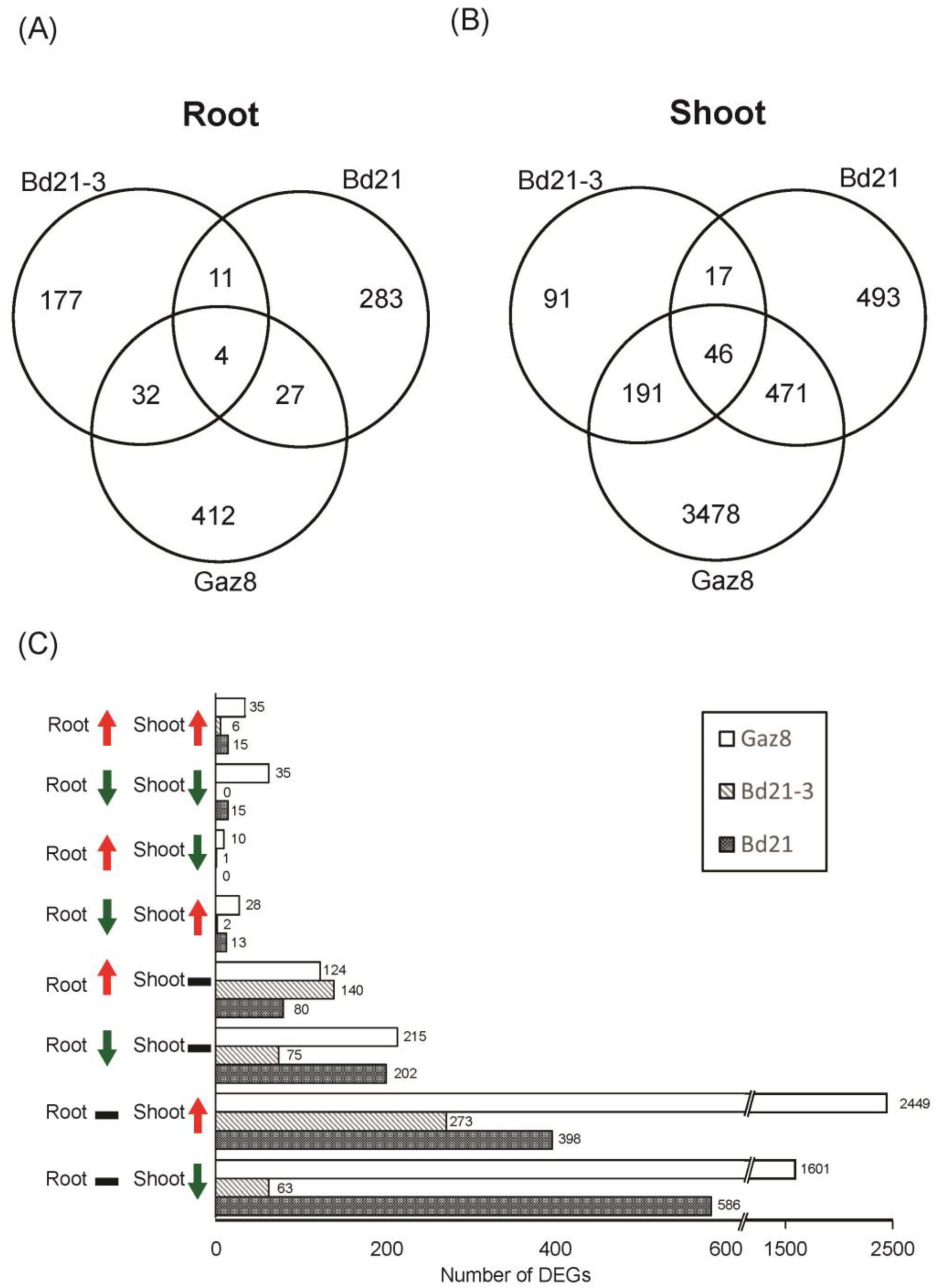
3.2. Microgravity-Exposed Seedling Roots and Shoots Display Distinct Transcriptomic Profiles Compared to Ground Controls
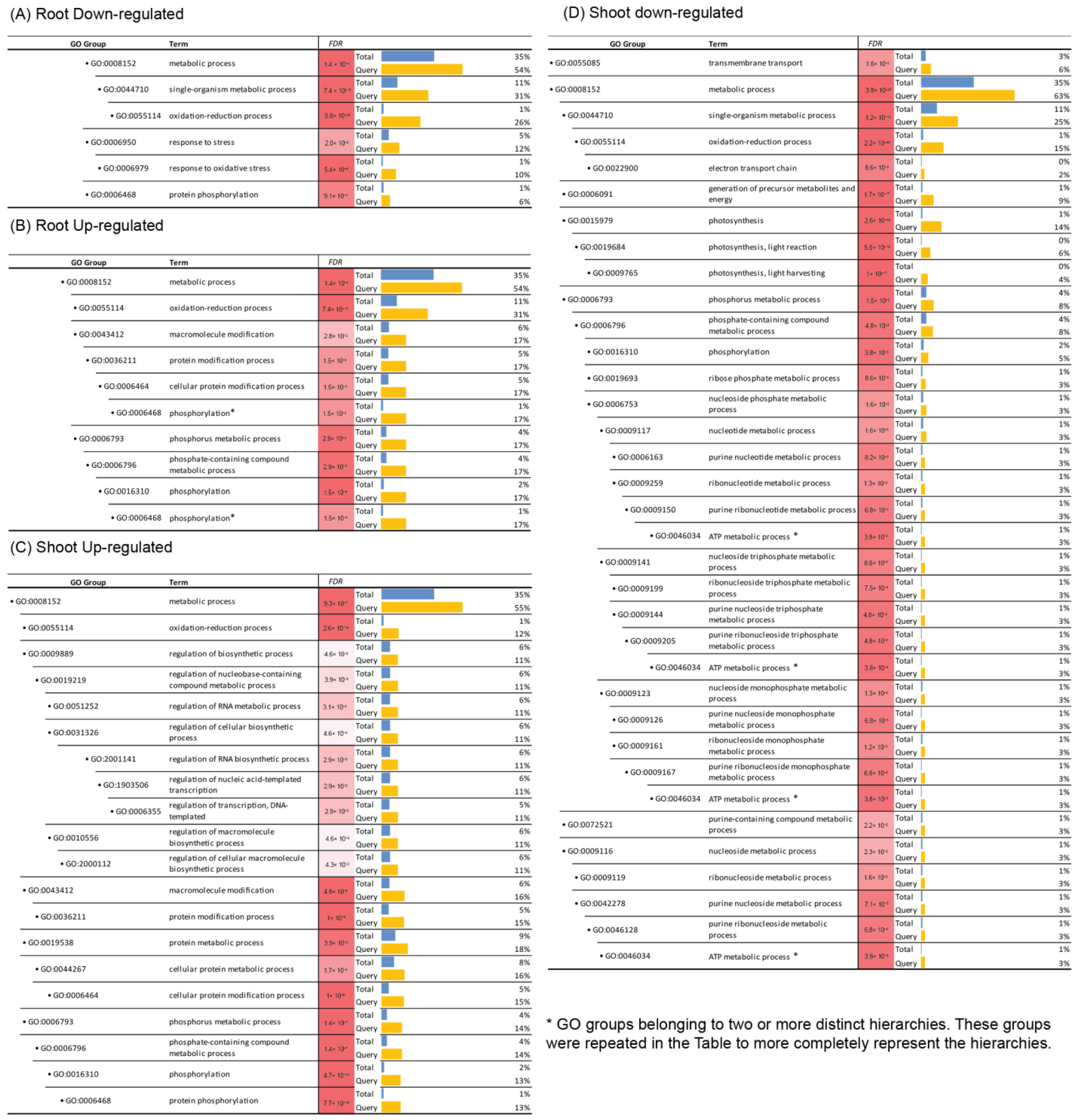
3.3. GO Enrichment Analysis Suggests That Brachypodium Roots and Shoots Deploy Distinct Biological Strategies to Cope with the Microgravity Environment on ISS
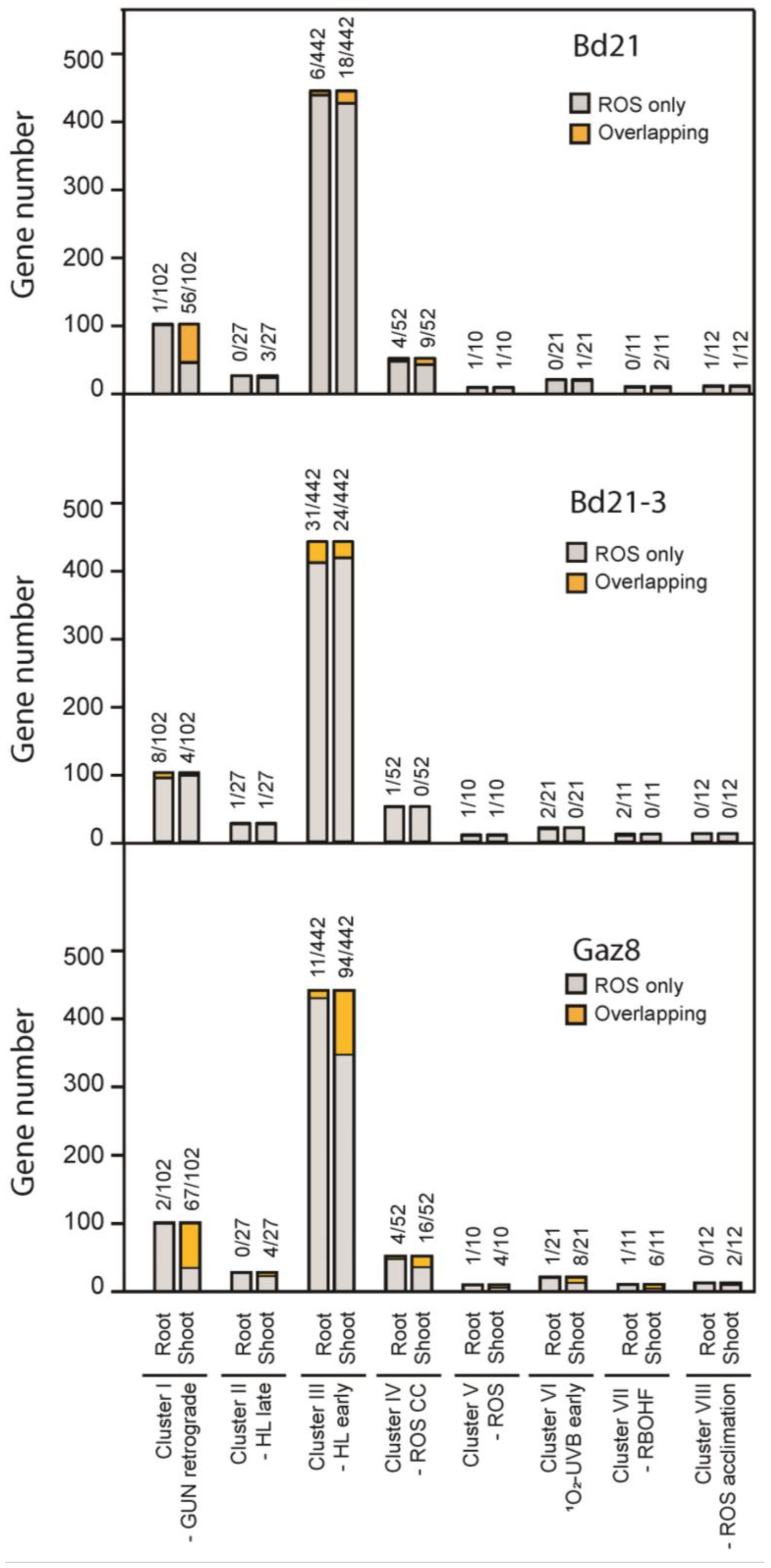
3.4. Brachypodium Genes Associated with Photosynthesis Are Down-Regulated under Microgravity Conditions on ISS
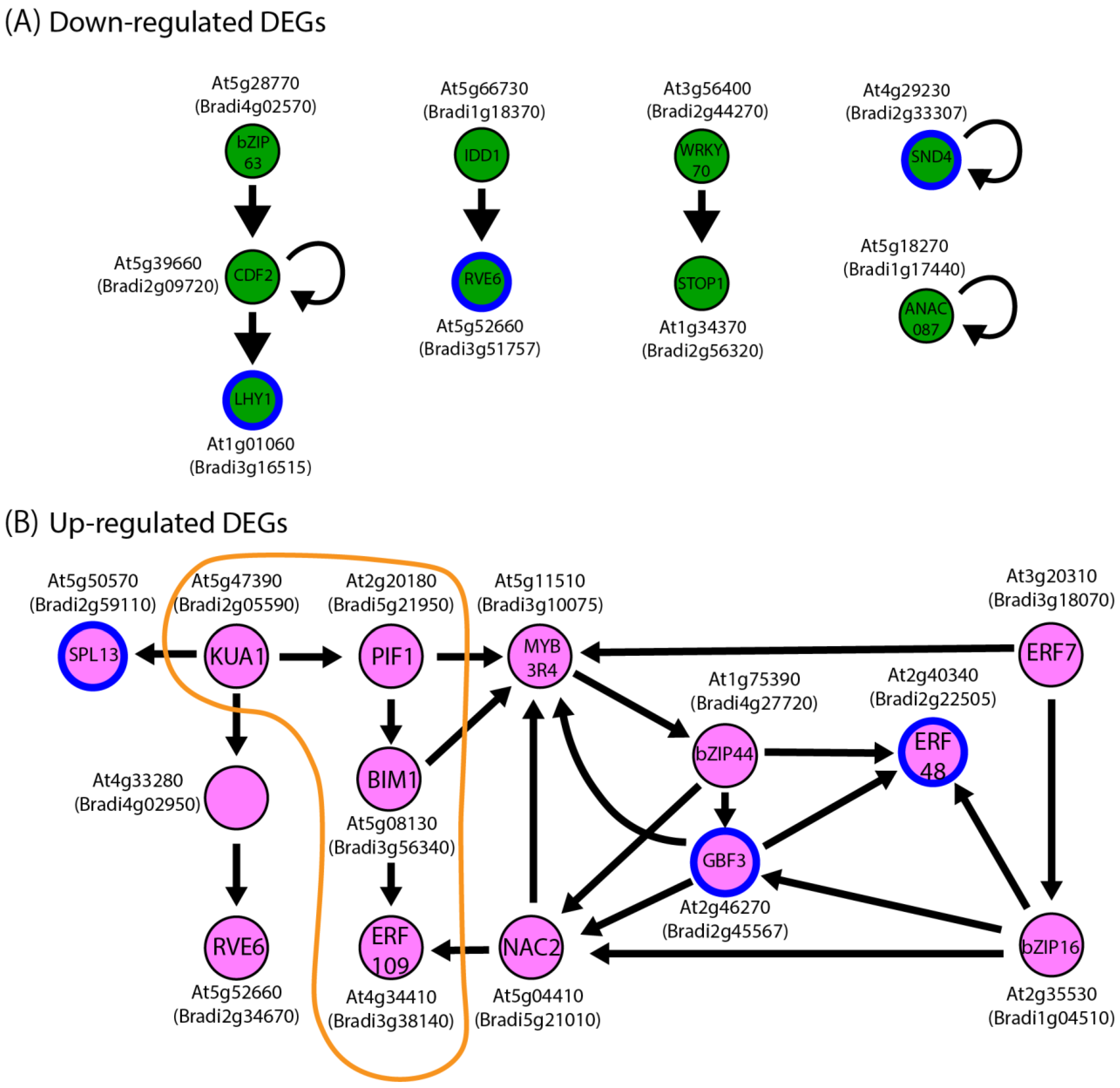
3.5. Genes Annotated as Potential Contributors to Transcriptional and Post-Translational Regulation Are Up-Regulated in Microgravity-Grown Bd21 and Gaz8 Shoots
3.6. Genes Associated with the “Translation” GO Group (GO:0006412) Are Down-Regulated in Gaz8 Shoots and Roots under the Microgravity Environment on ISS
3.7. Root Hair Growth Is Enhanced under Microgravity Conditions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Porterfield, D. The biophysical limitations in physiological transport and exchange in plants grown in microgravity. J. Plant Growth Reg. 2002, 21, 177–190. [Google Scholar] [CrossRef]
- Kiss, J.; Millar, K.; Edelmann, R. Phototropism of Arabidopsis thaliana in microgravity and fractional gravity on the International Space Station. Planta 2012, 236, 635–645. [Google Scholar] [CrossRef]
- Porterfield, D.; Musgrave, M. The tropic response of plant roots to oxygen: Oxytropism in Pisum sativum L. Planta 1998, 20, 1–6. [Google Scholar] [CrossRef]
- Yoshioka, R.; Soga, K.; Wakabayashi, K.; Takeba, G.; Hoson, T. Hypergravity-induced changes in gene expression in Arabidopsis hypocotyls. Adv. Space Res. 2003, 31, 2187–2193. [Google Scholar] [CrossRef]
- Liao, J.; Liu, G.; Monje, O.; Stutte, G.W.; Porterfield, D.M. Induction of hypoxic root metabolism results from physical limitations in O2 bioavailability in microgravity. Adv. Space Res. 2004, 34, 1579–1584. [Google Scholar] [CrossRef]
- Choi, W.; Barker, R.; Kim, S.; Swanson, S.; Gilroy, S. Variation in the transcriptome of different ecotypes of Arabidopsis thaliana reveals signatures of oxidative stress in plant responses to spaceflight. Am. J. Bot. 2019, 106, 123–136. [Google Scholar] [CrossRef]
- Kiss, J.; Brinkmann, E.; Brillouet, C. Development and growth of several strains of Arabidopsis seedlings in microgravity. Int. J. Plant Sci. 2000, 161, 55–62. [Google Scholar] [CrossRef]
- Millar, K.; Johnson, C.; Edelmann, R.; Kiss, J. An endogenous growth pattern of roots is revealed in seedlings grown in microgravity. Astrobiology 2011, 11, 786–797. [Google Scholar] [CrossRef]
- Chancellor, J.; Scott, G.; Sutton, J. Space radiation: The number one risk to astronaut health beyond low earth orbit. Life 2014, 4, 491–510. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.; Amalfitano, C.; Ferl, R. Plant growth strategies are remodeled by spaceflight. BMC Plant Biol. 2012, 12, 232. [Google Scholar] [CrossRef] [Green Version]
- Hilaire, E.; Paulsen, A.; Brown, C.; Guikema, J. Plastid distribution in columella cells of a starchless Arabidopsis mutant grown in microgravity. Plant Cell Physiol. 1997, 38, 490–494. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.; Zupanska, A.; Schultz, E.; Ferl, R. Organ-specific remodeling of the Arabidopsis transcriptome in response to spaceflight. BMC Plant Biol. 2013, 13, 112. [Google Scholar] [CrossRef] [Green Version]
- Stutte, G.; Monje, O.; Hatfield, R.; Paul, A.; Ferl, R.; Simone, C. Microgravity effects on leaf morphology, cell structure, carbon metabolism and mRNA expression of dwarf wheat. Planta 2006, 224, 1038–1049. [Google Scholar] [CrossRef]
- Kwon, T.; Sparks, J.; Nakashima, J.; Allen, S.; Tang, Y.; Blancaflor, E. Transcriptional response of Arabidopsis seedlings during spaceflight reveals peroxidase and cell wall remodeling genes associated with root hair development. Am. J. Bot. 2015, 102, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.-L.; Zupanska, A.; Ostrow, D.; Zhang, Y.; Sun, Y.; Li, J.-L.; Shanker, S.; Farmerie, W.; Amalfitano, C.; Ferl, R. Spaceflight transcriptomes: Unique responses to a novel environment. Astrobiology 2012, 12, 40–56. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.; Sng, N.; Zupanska, A.; Krishnamurthy, A.; Schultz, E.; Ferl, R. Genetic dissection of the Arabidopsis spaceflight transcriptome: Are some responses dispensable for the physiological adaptation of plants to spaceflight? PLoS ONE 2017, 12, e0180186. [Google Scholar] [CrossRef] [Green Version]
- Correll, M.; Pyle, T.; Millar, K.; Sun, Y.; Yao, J.; Edelmann, R.; Kiss, J. Transcriptome analyses of Arabidopsis thaliana seedlings grown in space: Implications for gravity-responsive genes. Planta 2013, 238, 519–533. [Google Scholar] [CrossRef]
- Zupanska, A.; Denison, F.; Ferl, R.; Paul, A.-L. Spaceflight engages heat shock protein and other molecular chaperone genes in tissue culture cells of Arabidopsis thaliana. Am. J. Bot. 2013, 100, 235–248. [Google Scholar] [CrossRef]
- Basu, P.; Kruse, C.; Luesse, D.; Wyatt, S. Growth in spaceflight hardware results in alterations to the transcriptome and proteome. Life Sci. Space Res. 2017, 15, 88–96. [Google Scholar] [CrossRef]
- Li, H.; Lu, J.; Zhao, H.; Sun, Q.; Yu, F.; Pan, Y.; Chen, Y.; Su, L.; Liu, M. The impact of space environment on gene expression in Arabidopsis thaliana seedlings. Sci. China 2017, 60, 902–910. [Google Scholar] [CrossRef]
- Fengler, S.; Spirer, I.; Neef, M.; Ecke, M.; Nieselt, K.; Hampp, R. A whole-genome microarray study of Arabidopsis thaliana semisolid callus cultures exposed to microgravity and in microgravity related spaceflight conditions for 5 days on board of Shenzhou 8. Biomed. Res. Int. 2015, 2015, 547495. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.; Subramanian, A.; Pattathil, S.; Correll, M.; Kiss, J. Comparative transcriptomics indicate changes in cell wall organization and stress response in seedlings during spaceflight. Am. J. Bot. 2017, 104, 1219–1231. [Google Scholar] [CrossRef] [Green Version]
- Ferl, R.; Koh, J.; Denison, F.; Paul, A.-L. Spaceflight induces specific alterations in the proteomes of Arabidopsis. Astrobiology 2015, 15, 32–56. [Google Scholar] [CrossRef]
- Zhou, M.; Sng, N.; LeFrois, C.; Paul, A.; Ferl, R. Epigenomics in an extraterrestrial environment: Organ-specific alteration of DNA methylation and gene expression elicited by spaceflight in Arabidopsis thaliana. BMC Genom. 2019, 20, 205. [Google Scholar] [CrossRef] [Green Version]
- Vandenbrink, J.; Kiss, J. Space, the final frontier: A critical review of recent experiments performed in microgravity. Plant Sci. 2016, 243, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, J.; Liao, F.; Sparks, J.A.; Tang, Y.; Blancaflor, E.B. The actin cytoskeleton is a suppressor of the endogenous skewing behaviour of Arabidopsis primary roots in microgravity. Plant Biol. 2014, 16, 142–150. [Google Scholar] [CrossRef]
- Chaw, S.-M.; Chang, C.-C.; Chen, H.-L.; Li, W.-H. Dating the Monocot-Dicot divergence and the origin of core eudicots using whole chloroplast genomes. J. Mol. Evol. 2004, 58, 424–441. [Google Scholar] [CrossRef]
- Tairbekov, M.; Parfyonov, G.; Platonova, R.; Aramova, V.; Golov, V.; Rostopshina, A.; Lyubchenko, V.; Chuchkin, V. Biological investigations aboard the biosatellite Cosmos-1129. Adv. Space Res. 1981, 1, 89–94. [Google Scholar] [CrossRef]
- Hoson, T.; Soga, K.; Wakabayashi, K.; Kamisaka, S.; Tanimoto, E. Growth and cell wall changes in rice roots during spaceflight. Plant Soil 2003, 255, 19–26. [Google Scholar] [CrossRef]
- Shagimardanova, E.; Gusev, O.; Sychev, V.; Levinskikh, M.; Sharipova, M.; Il’inskaya, O.; Bingham, G.; Sugimoto, M. Expression of stress response genes in barley Hordeum vulgare in a spaceflight environment. Mol. Biol. 2010, 44, 734–740. [Google Scholar] [CrossRef]
- Chen, B.; Wang, Y. Proteomic and physiological studies provide insight into photosynthetic response of rice (Oryza sativa L.) seedlings to microgravity. Photochem. Photobiol. 2016, 92, 561–570. [Google Scholar] [CrossRef]
- Brkljacic, J.; Grotewold, E.; Scholl, R.; Mockler, T.; Garvin, D.; Vain, P.; Brutnell, T.; Sibout, R.; Bevan, M.; Budak, H.; et al. Brachypodium as a model for the grasses: Today and the future. Plant Physiol. 2011, 157, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, C.; Barker, R.; Choi, W.-G.; Swanson, S.; Stephens, S.; Huber, C.; Nimunkar, A.; Gilroy, S. Development of equipment that uses far-red light to impose seed dormancy in Arabidopsis for spaceflight. Gravit. Space Res. 2016, 4, 8–19. [Google Scholar] [CrossRef]
- Barrero, J.M.; Jacobsen, J.V.; Talbot, M.J.; White, R.G.; Swain, S.M.; Garvin, D.F.; Gubler, F. Grain dormancy and light quality effects on germination in the model grass Brachypodium distachyon. New Phytol. 2012, 193, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.G.; Krikorian, A.D. Shoot growth in aseptically cultivated daylily and haplopappus plantlets after a 5-day spaceflight. Physiol. Plant 1992, 86, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, B.C.; Brown, C.S.; Levine, H.G.; Krikorian, A.D. Growth and photosynthetic responses of wheat plants grown in space. Plant Physiol. 1996, 110, 801–806. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, S.; Campana, S. ISS Payloads Office Cold Stowage Overview. Available online: https://www.nasa.gov/pdf/360392main_P03_0915_JSC.pdf (accessed on 4 June 2021).
- Massa, G.; Dufour, N.; Carver, J.; Hummerick, M.; Wheeler, R.; Morrow, R.; Smith, T. VEG-01: Veggie hardware validation testing on the International Space Station. Open Agric. 2017, 2, 33–41. [Google Scholar] [CrossRef]
- Trapnell, C.; Hendrickson, D.; Sauvageau, M.; Goff, L.; Rinn, J.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotech 2013, 31, 46–53. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—Te DESeq Package. Bioconductor. 2013. Available online: http://www.bioconductor.org/packages/devel/bioc/vignettes/DESeq/inst/doc/DESeq.pdf (accessed on 4 June 2021).
- Robinson, M.; McCarthy, D.; Smyth, G. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Vogel, J.P.; Garvin, D.F.; Mockler, T.C.; Schmutz, J.; Rokhsar, D.; Bevan, M.W.; Barry, K.; Lucas, S.; Harmon-Smith, M.; Lail, K.; et al. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef] [Green Version]
- Willems, P.; Mhamdi, A.; Stael, S.; Storme, V.; Kerchev, P.; Noctor, G.; Gevaert, K.; Van Breusegem, F. The ROS Wheel: Refining ROS Transcriptional Footprints. Plant Physiol. 2016, 171, 1720–1733. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. agriGO: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant Cis-Acting Regulatory DNA Elements (PLACE) Database; Oxford Academic: Oxford, UK, 1999; Volume 27, pp. 297–300. [Google Scholar]
- Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2019, 48, D1104–D1113. [Google Scholar] [CrossRef]
- Hong, S.Y.; Seo, P.J.; Yang, M.S.; Xiang, F.; Park, C.M. Exploring valid reference genes for gene expression studies in Brachypodium distachyon by real-time PCR. BMC Plant Biol. 2008, 8, 112. [Google Scholar] [CrossRef] [Green Version]
- Vandenbrink, J.P.; Herranz, R.; Poehlman, W.L.; Alex Feltus, F.; Villacampa, A.; Ciska, M.; Javier Medina, F.; Kiss, J.Z. RNA-seq analyses of Arabidopsis thaliana seedlings after exposure to blue-light phototropic stimuli in microgravity. Am. J. Bot. 2019, 106, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Soga, K.; Wakabayashi, K.; Kamisaka, S.; Hoson, T. Stimulation of elongation growth and xyloglucan breakdown in Arabidopsis hypocotyls under microgravity conditions in space. Planta 2002, 215, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, N.; Benske, A.; Betz, H.; Schuetz, M.; Samuels, A.L. Laccases and Peroxidases Co-Localize in Lignified Secondary Cell Walls throughout Stem Development. Plant Physiol. 2020, 184, 806–822. [Google Scholar] [CrossRef]
- Irshad, M.; Canut, H.; Borderies, G.; Pont-Lezica, R.; Jamet, E. A new picture of cell wall protein dynamics in elongating cells of Arabidopsis thaliana: Confirmed actors and newcomers. BMC Plant Biol. 2008, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, T.; Lehotai, N.; Strand, Å. The role of retrograde signals during plant stress responses. J. Exp. Bot. 2017, 69, 2783–2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Zhang, A.; Lu, Q.; Kuang, T.; Lu, C.; Wen, X. Characterization of photosystem I in rice (Oryza sativa L.) seedlings upon exposure to random positioning machine. Photosynth. Res. 2013, 116, 93–105. [Google Scholar] [CrossRef]
- Castillon, A.; Shen, H.; Huq, E. Phytochrome Interacting Factors: Central players in phytochrome-mediated light signaling networks. Trends Plant Sci. 2007, 12, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.N.; Kathare, P.K.; Huq, E. Phytochromes and Phytochrome Interacting Factors. Plant Physiol. 2017, 176, 1025–1038. [Google Scholar] [CrossRef] [Green Version]
- Huq, E.; Al-Sady, B.; Hudson, M.; Kim, C.; Apel, K.; Quail, P.H. Phytochrome-interacting factor 1 is a critical bHLH: Regulator of chlorophyll biosynthesis. Science 2004, 305, 1937–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sineshchekov, V.A.; Belyaeva, O.B. Regulation of Chlorophyll Biogenesis by Phytochrome A; Pleiades Publishing: New York, NY, USA, 2019; Volume 84, pp. 491–508. [Google Scholar]
- Zhang, Y.; Mayba, O.; Pfeiffer, A.; Shi, H.; Tepperman, J.M.; Speed, T.P.; Quail, P.H. A Quartet of PIF bHLH Factors Provides a Transcriptionally Centered Signaling Hub That Regulates Seedling Morphogenesis through Differential Expression-Patterning of Shared Target Genes in Arabidopsis. PLoS Genet. 2013, 9, e1003244. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.-L.; Xue, H.-W. Global Analysis of Expression Profiles of Rice Receptor-Like Kinase Genes. Mol. Plant 2012, 5, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Osakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Sensing the environment: Key roles of membrane-localized kinases in plant perception and response to abiotic stress. J. Exp. Bot. 2013, 64, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Dievart, A.; Gottin, C.; Périn, C.; Ranwez, V.; Chantret, N. Origin and Diversity of Plant Receptor-Like Kinases. Annu. Rev. Plant Biol. 2020, 71, 131–156. [Google Scholar] [CrossRef] [Green Version]
- Jost, A.-I.K.; Hoson, T.; Iversen, T.-H. The Utilization of Plant Facilities on the International Space Station—The Composition, Growth, and Development of Plant Cell Walls under Microgravity Conditions. Plants 2015, 4, 44–62. [Google Scholar] [CrossRef]
- Poovaiah, B.W.; Yang, T.; van Loon, J.J. Calcium/calmodulin-mediated gravitropic response in plants. J. Gravit. Physiol. 2002, 9, P211–P214. [Google Scholar] [PubMed]
- Kumar, K.; Raina, S.K.; Sultan, S.M. Arabidopsis MAPK signaling pathways and their cross talks in abiotic stress response. J. Plant Biochem. Biotechnol. 2020, 29, 700–714. [Google Scholar] [CrossRef]
- Liu, Y.; He, C. A review of redox signaling and the control of MAP kinase pathway in plants. Redox Biol. 2017, 11, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, Y.; Yang, T.; Zhang, L.; Xu, S.; Xue, L.; An, L. Diverse signals converge at MAPK cascades in plant. Plant Physiol. Biochem. 2006, 44, 274–283. [Google Scholar] [CrossRef]
- Kamal, K.Y.; Herranz, R.; van Loon, J.J.W.A.; Medina, F.J. Simulated microgravity, Mars gravity, and 2g hypergravity affect cell cycle regulation, ribosome biogenesis, and epigenetics in Arabidopsis cell cultures. Sci. Rep. 2018, 8, 6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matía, I.; González-Camacho, F.; Herranz, R.; Kiss, J.Z.; Gasset, G.; van Loon, J.J.; Marco, R.; Javier Medina, F. Plant cell proliferation and growth are altered by microgravity conditions in spaceflight. J. Plant Physiol. 2010, 167, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Lan, P.; Li, W.; Lin, W.D.; Santi, S.; Schmidt, W. Mapping gene activity of Arabidopsis root hairs. Genome Biol. 2013, 14, R67. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.; Chandran, A.K.N.; An, G.; Lee, C.; Jung, K.H. Genome-wide analysis of root hair-preferential genes in rice. Rice 2018, 11, 48. [Google Scholar] [CrossRef]
- Ganguly, A.; Lee, S.H.; Cho, M.; Lee, O.R.; Yoo, H.; Cho, H.T. Differential auxin-transporting activities of PIN-FORMED proteins in Arabidopsis root hair cells. Plant Physiol. 2010, 153, 1046–1061. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Yan, F.; Zourelidou, M.; Wang, M.; Ljung, K.; Fastner, A.; Hammes, U.Z.; Di Donato, M.; Geisler, M.; Schwechheimer, C.; et al. SHADE AVOIDANCE 4 Is Required for Proper Auxin Distribution in the Hypocotyl. Plant Physiol. 2017, 173, 788–800. [Google Scholar] [CrossRef] [Green Version]
- Villaécija-Aguilar, J.A.; Körösy, C.; Maisch, L.; Hamon-Josse, M.; Petrich, A.; Magosch, S.; Chapman, P.; Bennett, T.; Gutjahr, C. KAI2 promotes Arabidopsis root hair elongation at low external phosphate by controlling local accumulation of AUX1 and PIN2. Curr. Biol. 2022, 32, 228–236.e223. [Google Scholar] [CrossRef]
- Stéger, A.; Palmgren, M. Root hair growth from the pH point of view. Front. Plant Sci. 2022, 13, 949672. [Google Scholar] [CrossRef]
- Eljebbawi, A.; Guerrero, Y.; Dunand, C.; Estevez, J.M. Highlighting reactive oxygen species as multitaskers in root development. iScience 2021, 24, 101978. [Google Scholar] [CrossRef]
- Shibata, M.; Sugimoto, K. A gene regulatory network for root hair development. J. Plant Res. 2019, 132, 301–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsson, A.; Karlsson, C.; Iversen, T.-H.; Chapman, D.K. Random root movements in weightlessness. Physiol. Plant. 1996, 96, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Levine, L.H.; Heyenga, A.G.; Levine, H.G.; Choi, J.W.; Davin, L.B.; Krikorian, A.D.; Lewis, N.G. Cell-wall architecture and lignin composition of wheat developed in a microgravity environment. Phytochemistry 2001, 57, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Sathasivam, M.; Hosamani, R.; Swamy, B.K.; Kumaran, G.S. Plant responses to real and simulated microgravity. Life Sci. Space Res. 2021, 28, 74–86. [Google Scholar] [CrossRef]
- Levine, H.G.; Anderson, K.; Boody, A.; Cox, D.; Kuznetsov, O.A.; Hasenstein, K.H. Germination and elongation of flax in microgravity. Adv. Space Res. 2003, 31, 2261–2268. [Google Scholar] [CrossRef]
- Levine, H.G.; Krikorian, A.D. Enhanced root production in Haplopappus gracilis grown under spaceflight conditions. J. Gravit. Physiol. 1996, 3, 17–27. [Google Scholar] [PubMed]
- Levine, H.G.; Piastuch, W.C. Growth patterns for etiolated soybeans germinated under spaceflight conditions. Adv. Space Res. 2005, 36, 1237–1243. [Google Scholar] [CrossRef]
- Marowa, P.; Ding, A.; Kong, Y. Expansins: Roles in plant growth and potential applications in crop improvement. Plant Cell Rep. 2016, 35, 949–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.B.; Lu, S.M.; Zhang, J.F.; Liu, S.; Lu, Y.T. A xyloglucan endotransglucosylase/hydrolase involves in growth of primary root and alters the deposition of cellulose in Arabidopsis. Planta 2007, 226, 1547–1560. [Google Scholar] [CrossRef]
- Tyler, L.; Lee, S.J.; Young, N.D.; DeIulio, G.A.; Benavente, E.; Reagon, M.; Sysopha, J.; Baldini, R.M.; Troìa, A.; Hazen, S.P.; et al. Population Structure in the Model Grass Brachypodium distachyon Is Highly Correlated with Flowering Differences across Broad Geographic Areas. Plant Genome 2016, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemura, K.; Kamachi, H.; Kume, A.; Fujita, T.; Karahara, I.; Hanba, Y.T. A hypergravity environment increases chloroplast size, photosynthesis, and plant growth in the moss Physcomitrella patens. J. Plant Res. 2017, 130, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Wen, Q.; Yu, R.; Han, X.; Wang Deng, X.; Chen, H. Light modulates the gravitropic responses through organ-specific PIFs and HY5 regulation of LAZY4 expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 18840–18848. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, S.-H.; Levine, H.G.; Masson, P.H. Brachypodium distachyon Seedlings Display Accession-Specific Morphological and Transcriptomic Responses to the Microgravity Environment of the International Space Station. Life 2023, 13, 626. https://doi.org/10.3390/life13030626
Su S-H, Levine HG, Masson PH. Brachypodium distachyon Seedlings Display Accession-Specific Morphological and Transcriptomic Responses to the Microgravity Environment of the International Space Station. Life. 2023; 13(3):626. https://doi.org/10.3390/life13030626
Chicago/Turabian StyleSu, Shih-Heng, Howard G. Levine, and Patrick H. Masson. 2023. "Brachypodium distachyon Seedlings Display Accession-Specific Morphological and Transcriptomic Responses to the Microgravity Environment of the International Space Station" Life 13, no. 3: 626. https://doi.org/10.3390/life13030626