

Testing the Single and Combined Effect of Kaolin and Spinosad against Bactrocera oleae and Its Natural Antagonist Insects in an Organic Olive Grove
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Identification and Population Dynamic Monitoring of Naturally Occurring B. oleae Egg/Larval Parasitoids
2.3. Statistical Analysis
3. Results and Discussion
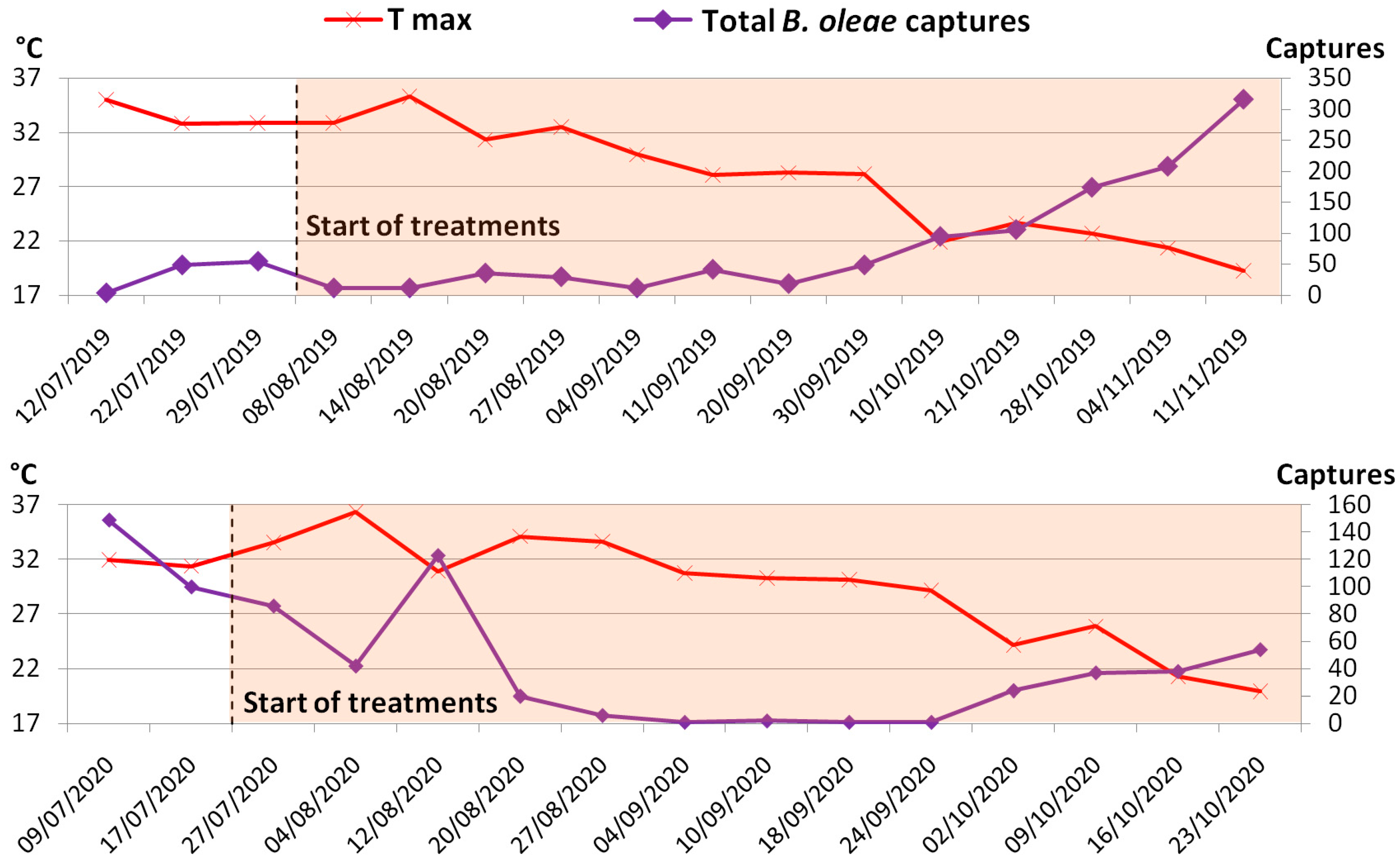
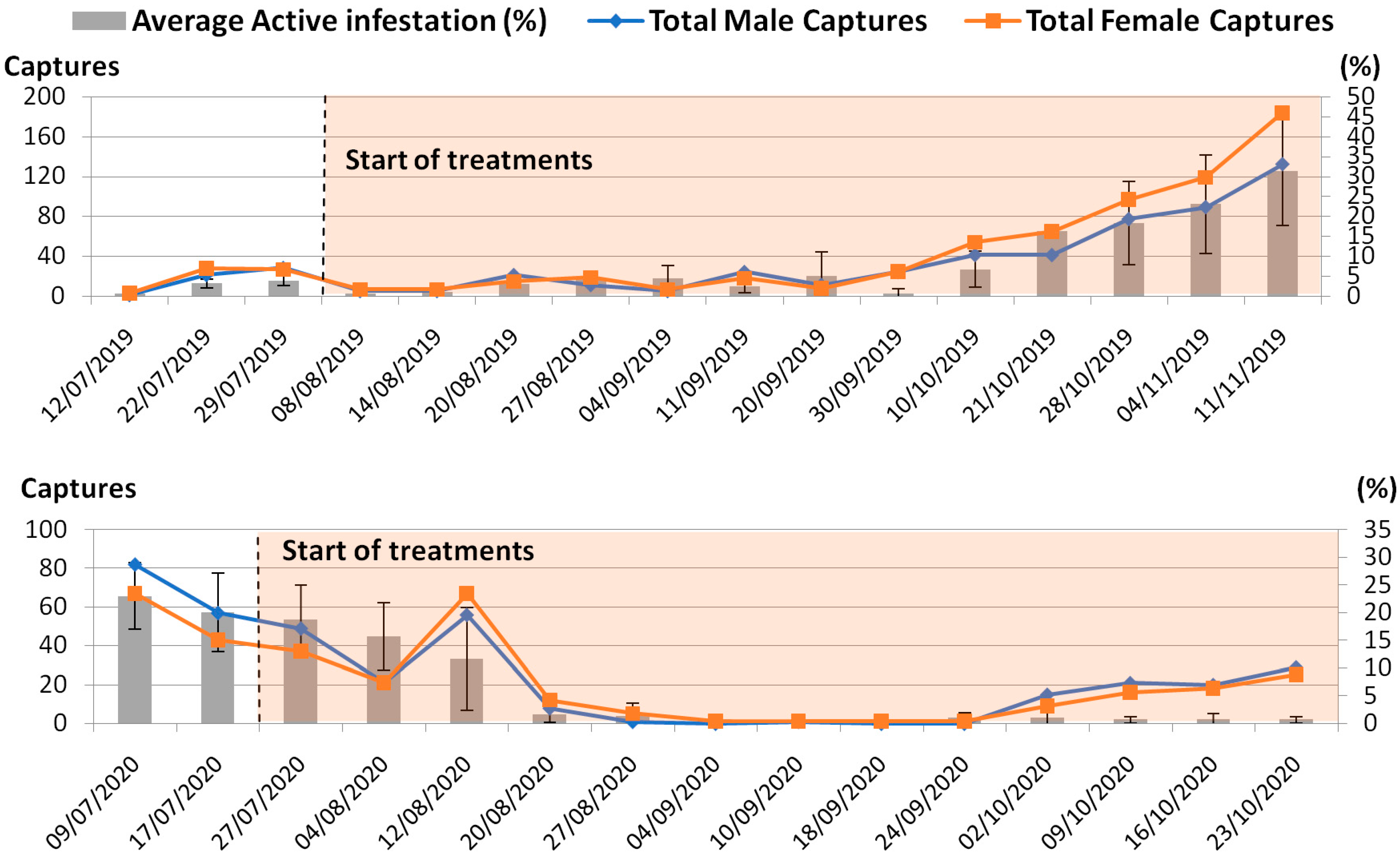
3.1. B. oleae Population Dynamics
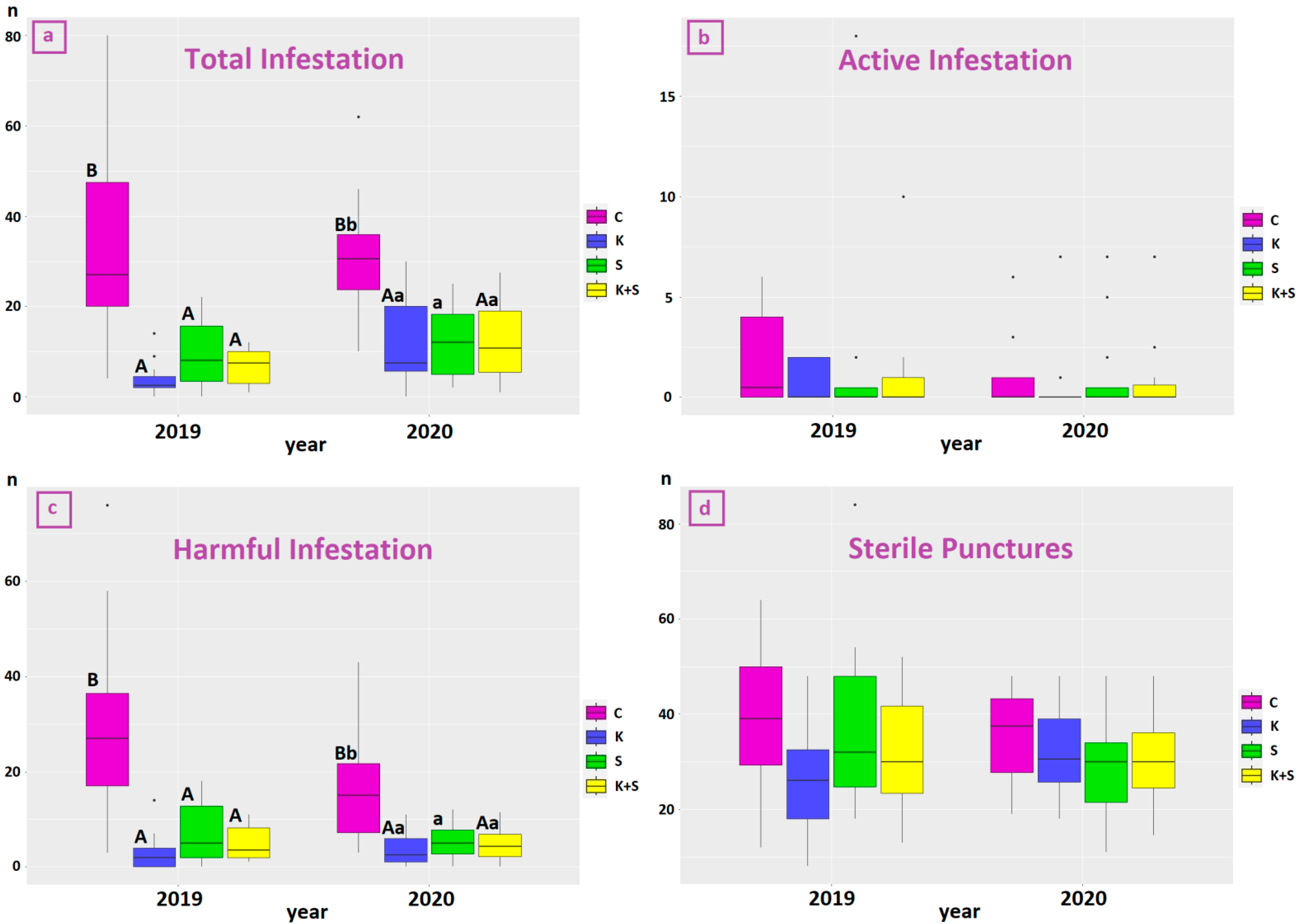
3.2. Effect of Treatment on Drupes’ Infestation Level and B. oleae Captures
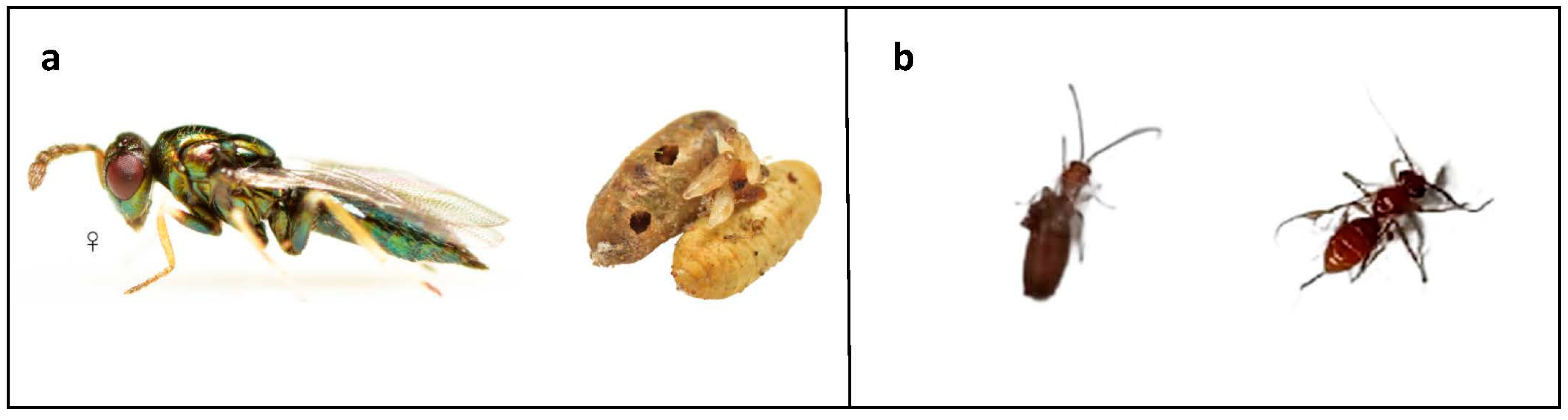
3.3. Identification and Monitoring of Naturally Occurring Parasitoids of B. oleae
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Daane, K.M.; Johnson, M.W. Olive fruit fly: Managing an ancient pest in modern times. Annu. Rev. Entomol. 2010, 55, 151–169. [Google Scholar] [CrossRef]
- Gómez-Caravaca, A.M.; Cerretani, L.; Bendini, A.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Del Carlo, M.; Compagnone, D.; Cichelli, A. Effects of fly attack (Bactrocera oleae) on the phenolic profile and selected chemical parameters of olive oil. J. Agric. Food Chem. 2008, 56, 4577–4583. [Google Scholar] [CrossRef]
- Al-Salti, M.N.; Edriss, O.; Al-Ali, M. Susceptibility of two olive varieties Aldeibli and Alkhudairi to olive fruit fly Bactrocera oleae (Gmelin)(Diptera: Tephritidae). J. Agric. Sci. Technol. 2011, 1, 987–996. [Google Scholar]
- Allmuça, F.; Dano, S.; Uka, R. Olive fly (Bactrocera oleae Gmelin) management with new allowed formulations, satisfying the olive oil quality requirements. Int. Refereed J. Eng. Sci. 2013, 2, 13. [Google Scholar]
- Nardi, F.; Carapelli, A.; Boore, J.L.; Roderick, G.K.; Dallai, R.; Frati, F. Domestication of olive fly through a multi-regional host shift to cultivated olives: Comparative dating using complete mitochondrial genomes. Mol. Phylogenetics Evol. 2010, 57, 678–686. [Google Scholar] [CrossRef]
- Sieuve, M. Mémoire et Journal D’observations sur les Moyens de Garantir les Olives de la Piqûre des Insectes et Nouvelle Méthode Pour enExtraireL’huile plus Abondante par L’invention d’un Moulin Domestique avec la Manière de la Garantir de TouteRancissure, par Sieve de Marseille; Paris tn-ia; Ace Books: New York, NY, USA, 1769. [Google Scholar]
- Grimaldi, D. Istruzioni Sulla Nuova Manifatturadell’olioIntrodottanella Calabria dal Marchese Domenico Grimaldi di Messimeri, Patrizio Genovese. Raffaele Lanciano, Napoli 1773, cap. V; Vincenzo Orsino: Philadelphia, PA, USA, 1777. [Google Scholar]
- Delrio, G. La diatribafra f. silvestri e a. berlesesulla lotta naturale e artificialecontro la moscadelle olive ed evoluzione di questetecniche di lotta. Atti Accad. Naz. Ital. Di Entomol. Anno LXII 2014, 81–87. [Google Scholar]
- Spanedda, A.F.; Terrosi, A. A field method for assessing the harmfulness to olive tree entomofauna of pesticides used for olive fly control. In IV International Symposium on Olive Growing; ISHS: Valenzano, Italy, 2000; Volume 586, pp. 849–852. [Google Scholar]
- Spanedda, A.F.; Terrosi, A. Toxic residue patterns in olive fruit, oil, and waste water of the most common insecticides used for controlling olive fly in Central Italy. In IV International Symposium on Olive Growing; ISHS: Valenzano, Italy, 2000; Volume 586, pp. 853–856. [Google Scholar]
- Sinab. La La Filiera Olivicola Italiana Superfici, Operatori e Produzione. 2020. Available online: https://www.sinab.it/sites/default/files/La%20filiera%20olivicola%20italiana.%20%20Superfici%2C%20operatori%20e%20produzione%20-%20R.%20Callieris.pdf (accessed on 11 December 2022).
- Lombardo, L.; Farolfi, C.; Capri, E. Sustainability certification, a new path of value creation in the olive oil sector: The ITALIAN case study. Foods 2021, 10, 501. [Google Scholar] [CrossRef]
- Lombardo, L.; Farolfi, C.; Capri, E. Development of a sustainability technical guide for the Italian olive oil supply chain. Sci. Total Environ. 2021, 820, 153332. [Google Scholar] [CrossRef]
- European Commission. Potential Certification and Labelling as Organic of Oils Extracted from Olives. 2019. Available online: https://www.ccpb.it/wp-content/uploads/2019/07/2019-07-04-Potential-certification-and-labelling-as-organic-of-oils-extracted-from-olives.pdf (accessed on 5 December 2022).
- Barry, J.D.; Sciarappa, W.J.; Teixeira, L.A.; Polavarapu, S. Comparative effectiveness of different insecticides for organic management of blueberry maggot (Diptera: Tephritidae). J. Econ. Entomol. 2005, 98, 1236–1241. [Google Scholar] [CrossRef]
- Skouras, P.J.; Margaritopoulos, J.T.; Seraphides, N.A.; Ioannides, I.M.; Kakani, E.G.; Mathiopoulos, K.D.; Tsitsipis, J.A. Organophosphate resistance in olive fruit fly, Bactrocera oleae, populations in Greece and Cyprus. Pest Manag. Sci. 2007, 63, 42–48. [Google Scholar] [CrossRef]
- Sanon, A.; Ba, M.; Dabire-Binso, C.; Pittendrigh, B. Effectiveness of Spinosad (Naturalytes) in Controlling the Cowpea Storage Pest, Callosobruchus maculatus (Coleoptera: Bruchidae). J. Econ. Entomol. 2010, 103, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.F.; Santos, S.A.P.; Torres, L.M. Efficacy of spinosad bait sprays to control Bactrocera oleae and impact on non-target arthropods. Phytoparasitica 2012, 40, 17–28. [Google Scholar] [CrossRef]
- Pascual, S.; Cobos, G.; Seris, E.; Sánchez-Ramos, I.; González-Núñez, M. Spinosad bait sprays against the olive fruit fly (Bactrocera oleae (Rossi)): Effect on the canopy non-target arthropod fauna. Int. J. Pest Manag. 2014, 60, 258–268. [Google Scholar] [CrossRef]
- Glenn, D.M.; Puterka, G.J.; Vanderzwet, T.; Byers, R.E.; Feldhake, C. Hydrophobic particle films: A new paradigm for suppression of arthropod pests and plant diseases. J. Econ. Entomol. 1999, 92, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Puterka, G.J.; Glenn, D.M.; Sekutowski, D.G.; Unruh, T.R.; Jones, S.K. Progress toward liquid formulations of particle films for insect and disease control in pear. Environ. Entomol. 2000, 29, 329–339. [Google Scholar] [CrossRef]
- Saour, G.; Makee, H. A kaolin-based particle film for suppression of olive fruit fly Bactrocera oleae Gmelin (Dipt.; Tephritidae) in olive groves. J. Appl. Entomol. 2004, 128, 28–31. [Google Scholar] [CrossRef]
- Pascual, S.; Cobos, G.; Seris, E.; González-Núñez, M. Effects of processed kaolin on pests and non-target arthropods in a Spanish olive grove. J. Pest Sci. 2009, 83, 121–133. [Google Scholar] [CrossRef]
- Gharbi, N.; Abdallah, S.B. Effectiveness of Kaolin Treatment for the Control of the Olive Fruit Fly Bactrocera oleae in Tunisian Olive Groves. Tunis. J. Plant Prot. 2016, 11, 73–81. [Google Scholar]
- González-Núñez, M.; Pascual, S.; Cobo, A.; Seris, E.; Cobos, G.; Fernández, C.E.; Sánchez-Ramos, I. Copper and kaolin sprays as tools for controlling the olive fruit fly. Entomol. Gen. 2021, 41, 97–110. [Google Scholar] [CrossRef]
- Santilli, E.; Lombardo, L.; Varlaro, M.E.; Nannelli, R.; Gagnarli, E.; BriccoliBati, C. Effectiveness of the GAEC cross-compliance Standard “Maintenance of olive groves in good vegetative condition” in avoiding the deterioration of habitats and land abandonment. Ital. J. Agron. 2011, 6, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Iannotta, N.; Perri, L.; Tocci, C.; Zaffina, F. The behaviour of different olive cultivars following attacks by Bactrocera oleae (Gmel.). Acta Hortic. 1999, 474, 545–548. [Google Scholar] [CrossRef]
- Malheiro, R.; Casal, S.; Pinheiro, L.; Baptista, P.; Pereira, J. Olive cultivar and maturation process on the oviposition preference of Bactrocera oleae (Rossi) (Diptera: Tephritidae). Bull. Entomol. Res. 2019, 109, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, L.; Fila, G.; Lombardo, N.; Epifani, C.; Duffy, D.H., III; Godino, G.; Salimonti, A.; Zelasco, S. Uncovering Olive Biodiversity through Analysis of Floral and Fruiting Biology and Assessment of Genetic Diversity of 120 Italian Cultivars with Minor or Marginal Diffusion. Biology 2019, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, R.; Caleca, V.; Lombardo, A. Relation of fruit color, elongation, hardness, and volume to the infestation of olive cultivars by the olive fruit fly, Bactrocera oleae. Entomol. Exp. Appl. 2012, 145, 15–22. [Google Scholar] [CrossRef]
- Pannelli, G.; Perri, E. ScelteVarietali in Olivicoltura. Collana Divulgativa dell’Accademia Volume XVIII; Accademia Nazionale dell’ Olivo e dell’ Olio Spoleto: Spoleto, Italy, 2012; pp. 1–26. ISSN 2281-4930. [Google Scholar]
- Ruiz-Torres, N.I.; Madueño-Magdaleno, C.; Montiel-Bueno, A. Efficacy of ground application with Spinosadand Imidacloprid bait sprays to control olive fruit fly (Bactrocera oleae). Preliminary results. Boletín Sanid. Veg. Plagas 2004, 30, 415–425. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Genç, H.; Nation, J.L. Survival and development of Bactrocera oleae Gmelin (Diptera:Tephritidae) immature stages at four temperatures in the laboratory. Afr. J. Biotechnol. 2008, 7, 2495–2500. [Google Scholar]
- Rizzo, R.; Caruso, T.; Caleca, V. Control of Bactrocera oleae (Rossi) in organic olive orchards: Use of clays and spinosad-based bait. IOBC/WPRS Bull. 2019, 41, 67–70. [Google Scholar]
- Perri, E.; Iannotta, N.; Muzzalupo, I.; Russo, A.; Caravita, M.A.; Pellegrino, M.; Parise, A.; Tucci, P. Kaolin protects olive fruits from Bactrocera oleae (Gmelin) infestations unaffecting olive oil quality. IOBC/WPRS Bull. 2006, 30, 153. [Google Scholar]
- Pennino, G.; Pane, G.; Raiti, G.; Perri, E.; Carovita, M.A.; Macchione, B.; Tucci, P.; Socievole, P.; Pellegrino, M.; Cartabellotta, D.; et al. Three years field trials to assess the effect of kaolin made particles and copper on olive-fruit fly (B. oleae Gmelin) infestations in Sicily. Second International Seminar “Biotechnology and Quality of Olive tree Products around the Mediterranean Basin”. In Proceedings of the DCA–Università di Palermo; Regione Siciliana–Assessorato Agricoltura e Foreste, II, Marsala, Italy, 5–10 November 2006; pp. 303–306. Available online: http://orgprints.org/14278/ (accessed on 8 June 2010).
- Randazzo, C.L.; Fava, G.; Tomaselli, F.; Romeo, F.V.; Pennino, G.; Vitello, E.; Caggia, C. Effect of kaolin and copper based products and of starter cultures on green table olive fermentation. Food Microbiol. 2011, 28, 910–919. [Google Scholar] [CrossRef]
- Kakani, E.G.; Zygouridis, N.E.; Tsoumani, K.T.; Seraphides, N.; Zalom, F.G.; Mathiopoulos, K.D. Spinosad resistance development in wild olive fruit fly Bactrocera oleae (Diptera: Tephritidae) populations in California. Pest Manag. Sci. 2010, 66, 447–453. [Google Scholar] [CrossRef]
- Mayes, M.; Thompson, G.; Husband, B.; Miles, M. Spinosad Toxicity to Pollinators and Associated Risk. Rev. Environ. Contam. Toxicol. 2003, 179, 37–71. [Google Scholar] [CrossRef] [PubMed]
- Miles, M. The effects of spinosad, a naturally derived insect control agent to the honeybee. Bull. Insectology 2003, 56, 119–124. [Google Scholar]
- Miles, M. The effects of spinosad on beneficial insects and mites used in integrated pest management systems in greenhouses. pesticide and Beneficial Organisms. IOBC/WRPS Bull. 2006, 29, 53–59. [Google Scholar]
- Markó, V.; Bogya, S.; Kondorosy, E.; Blommers, L. Side effects of kaolin particle films on apple orchard bug, beetle and spider communities. Int. J. Pest Manag. 2010, 56, 189–199. [Google Scholar] [CrossRef]
- Sánchez-Ramos, I.; Marcotegui, A.; Pascual, S.; Fernández, C.E.; Cobos, G.; González-Núñez, M. Compatibility of organic farming treatments against Monosteira unicostata with non-target arthropod fauna of almond trees canopy. Span. J. Agric. Res. 2017, 15, e1004. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, G.; Perrino, E.V.; Ladisa, G.; Aly, A.; Tesfmichael Solomon, M.; Mazdaric, S.; Benedetti, A.; Ceglie, F.G. Short-term effects of different soil management practices on biodiversity and soil quality of Mediterranean ancient olive orchards. Org. Agr. 2015, 5, 209–223. [Google Scholar] [CrossRef]
- Lombardo, L.; Palese, A.M.; Grasso, F.; Duffy, D.H., III; BriccoliBati, C.; Xiloyannis, C. Mechanical Tillage Diversely Affects Glomalin Content, Water Stable Aggregates and AM Fungal Community in the Soil Profiles of Two Differently Managed Olive Orchards. Biomolecules 2019, 9, 639. [Google Scholar] [CrossRef] [Green Version]
- García-Ruiz, R.; Ochoa, V.; Viñegla, B.; Hinojosa, M.B.; Peña-Santiago, R.; Liébanas, G.; Linares, J.C.; Carreira, J.A. Soil enzymes, nematode community and selected physico-chemical properties as soil quality indicators in organic and conventional olive oil farming: Influence of seasonality and site features. Appl. Soil Ecol. 2009, 41, 305–314. [Google Scholar] [CrossRef]
- Vizzarri, V.; Novellis, C.; Rizzo, P. First report of Baryscapus silvestrii in Calabria, Italy (Chalcidoidea Eulophidae). Boll. Della Soc. Entomol. Ital. 2020, 152, 75–78. [Google Scholar] [CrossRef]
- Ras, E.; Beukeboom, L.W.; Caceres, C.; Bourtzis, K. Review of the role of gut microbiota in mass rearing of the olive fruit fly, Bactrocera oleae, and its parasitoids. Entomol. Exp. Appl. 2017, 164, 237–256. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, L.A.; Dáder, B.; Wanumen, A.C.; Pereira, J.A.; Santos, S.A.P.; Medina, P. Side Effects of Pesticides on the Olive Fruit Fly Parasitoid Psyttalia concolor (Szépligeti): A Review. Agronomy 2020, 10, 1755. [Google Scholar] [CrossRef]
- Bengochea, P.; Budia, F.; Viñuela, E.; Medina, P. Are kaolin and copper treatments safe to the olive fruit fly parasitoid Psyttalia concolor? J. Pest Sci. 2014, 87, 351–359. [Google Scholar] [CrossRef]
- Bengochea, P.; Hernando, S.; Saelices, R.; Adán, A.; Budia, F.; González-Núñez, M.; Viñuela, E.; Medina, P. Side effects of kaolin on natural enemies found on olive crops. Pestic. Benef. Org. IOBC/WPRS Bull. 2010, 55, 61–67. [Google Scholar]
- Bengochea, P.; Amor, F.; Saelices, R.; Hernando, S.; Budia, F.; Adán, A.; Medina, P. Kaolin and copper-based products applications: Ecotoxicology on four natural enemies. Chemosphere 2013, 91, 1189–1195. [Google Scholar] [CrossRef]
- Williams, T.; Javier Valle, J.; Viñuela, E. Is the naturally derived insecticide spinosad® compatible with insect natural enemies? Biocontrol Sci. Technol. 2003, 13, 459–475. [Google Scholar] [CrossRef]
- Biondi, A.; Mommaerts, V.; Smagghe, G.; Viñuela, E.; Zappalà, L.; Desneux, N. The non-target impact of spinosyns on beneficial arthropods. Pest Manag. Sci. 2012, 68, 1523–1536. [Google Scholar] [CrossRef]
- Tello, V.; Díaz, L.; Sánchez, M. Side effects of the natural pesticide Spinosad (GF-120 Formulation) on Eretmocerus paulistus (Hymenoptera: Aphelinidae), a parasitoid of the whitefly Aleurothrixus floccosus (Hemiptera: Aleyrodidae), under laboratory conditions. Cienc. Investig. Agrar. 2013, 40, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Viñuela, E.; Medina, M.P.; Schneider, M.; González, M.; Budia, F.; Adán, A.; Del Estal, P. Comparison of side-effects of spinosad, tebufenozide and azadirachtin on the predators Chrysoperla carnea and Podisus maculiventris and the parasitoids Opius concolor and Hyposoter didymator under laboratory conditions. OILB/IOBC Bull. 2001, 24, 25–34. [Google Scholar]
- Moral, J.; Eldesouki-Arafat, I.; Lopez-Escudero, F.; Vargas-Osuna, E.; Trapero, A.; Aldebis, H. Olive escudete, caused by Botryosphaeria dothidea, as result of the interaction fly-mosquito-fungus. Phytopathology 2016, 106, S135. [Google Scholar]
- University of California. UC IPM Pest Management Guidelines; OLIVE; University of California: San Francisco, CA, USA, 2014; p. 71. [Google Scholar]
- Moral, J.; Morgan, D.; Trapero, A.; Michailides, T.J. Ecology and Epidemiology of Diseases of Nut Crops and Olives Caused by Botryosphaeriaceae Fungi in California and Spain. Plant Dis. 2019, 103, 1809–1827. [Google Scholar] [CrossRef] [Green Version]
- Lazzizera, C.; Frisullo, S.; Alves, A.; Phillips, A.J.L. Morphology, phylogeny and pathogenicity of Botryosphaeria and Neofusicoccum species associated with drupe rot of olives in southern Italy. Plant Pathol. 2008, 57, 948–956. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasitoid | Treatment | Total Individuals | % of Endoparasitism | % of Ectoparasitism | |||
|---|---|---|---|---|---|---|---|
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | ||
| Eurytoma martellii | C | 5 | 7 | 2.02 | 2.23 | ||
| K | 1 | 4 | 2.08 | 3.17 | |||
| S | 1 | 3 | 1.67 | 2.46 | |||
| K + S | 2 | 4 | 1.85 | 2.31 | |||
| Prolasioptera berlesiana | C | 9 | 11 | 3.03 | 3.36 | ||
| K | 4 | 5 | 4.17 | 3.17 | |||
| S | 3 | 6 | 3.33 | 3.28 | |||
| K + S | 3 | 6 | 3.70 | 3.84 | |||
| Eupelmus urozonus | C | 4 | 6 | 1.51 | 1.86 | ||
| K | 1 | 3 | 2.08 | 2.38 | |||
| S | 1 | 4 | 1.67 | 2.46 | |||
| K + S | 1 | 3 | 1.85 | 1.54 | |||
| Psyttalia concolor | C | 12 | 6 | 5.78 | 3.48 | ||
| K | 3 | 1 | 6.89 | 3.84 | |||
| S | 1 | 1 | 2.78 | 0 | |||
| K + S | 2 | 1 | 2.63 | 3.12 | |||
| Baryscapus sylvestrii | C | 8 | 6 | 4.04 | 4.65 | ||
| K | 1 | 3 | 3.44 | 3.84 | |||
| S | 2 | 2 | 2.77 | 2.5 | |||
| K + S | 2 | 1 | 5.26 | 3.12 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizzarri, V.; Lombardo, L.; Novellis, C.; Rizzo, P.; Pellegrino, M.; Cruceli, G.; Godino, G.; Zaffina, F.; Ienco, A. Testing the Single and Combined Effect of Kaolin and Spinosad against Bactrocera oleae and Its Natural Antagonist Insects in an Organic Olive Grove. Life 2023, 13, 607. https://doi.org/10.3390/life13030607
Vizzarri V, Lombardo L, Novellis C, Rizzo P, Pellegrino M, Cruceli G, Godino G, Zaffina F, Ienco A. Testing the Single and Combined Effect of Kaolin and Spinosad against Bactrocera oleae and Its Natural Antagonist Insects in an Organic Olive Grove. Life. 2023; 13(3):607. https://doi.org/10.3390/life13030607
Chicago/Turabian StyleVizzarri, Veronica, Luca Lombardo, Carmine Novellis, Pierluigi Rizzo, Massimiliano Pellegrino, Giuseppe Cruceli, Gianluca Godino, Francesco Zaffina, and Annamaria Ienco. 2023. "Testing the Single and Combined Effect of Kaolin and Spinosad against Bactrocera oleae and Its Natural Antagonist Insects in an Organic Olive Grove" Life 13, no. 3: 607. https://doi.org/10.3390/life13030607