
Proteomic Analysis of Female Synovial Fluid to Identify Novel Biomarkers for Osteoarthritis
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Protein Extraction, Digestion, and Liquid Chromatography with Tandem Mass Spectrometry (LC-MS-MS) LC-MS/MS Analysis
2.3. Statistical and Bioinformatics Analyses
3. Results
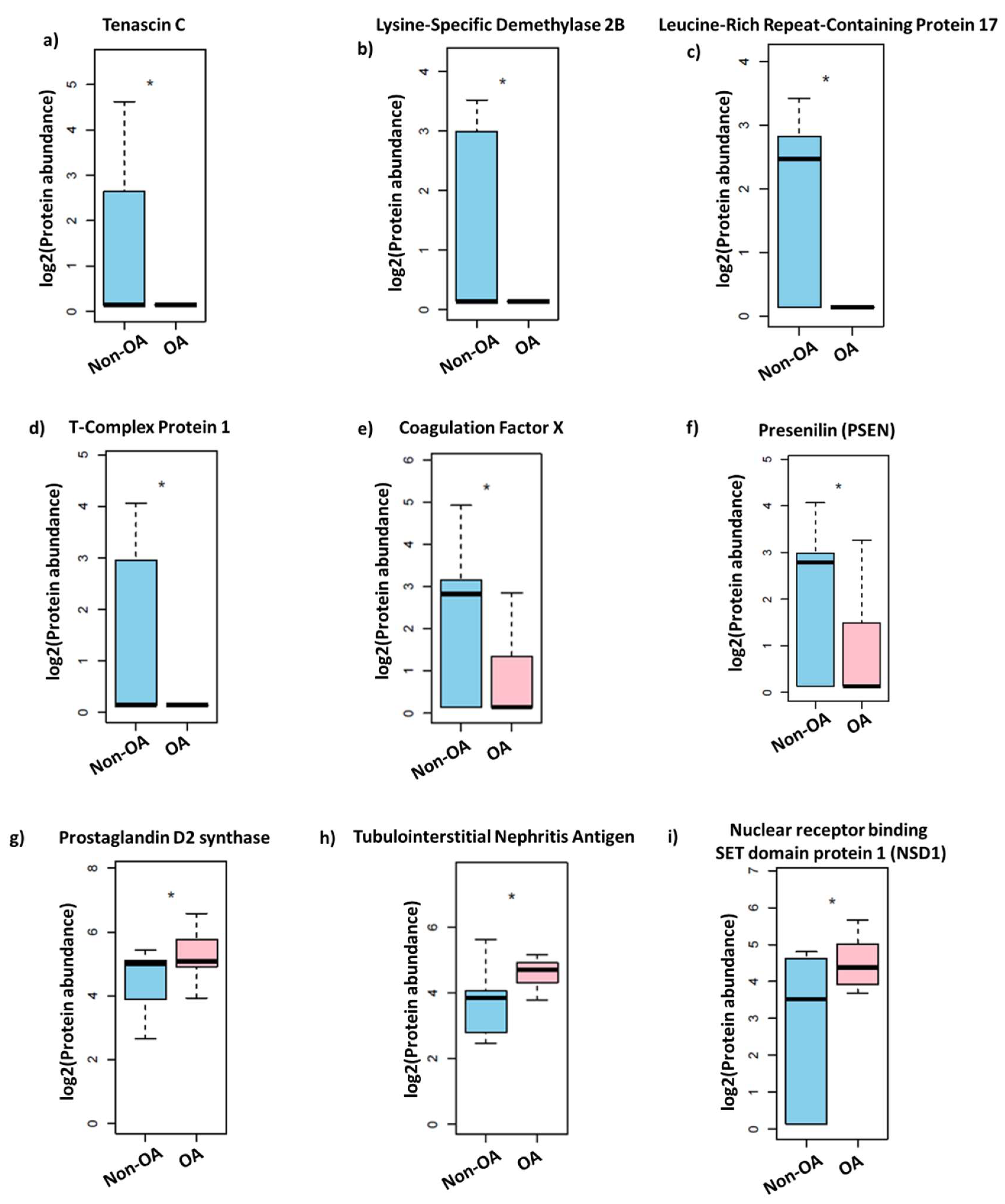
3.1. Synovial Fluid Protein Content in Women Differs Significantly from OA Patients
3.2. DAVID and QuickGO Analysis of Differentially Expressed Proteins
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zhang, Z.; Sheng, P.; Mobasheri, A. The role of metabolism in chondrocyte dysfunction and the progression of osteoarthritis. Ageing Res. Rev. 2021, 66, 101249. [Google Scholar] [CrossRef]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Deshpande, B.R.; Katz, J.N.; Solomon, D.H.; Yelin, E.H.; Hunter, D.J.; Messier, S.P.; Suter, L.G.; Losina, E. Number of Persons with Symptomatic Knee Osteoarthritis in the US: Impact of Race and Ethnicity, Age, Sex, and Obesity. Arthritis Care Res. 2016, 68, 1743–1750. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.D.; Golightly, Y.M. State of the evidence. Curr. Opin. Rheumatol. 2015, 27, 276–283. [Google Scholar] [CrossRef]
- Johnson, V.L.; Hunter, D.J. The epidemiology of osteoarthritis. Best Pract. Res. Clin. Rheumatol. 2014, 28, 5–15. [Google Scholar] [CrossRef]
- Debi, R.; Mor, A.; Segal, O.; Segal, G.; Debbi, E.; Agar, G.; Halperin, N.; Haim, A.; Elbaz, A. Differences in gait patterns, pain, function and quality of life between males and females with knee osteoarthritis: A clinical trial. BMC Musculoskelet. Disord. 2009, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Hanna, F.S.; Teichtahl, A.J.; Wluka, A.E.; Wang, Y.; Urquhart, D.M.; English, D.; Giles, G.; Cicuttini, F.M. Women have increased rates of cartilage loss and progression of cartilage defects at the knee than men: A gender study of adults without clinical knee osteoarthritis. Menopause 2009, 16, 666–670. [Google Scholar] [CrossRef]
- Palazzo, C.; Ravaud, J.F.; Papelard, A.; Ravaud, P.; Poiraudeau, S. The burden of musculoskeletal conditions. PLoS ONE 2014, 9, e90633. [Google Scholar] [CrossRef]
- GBD 2016 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259, Erratum in Lancet 2017, 390, e38. [Google Scholar] [CrossRef] [Green Version]
- Losina, E.; Walensky, R.P.; Kessler, C.L.; Emrani, P.S.; Reichmann, W.M.; Wright, E.A.; Holt, H.L.; Solomon, D.H.; Yelin, E.; Paltiel, A.D.; et al. Cost-effectiveness of total knee arthroplasty in the United States: Patient risk and hospital volume. Arch. Intern. Med. 2009, 169, 1113–1122. [Google Scholar] [CrossRef] [Green Version]
- Kotlarz, H.; Gunnarsson, C.L.; Fang, H.; Rizzo, J.A. Osteoarthritis and absenteeism costs: Evidence from US National Survey Data. J. Occup. Environ. Med. 2010, 52, 263–268. [Google Scholar] [CrossRef]
- Deyle, G.D.; Allen, C.S.; Allison, S.C.; Gill, N.W.; Hando, B.R.; Petersen, E.J.; Dusenberry, D.I.; Rhon, D.I. Physical Therapy versus Glucocorticoid Injection for Osteoarthritis of the Knee. N. Engl. J. Med. 2020, 382, 1420–1429. [Google Scholar] [CrossRef]
- Dziedzic, K.S.; Allen, K.D. Challenges and controversies of complex interventions in osteoarthritis management: Recognizing inappropriate and discordant care. Rheumatology 2018, 57 (Suppl. S4), iv88–iv98. [Google Scholar] [CrossRef] [Green Version]
- Kolhe, R.; Hunter, M.; Liu, S.; Jadeja, R.N.; Pundkar, C.; Mondal, A.K.; Mendhe, B.; Drewry, M.; Rojiani, M.V.; Liu, Y.; et al. Gender-specific differential expression of exosomal miRNA in synovial fluid of patients with osteoarthritis. Sci. Rep. 2017, 7, 2029. [Google Scholar] [CrossRef] [Green Version]
- Kolhe, R.; Owens, V.; Sharma, A.; Lee, T.J.; Zhi, W.; Ghilzai, U.; Mondal, A.K.; Liu, Y.; Isales, C.M.; Hamrick, M.W.; et al. Sex-Specific Differences in Extracellular Vesicle Protein Cargo in Synovial Fluid of Patients with Osteoarthritis. Life 2020, 10, 337. [Google Scholar] [CrossRef]
- Liu, P.; Ping, Y.; Ma, M.; Zhang, D.; Liu, C.; Zaidi, S.; Gao, S.; Ji, Y.; Lou, F.; Yu, F.; et al. Anabolic actions of Notch on mature bone. Proc. Natl. Acad. Sci. USA 2016, 113, E2152–E2161. [Google Scholar] [CrossRef] [Green Version]
- Mohan, S.; Hu, Y.; Edderkaoui, B. Identification of gender-specific candidate genes that influence bone microarchitecture in chromosome 1. Calcif. Tissue Int. 2013, 92, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, M.; Yoshida, T.; Sudo, A. Tenascin-C in Osteoarthritis and Rheumatoid Arthritis. Front. Immunol. 2020, 11, 577015. [Google Scholar] [CrossRef]
- Jakowlev, K.; Livshits, G.; Kalichman, L.; Ben-Asher, E.; Malkin, I.; Lancet, D.; Kobyliansky, E. Search for hand osteoarthritis susceptibility locus on chromosome 6p12.3-p12.1. Hum. Biol. 2007, 79, 1–14. [Google Scholar] [CrossRef]
- Li, M.; Zhi, L.; Zhang, Z.; Bian, W.; Qiu, Y. Identification of potential target genes associated with the pathogenesis of osteoarthritis using microarray based analysis. Mol. Med. Rep. 2017, 16, 2799–2806. [Google Scholar] [CrossRef] [Green Version]
- Najar, M.; Ouhaddi, Y.; Paré, F.; Lussier, B.; Urade, Y.; Kapoor, M.; Pelletier, J.; Martel-Pelletier, J.; Benderdour, M.; Fahmi, H. Role of Lipocalin-Type Prostaglandin D Synthase in Experimental Osteoarthritis. Arthritis Rheumatol. 2020, 72, 1524–1533. [Google Scholar] [CrossRef]
- Ouhaddi, Y.; Najar, M.; Paré, F.; Lussier, B.; Urade, Y.; Benderdour, M.; Pelletier, J.-P.; Martel-Pelletier, J.; Fahmi, H. L-PGDS deficiency accelerated the development of naturally occurring age-related osteoarthritis. Aging 2020, 12, 24778–24797. [Google Scholar] [CrossRef]
- Haworth, O.; Buckley, C.D. Resolving the problem of persistence in the switch from acute to chronic inflammation. Proc. Natl. Acad. Sci. USA 2007, 104, 20647–20648. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Wu, Z.S.; Xu, Q.; Wang, K.; Xu, M.-T.; Ha, C.-Z.; Zhang, C.; Wang, D.-W. LRRC17 regulates the bone metabolism of human bone marrow mesenchymal stem cells from patients with idiopathic necrosis of femoral head through Wnt signaling pathways: A preliminary report. Exp. Ther. Med. 2021, 22, 666. [Google Scholar] [CrossRef]
- Zhong, X.L.; Qian, B.P.; Huang, J.C.; Zhao, S.Z.; Li, Y.; Qiu, Y. Low expression of TCP1 (T-Complex 1) and PSMC1 (Proteasome 26S subunit, ATPase 1) in heterotopic ossification during ankylosing spondylitis. Bioengineered 2021, 12, 7459–7469. [Google Scholar] [CrossRef]
- van Haelst, M.M.; Hoogeboom, J.J.; Baujat, G.; Brüggenwirth, H.T.; Van De Laar, I.; Coleman, K.; Rahman, N.; Niermeijer, M.F.; Drop, S.L.; Scambler, P.J. Familial gigantism caused by an NSD1 mutation. Am. J. Med. Genet. A 2005, 139, 40–44. [Google Scholar] [CrossRef]
- Funck-Brentano, T.; Cohen-Solal, M. Subchondral bone and osteoarthritis. Curr. Opin. Rheumatol. 2015, 27, 420–426. [Google Scholar] [CrossRef]
- Nevitt, M.C.; Felson, D.T. High bone density and radiographic osteoarthritis: Questions answered and unanswered. Osteoarthr. Cartil. 2020, 28, 1151–1153. [Google Scholar] [CrossRef]
- Geusens, P.P.; van den Bergh, J.P. Osteoporosis and osteoarthritis: Shared mechanisms and epidemiology. Curr. Opin. Rheumatol. 2016, 28, 97–103. [Google Scholar] [CrossRef]



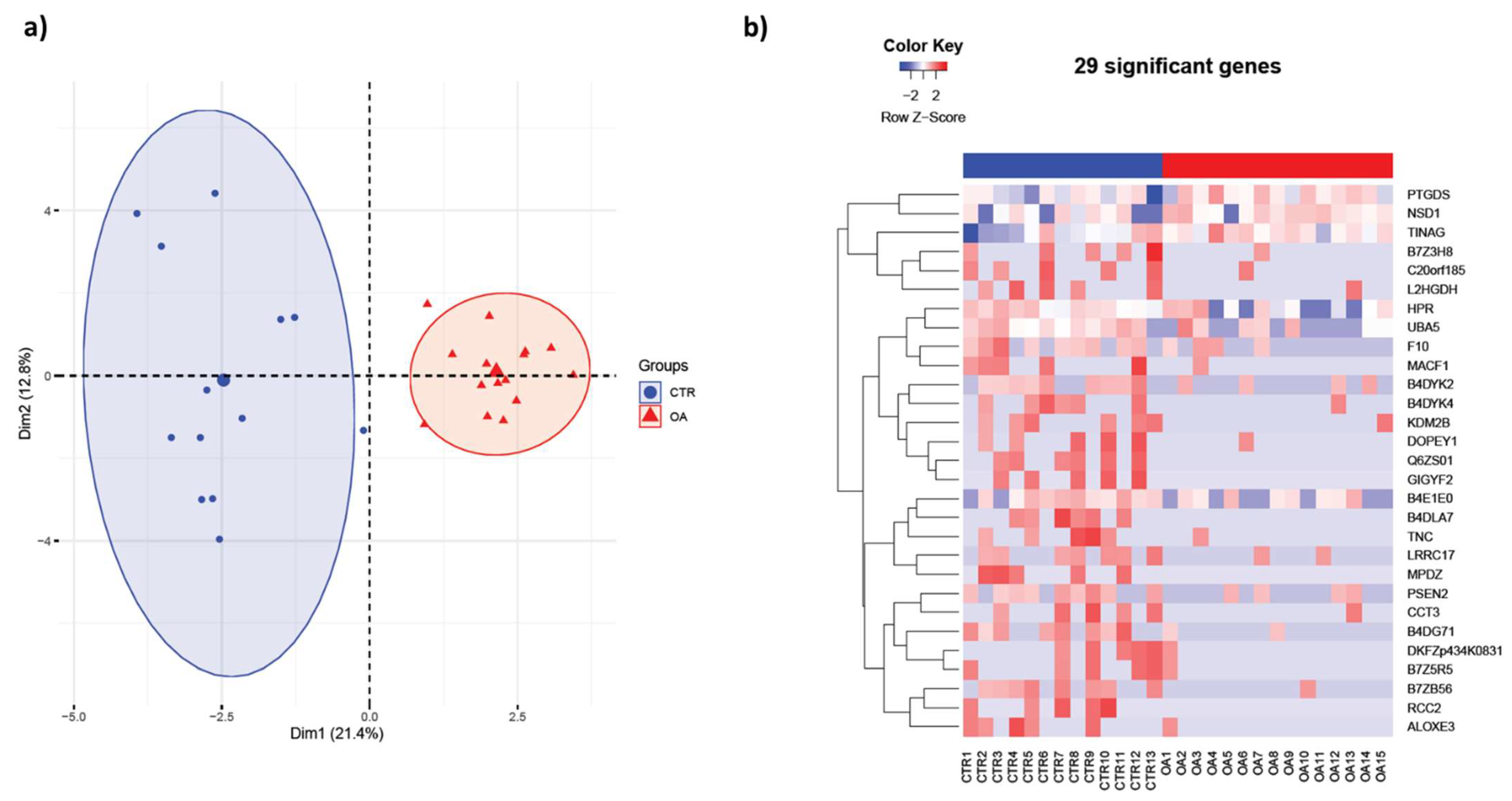{kind=link}
{kind=link}
| Accession | Description | Fold Change | p Value |
|---|---|---|---|
| B2RWP5 | Nuclear receptor binding SET domain protein 1 (NSD1) | 2.65 | 0.028 |
| A0A024R8G3 | Prostaglandin D2 synthase 21kDa (Brain), isoform CRA_a | 1.95 | 0.035 |
| Q5T466 | Tubulointerstitial Nephritis Antigen | 1.90 | 0.040 |
| J3KPH2 | Arachidonate lipoxygenase 3, isoform CRA_a | 0.53 | 0.030 |
| Q14DE0 | Chromosome 20 open reading frame 185 | 0.52 | 0.037 |
| B7Z5R5 | cDNA FLJ61652, highly similar to Mus musculus DEP domain containing 1a (Depdc1a), mRNA | 0.51 | 0.027 |
| C9JVN9 | L-2-hydroxyglutarate dehydrogenase, mitochondrial | 0.51 | 0.042 |
| Q69YG3 | Putative uncharacterized protein DKFZp434K0831 (Fragment) | 0.51 | 0.027 |
| B4DUR8 | T-Complex Protein 1 subunit gamma | 0.50 | 0.039 |
| I1E4Y6 | PERQ amino acid-rich with GYF domain-containing protein 2 | 0.49 | 0.006 |
| H0UI11 | Dopey family member 1, isoform CRA_a | 0.48 | 0.033 |
| A0A024R884 | Tenascin C (Hexabrachion), isoform CRA_a | 0.48 | 0.033 |
| B4DYK4 | cDNA FLJ56807, highly similar to Rab6-interacting protein 1 | 0.47 | 0.015 |
| C9JT74 | Leucine-rich repeat-containing protein 17 (Fragment) | 0.47 | 0.020 |
| H0YGQ3 | Multiple PDZ domain protein (Fragment) | 0.46 | 0.006 |
| A0A0C4DGG3 | Lysine-Specific Demethylase 2B | 0.45 | 0.018 |
| B7Z3H8 | cDNA FLJ57997, highly similar to Transmembrane GTPase MFN2 (EC 3.6.5.-) | 0.45 | 0.030 |
| H3BPE1 | Microtubule-actin cross-linking factor 1, isoforms 1/2/3/5 | 0.45 | 0.027 |
| A5PLK7 | RCC2 protein (Fragment) | 0.44 | 0.006 |
| B4DYK2 | cDNA FLJ56270, highly similar to Homo sapiens pleckstrin homology domain containing, family G (with RhoGef domain) member 2, mRNA | 0.43 | 0.028 |
| B4E1E0 | CTP synthase | 0.42 | 0.038 |
| Q6ZS01 | cDNA FLJ45938 fis, similar to Mus musculus zinc finger protein 292 (Zfp292) (Fragment) | 0.42 | 0.002 |
| E5RFW4 | Presenilin (Fragment) | 0.42 | 0.021 |
| Q5JVE7 | Coagulation Factor X | 0.40 | 0.025 |
| B4DLA7 | cDNA FLJ59923, highly similar to Cytohesin-1 | 0.39 | 0.002 |
| E7EWE1 | Ubiquitin-like modifier-activating enzyme 5 | 0.34 | 0.020 |
| B7ZB56 | cDNA, FLJ79420, highly similar to Homo sapiens nebulette (NEBL), transcript variant 2, mRNA | 0.33 | 0.001 |
| J3KTC3 | Haptoglobin-related protein | 0.32 | 0.025 |
| B4DG71 | cDNA FLJ56619, highly similar to cAMP-specific 3’,5’-cyclic phosphodiesterase4B (EC 3.1.4.17) | 0.26 | 0.002 |
| Gene Ontology | Term | Ont | p Value |
|---|---|---|---|
| GO:0006950 | response to stress | BP | 0.007218 |
| GO:0009611 | response to wounding | BP | 0.007153 |
| GO:0016043 | cellular component organization | BP | 0.024775 |
| GO:0019538 | protein metabolic process | BP | 0.013128 |
| GO:0030154 | cell differentiation | BP | 0.037616 |
| GO:0042060 | wound healing | BP | 0.004072 |
| GO:0048518 | positive regulation of biological process | BP | 0.00079 |
| GO:0048523 | negative regulation of cellular process | BP | 0.023211 |
| GO:0048869 | cellular developmental process | BP | 0.040836 |
| GO:0050789 | regulation of biological process | BP | 0.012056 |
| GO:0050794 | regulation of cellular process | BP | 0.005146 |
| GO:0065007 | biological regulation | BP | 0.021133 |
| GO:0071840 | cellular component organization or biogenesis | BP | 0.029521 |
| GO:1901564 | organonitrogen compound metabolic process | BP | 0.033099 |
| GO:0048522 | positive regulation of cellular process | BP | 0.000313 |
| GO:0030334 | regulation of cell migration | BP | 0.022019 |
| GO:0032879 | regulation of localization | BP | 0.025221 |
| GO:0040012 | regulation of locomotion | BP | 0.028507 |
| GO:0051270 | regulation of cellular component movement | BP | 0.031709 |
| GO:2000145 | regulation of cell motility | BP | 0.02594 |
| GO:0007155 | cell adhesion | BP | 0.010892 |
| GO:0022610 | biological adhesion | BP | 0.011072 |
| GO:0030155 | regulation of cell adhesion | BP | 0.009815 |
| GO:0051128 | regulation of cellular component organization | BP | 0.012686 |
| GO:0034330 | cell junction organization | BP | 0.008997 |
| GO:0040007 | growth | BP | 0.002564 |
| GO:0048589 | developmental growth | BP | 0.007582 |
| GO:0051276 | chromosome organization | BP | 0.040125 |
| GO:0012505 | endomembrane system | CC | 0.012368 |
| GO:0031982 | vesicle | CC | 0.027865 |
| GO:0043227 | membrane-bounded organelle | CC | 0.026887 |
| GO:0015630 | microtubule cytoskeleton | CC | 0.006241 |
| GO:0043228 | non-membrane-bounded organelle | CC | 0.043619 |
| GO:0043232 | intracellular non-membrane-bounded organelle | CC | 0.043272 |
| GO:0042995 | cell projection | CC | 0.049255 |
| GO:0043005 | neuron projection | CC | 0.049972 |
| GO:0005768 | endosome | CC | 0.020981 |
| GO:0005794 | Golgi apparatus | CC | 0.002081 |
| GO:0005783 | endoplasmic reticulum | CC | 0.009801 |
| GO:0030425 | dendrite | CC | 0.005927 |
| GO:0036477 | somatodendritic compartment | CC | 0.014925 |
| GO:0044297 | cell body | CC | 0.005378 |
| GO:0097447 | dendritic tree | CC | 0.005981 |
| GO:0099080 | supramolecular complex | CC | 0.000852 |
| GO:0099081 | supramolecular polymer | CC | 0.0027 |
| GO:0099512 | supramolecular fiber | CC | 0.002622 |
| GO:0099513 | polymeric cytoskeletal fiber | CC | 0.010599 |
| GO:0005874 | microtubule | CC | 0.002197 |
| GO:0005515 | protein binding | MF | 0.035746 |
| GO:0003824 | catalytic activity | MF | 0.039472 |
| GO:0003676 | nucleic acid binding | MF | 0.035847 |
| GO:0003723 | RNA binding | MF | 0.005421 |
| GO:0008270 | zinc ion binding | MF | 0.013125 |
| GO:0046914 | transition metal ion binding | MF | 0.028036 |
| GO:0140096 | catalytic activity, acting on a protein | MF | 0.008336 |
| GO:0004175 | endopeptidase activity | MF | 0.00233 |
| GO:0008233 | peptidase activity | MF | 0.006311 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muller, P.R.; Lee, T.J.; Zhi, W.; Kumar, S.; Vyavahare, S.; Sharma, A.; Kumar, V.; Isales, C.M.; Hunter, M.; Fulzele, S. Proteomic Analysis of Female Synovial Fluid to Identify Novel Biomarkers for Osteoarthritis. Life 2023, 13, 605. https://doi.org/10.3390/life13030605
Muller PR, Lee TJ, Zhi W, Kumar S, Vyavahare S, Sharma A, Kumar V, Isales CM, Hunter M, Fulzele S. Proteomic Analysis of Female Synovial Fluid to Identify Novel Biomarkers for Osteoarthritis. Life. 2023; 13(3):605. https://doi.org/10.3390/life13030605
Chicago/Turabian StyleMuller, P. Robinson, Tae Jin Lee, Wenbo Zhi, Sandeep Kumar, Sagar Vyavahare, Ashok Sharma, Vikas Kumar, Carlos M. Isales, Monte Hunter, and Sadanand Fulzele. 2023. "Proteomic Analysis of Female Synovial Fluid to Identify Novel Biomarkers for Osteoarthritis" Life 13, no. 3: 605. https://doi.org/10.3390/life13030605