
Cytotoxic Effect of Phenylethanoid Glycosides Isolated from Plantago lanceolata L.

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
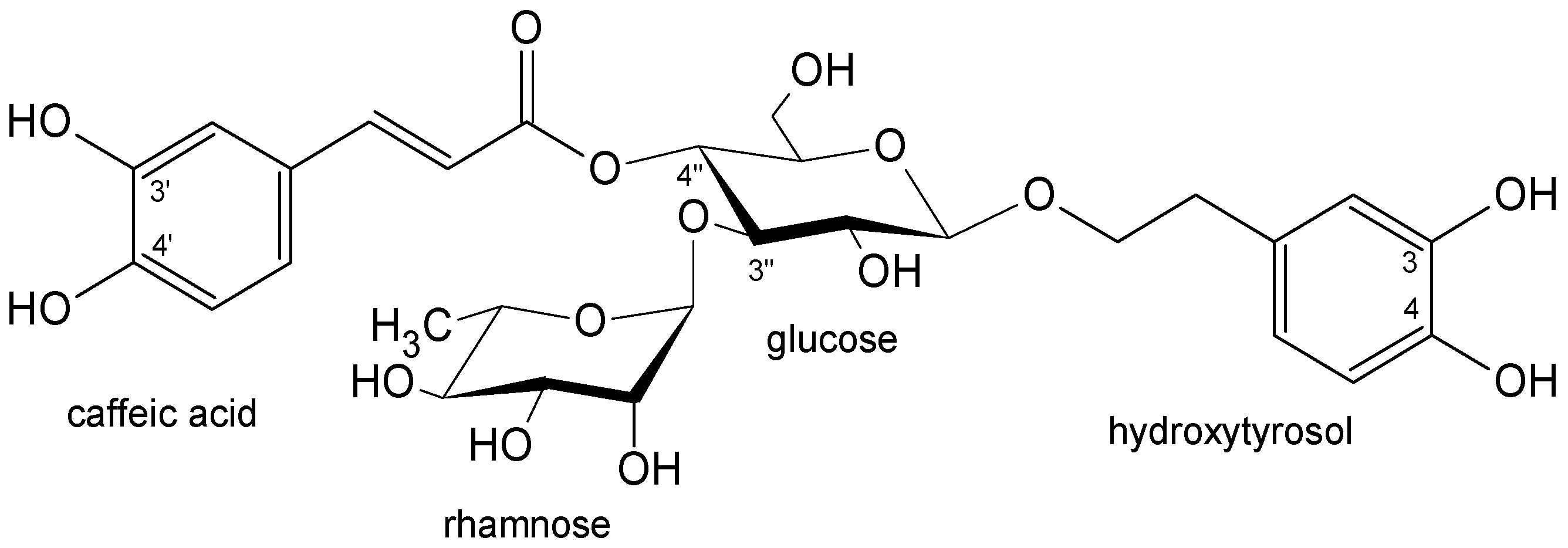
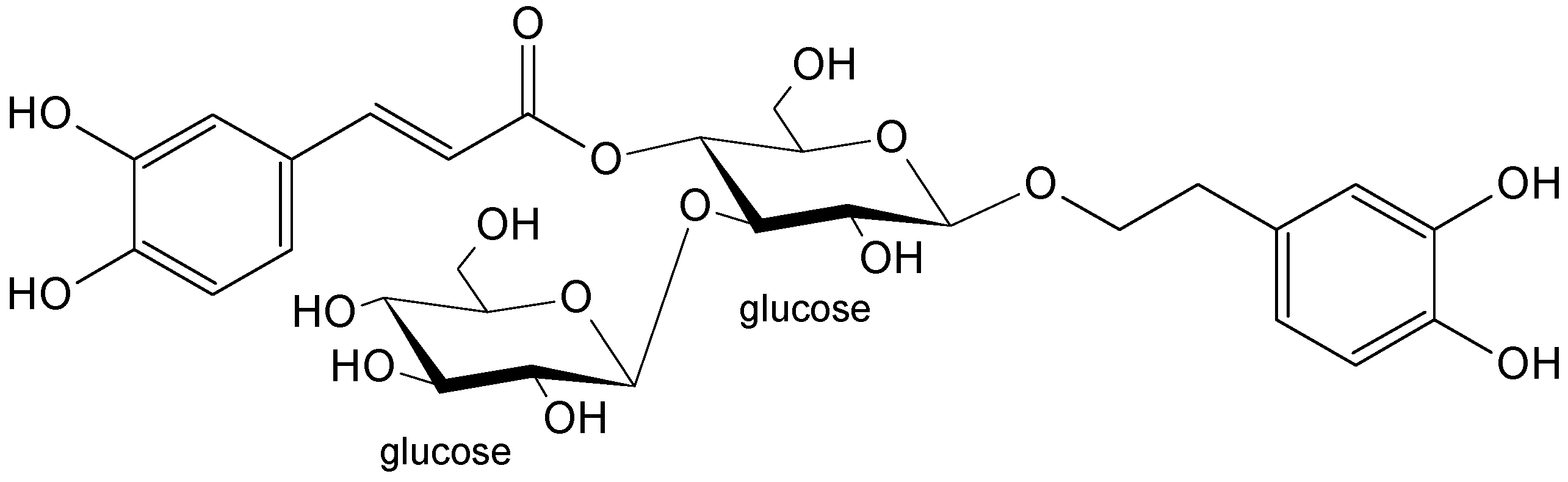
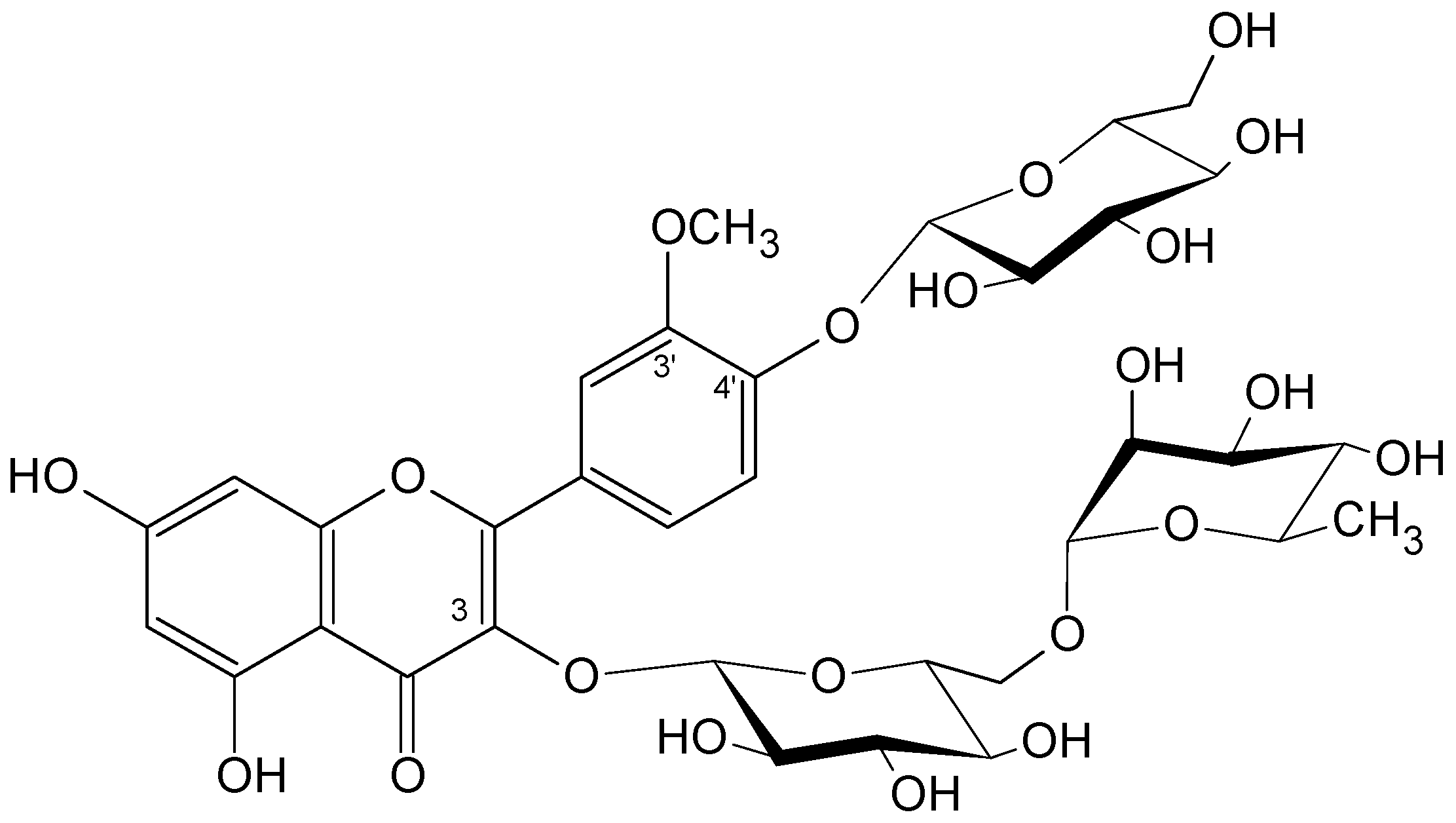
2.1. Plant Material and Isolated Compounds
2.2. Cell Culture
2.3. Cell Proliferation Assay
2.4. Statistical Analysis
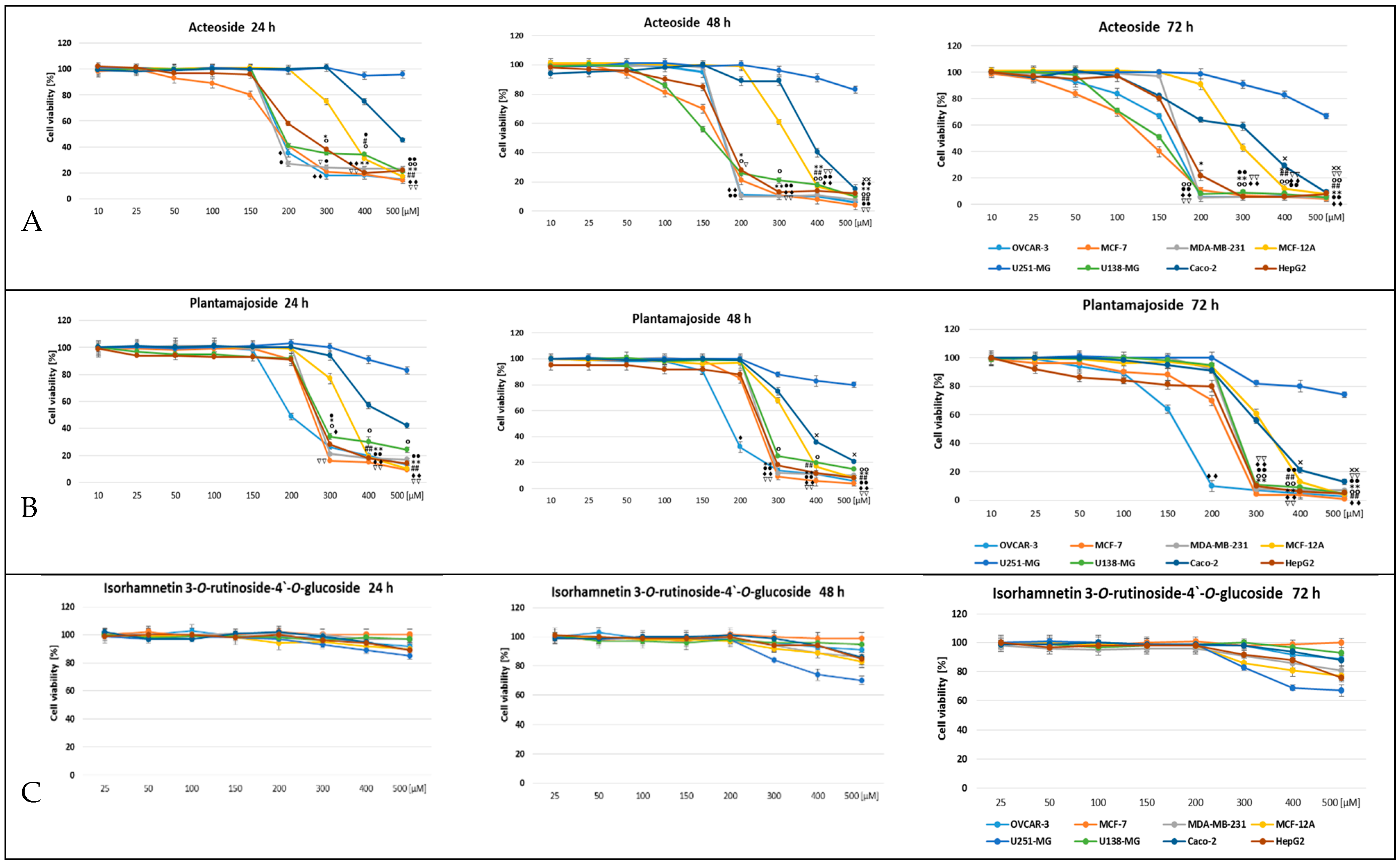
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pol, M.; Schmidtke, K.; Lewandowska, S. Plantago lanceolata—An overview of its agronomically and healing valuable features. Open Agric. 2021, 6, 479–488. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet; 2023. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:321285-2 (accessed on 30 January 2023).
- Mayer, J.G. Plantain (Plantago lanceolata)—Medicinal plant of the year 2014. Z. Phytother. 2013, 34, 242–243. [Google Scholar] [CrossRef]
- Gonçalves, S.; Romano, A. The medicinal potential of plants from the genus Plantago (Plantaginaceae). Ind. Crops Prod. 2016, 83, 213–226. [Google Scholar] [CrossRef]
- Alsaraf, K.M.; Mohammad, M.H.; Al-Shammari, A.M.; Abbas, I.S. Selective cytotoxic effect of Plantago lanceolata L. against breast cancer cells. J. Egypt Nat. Cancer Inst. 2019, 31, 10. [Google Scholar] [CrossRef] [Green Version]
- Bahadori, M.B.; Sarikurkcu, C.; Kocak, M.S.; Calapoglu, M.; Uren, M.C.; Ceylan, O. Plantago lanceolata as a source of health-beneficial phytochemicals: Phenolic profile and antioxidant capacity. Food Biosci. 2020, 34, 1000536. [Google Scholar] [CrossRef]
- Majkić, T.; Bekvalac, K.; Beara, I. Plantain (Plantago L.) species as modulators of prostaglandin E2 and thromboxane A2 production in inflammation. J. Ethnopharmacol. 2020, 262, 113140. [Google Scholar] [CrossRef]
- Gálvez, M.; Martin-Cordero, C.; Lopez-Lazaro, M.; Cortes, F.; Ayuso, M.J. Cytotoxic effect of Plantago spp. on cancer cell lines. J. Ethnopharmacol. 2003, 88, 125–130. [Google Scholar] [CrossRef]
- Rønsted, N.; Göbel, E.; Franzyk, H.; Jensen, S.R.; Olsen, C.E. Chemotaxonomy of Plantago. Iridoid glucosides and caffeoyl phenylethanoid glycosides. Phytochemistry 2000, 55, 337–348. [Google Scholar] [CrossRef]
- Rønsted, N.; Franzyk, H.; Mølgaard, P.; Jaroszewski, J.W.; Jensen, S.R. Chemotaxonomy and evolution of Plantago L. Plant Syst. Evol. 2003, 242, 63–82. [Google Scholar] [CrossRef]
- Murai, M.; Tamayama, Y.; Nishibe, S. Phenylethanoids in the herb of Plantago lanceolata and inhibitory effects on arachidonic acid-induced mouse ear edema. Planta Med. 1995, 61, 479–480. [Google Scholar] [CrossRef]
- Zeng, X.; Guo, F.; Ouyang, D. A review of the pharmacology and toxicology of aucubin. Fitoterapia 2020, 140, 104443. [Google Scholar] [CrossRef] [PubMed]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Ravn, H.W.; Mondolot, L.; Kelly, M.T.; Lykke, A.M. Plantamajoside—A current review. Phytochem. Lett. 2015, 12, 42–53. [Google Scholar] [CrossRef]
- Kawashty, S.A.; Gamal-el-Din, E.; Abdalla, M.F.; Saleh, N.A.M. Flavonoids of Plantago species in Egypt. Biochem. Syst. Ecol. 1994, 22, 729–733. [Google Scholar] [CrossRef]
- Fleer, H.; Verspohl, E.J. Antispasmodic activity of an extract from Plantago lanceolata L. and some isolated compounds. Phytomedicine 2007, 14, 409–415. [Google Scholar] [CrossRef]
- Beara, I.N.; Lesjak, M.M.; Orčić, D.Z.; Simin, N.Đ.; Četojević-Simin, D.D.; Božin, B.N.; Mimica-Dukić, N.M. Comparative analysis of phenolic profile, antioxidant, anti-inflammatory and cytotoxic activity of two closely-related Plantain species: Plantago altissima L. and Plantago lanceolata L. LWT-Food Sci. Technol. 2012, 47, 64–70. [Google Scholar] [CrossRef]
- Janković, T.; Zdunić, G.; Beara, I.; Balog, K.; Pljevljakušić, D.; Stešević, D.; Šavikin, K. Comparative study of some polyphenols in Plantago species. Biochem. Syst. Ecol. 2012, 42, 69–74. [Google Scholar] [CrossRef]
- European Pharmacopoeia. Ribwort leaf. Plantaginis Lanceolatae Folium, 10th ed.; Council of Europe: Strasbourg, France, 2019; p. 1599. [Google Scholar]
- Urząd Rejestracji Produktów Leczniczych, Wyrobów Medycznych i Produktów Biobójczych [Office for Registration of Medicinal Products, Medical Devices and Biocidal Products]. Plantaginis Lanceolatae Folium. In Polish Pharmacopoeia XI; Urząd Rejestracji Produktów Leczniczych, Wyrobów Medycznych i Produktów Biobójczych [Office for Registration of Medicinal Products, Medical Devices and Biocidal Products]: Warsaw, Poland, 2017; p. 1725. [Google Scholar]
- ESCOP (European Scientific Cooperation On Phytotherapy). Plantaginis lanceolatae folium/herba. In ESCOP Monographs: The Scientific Foundation for Herbal Medicinal Products, 2nd ed.; Thieme: Stuttgart, Germany, 2003; pp. 383–387. [Google Scholar]
- Schlauer, J.; Budzianowski, J.; Kukulczanka, K.; Ratajczak, L. Acteoside and related phenylethanoid glycosides in Byblis liniflora Salisb. plants propagated in vitro and its systematic significance. Acta Soc. Bot. Pol. 2004, 73, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.A.; Hossain, R.; Roy, P.; Jain, D.; Mohammad Saikat, A.S.; Roy Shuvo, A.P.; Akram, M.; Elbossaty, W.F.; Khan, I.N.; Painuli, S.; et al. Anticancer effects of acteoside: Mechanistic insights and therapeutic status. Eur. J. Pharmacol. 2022, 916, 174699. [Google Scholar] [CrossRef]
- Xiao, Y.; Ren, Q.; Wu, L. The pharmacokinetic property and pharmacological activity of acteoside: A review. Biomed. Pharmacother. 2022, 153, 113296. [Google Scholar] [CrossRef]
- Goyal, A.; Kushwah, P.S.; Agrawal, N. Therapeutic potential of plantamajoside. Rev. Bras. Farmacogn. 2022, 32, 355–364. [Google Scholar] [CrossRef]
- Budzianowska, A.; Budzianowski, J. A new flavonoid, a new phenylethanoid glycoside and related compounds isolated from the inflorescences of Plantago lanceolata L. Nat. Prod. Res. 2022, 36, 3813–3824. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Loizzo, M.R.; Bonesi, M.; Menichini, F.; Statti, G.A.; Menichini, F.Z. In vitro cytotoxic activity of Salsola oppositifolia Desf. (Amaranthaceae) in a panel of tumour cell lines. Z. Nat. C 2008, 63, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Available online: www.who.int (accessed on 29 November 2022).
- World Cancer Research Fund International. Available online: www.wcrf.org (accessed on 29 November 2022).
- Desai, A.G.; Qazi, G.N.; Ganju, R.K.; El-Tamer, M.; Singh, J.; Saxena, A.K.; Bedi, Y.S.; Taneja, S.C.; Bhat, H.K. Medicinal plants and cancer chemoprevention. Curr. Drug Metab. 2008, 9, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Lichota, A.; Gwozdzinski, K. Anticancer Activity of Natural Compounds from Plant and Marine Environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef] [Green Version]
- Dehelean, C.A.; Marcovici, I.; Soica, C.; Mioc, M.; Coricovac, D.; Iurciuc, S.; Cretu, O.M.; Pinzaru, I. Plant-Derived Anticancer Compounds as New Perspectives in Drug Discovery and Alternative Therapy. Molecules 2021, 26, 1109. [Google Scholar] [CrossRef]
- Ram, V.J.; Kumari, S. Natural products of plant origin as anticancer agents. Drug News Perspect. 2001, 14, 465–482. [Google Scholar] [CrossRef]
- Chou, T.C.; Martin, N. CompuSyn Software. In CompuSyn for Drug Combinations: PC Software and User’s Guide: A Computer Program for Quantitation of Synergism and Antagonism in Drug Combinations, and the Determination of IC50 and ED50 and LD50 Values; ComboSyn Inc.: Paramus, NJ, USA, 2005; Available online: https://www.combosyn.com (accessed on 27 September 2022).
- Pettit, G.R.; Numata, A.; Takemura, T.; Ode, R.H.; Narula, A.S.; Schmidt, J.M.; Cragg, G.M.; Pase, C.P. Antineoplastic agents, 107. Isolation of acteoside and isoacteoside from Castilleja linariaefolia. J. Nat. Prod. 1990, 53, 456–458. [Google Scholar] [CrossRef]
- Saracoglu, I.; Inoue, M.; Calis, I.; Ogihara, Y. Studies on constituents with cytotoxic and cytostatic activity of two Turkish medicinal plants Phlomis armeniaca and Scutellaria salviifolia. Biol. Pharm. Bull. 1995, 18, 1396–1400. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.; Yun, Z.; Hong, Z.; Baoning, S.; Rong-liang, Z. Differentiation of human gastric adenocarcinoma cell line MGc80-3 induced by verbascoside. Planta Med. 1997, 63, 499–502. [Google Scholar] [CrossRef]
- Inoue, M.; Sakuma, Z.; Ogihara, Y.; Saracoglu, I. Induction of apoptotic cell death in HL-60 cells by acteoside, a phenylpropanoid glycoside. Biol. Pharm. Bull. 1998, 21, 81–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cometa, F.; Tomassini, L.; Nicoletti, M. Phenylpropanoid glycosides. Distribution and pharmacological activity. Fitoterapia 1993, 64, 195–217. [Google Scholar]
- Şenol, H.; Tulay, P.; Ergören, M.Ç.; Hanoğlu, A.; Çalış, İ.; Mocan, G. Cytotoxic Effects of Verbascoside on MCF-7 and MDA-MB-231. Turk. J. Pharm. Sci. 2021, 18, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Cheimonidi, C.; Samara, P.; Polychronopoulos, P.; Tsakiri, E.N.; Nikou, T.; Myrianthopoulos, V.; Sakellaropoulos, T.; Zoumpourlis, V.; Mikros, E.; Papassideri, I.; et al. Selective cytotoxicity of the herbal substance acteoside against tumor cells and its mechanistic insights. Redox Biol. 2018, 16, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.M.; El-Kersh, D.M.; Wagdy, H.A.; Elmazar, M.M. Verbascoside: Identification, quantification, and potential sensitization of colorectal Cancer cells to 5-FU by targeting PI3K/AKT pathway. Sci. Rep. 2018, 8, 16939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, F.; Nagoa, T.; Okabe, H. Antiproliferative constituents in plants 9. Aerial parts of Lippia dulcis and Lippia canescens. Biol. Pharm. Bull. 2002, 25, 920–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagoa, T.; Abe, F.; Okabe, H. Antiproliferative constituents in the plants 7. Leaves of Clerodendron bungei and leaves and bark of C. trichotomum. Biol. Pharm. Bull. 2001, 24, 1338–1341. [Google Scholar] [CrossRef] [Green Version]
- Emma, M.R.; Augello, G.; Di Stefano, V.; Azzolina, A.; Giannitrapani, L.; Montalto, G.; Cervello, M.; Cusimano, A. Potential Uses of Olive Oil Secoiridoids for the Prevention and Treatment of Cancer: A Narrative Review of Preclinical Studies. Int. J. Mol. Sci. 2021, 22, 1234. [Google Scholar] [CrossRef]
- Pei, S.; Yang, X.; Wang, H.; Zhang, H.; Zhou, B.; Zhang, D.; Lin, D. Plantamajoside, a potential antitumor herbal medicine inhibits breast cancer growth and pulmonary metastasis by decreasing the activity of matrix metalloproteinase-9 and -2. BMC Cancer 2015, 15, 965–976. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Jiang, X.; Yin, G.; Liu, Y.; Liu, Z.; Meng, L.; Wu, J.; Wu, H. The herbal agent plantamajoside, exerts a potential inhibitory effect on the development of hepatocellular carcinoma. Exp. Ther. Med. 2021, 21, 573–583. [Google Scholar] [CrossRef]
- Wang, Z.; Zuo, J.; Zhang, L.; Zhang, Z.; Wei, Y. Plantamajoside promotes metformin-induced apoptosis, autophagy and proliferation arrest of liver cancer cells via suppressing Akt/GSK3β signaling. Hum. Exp. Toxicol. 2022, 41, 9603271221078868. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Xu, J.; Li, C.; Dai, X.; Wu, T.; Wen, J. Plantamajoside inhibits the proliferation and epithelial-to-mesenchymal transition in hepatocellular carcinoma cells via modulating hypoxia-inducible factor-1α-dependent gene expression. Cell Biol. Int. 2020, 44, 1616–1627. [Google Scholar] [CrossRef] [PubMed]
- Zan, Y.; Dai, Z.; Liang, L.; Deng, Y.; Dong, L. Co-delivery of plantamajoside and sorafenib by a multi-functional nanoparticle to combat the drug resistance of hepatocellular carcinoma through reprograming the tumor hypoxic microenvironment. Drug Deliv. 2019, 26, 1080–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, X.; Li, L.; Sun, L. Plantamajoside inhibits hypoxia-induced migration and invasion of human cervical cancer cells through the NF-κB and PI3K/akt pathways. J. Recept. Signal Transduct. 2021, 41, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, D.; Li, M.; Gao, X.; Shi, G.; Zhao, H. Plantamajoside inhibits lipopolysaccharideinduced epithelial-mesenchymal transition through suppressing the NF-κB/IL-6 signaling in esophageal squamous cell carcinoma cells. Biomed. Pharmacother. 2018, 102, 1045–1051. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, M.; Chen, S.; Wu, Q. Plantamajoside represses the growth and metastasis of malignant melanoma. Exp. Ther. Med. 2020, 19, 2296–2302. [Google Scholar] [CrossRef] [Green Version]
- Piwowarczyk, L.; Mlynarczyk, D.T.; Krajka-Kuźniak, V.; Majchrzak-Celińska, A.; Budzianowska, A.; Tomczak, S.; Budzianowski, J.; Woźniak-Braszak, A.; Pietrzyk, R.; Baranowski, M.; et al. Natural compounds in liposomal nanoformulations of potential clinical application in glioblastoma. Cancers 2022, 14, 6222. [Google Scholar] [CrossRef]
- Hwang, T.W.; Kim, D.H.; Kim, D.B.; Jang, T.W.; Kim, G.H.; Moon, M.; Yoon, K.A.; Choi, D.E.; Park, J.H.; Kim, J.J. Synergistic anticancer effect of acteoside and temozolomide-based glioblastoma chemotherapy. Int. J. Mol. Med. 2019, 43, 1478–1486. [Google Scholar] [CrossRef]
- Antunes-Ricardo, M.; Moreno-García, B.E.; Gutiérrez-Uribe, J.A.; Aráiz-Hernández, D.; Alvarez, M.M.; Serna-Saldivar, S.O. Induction of apoptosis in colon cancer cells treated with isorhamnetin glycosides from Opuntia ficus-indica pads. Plant Foods Hum. Nutr. 2014, 69, 331–336. [Google Scholar] [CrossRef]
- Liao, Y.-F.; Rao, Y.K.; Tzeng, Y.-M. Aqueous extract of Anisomeles indica and its purified compound exerts anti-metastatic activity through inhibition of NF-κB/AP-1-dependent MMP-9 activation in human breast cancer MCF-7 cells. Food Chem. Toxicol. 2012, 50, 2930–2936. [Google Scholar] [CrossRef]
- Guangmiao, F.; Haihong, P.; Wong, Y.H. Naturally Occurring Phenylethanoid Glycosides: Potential Leads for New Therapeutics. Curr. Med. Chem. 2008, 15, 2592–2613. [Google Scholar]
- Wu, L.; Georgiev, M.I.; Cao, H.; Nahar, L.; El-Seedi, H.R.; Sarker, S.D.; Xiao, J.; Lu, B. Therapeutic potential of phenylethanoid glycosides: A systematic review. Med. Res. Rev. 2020, 40, 2605–2649. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Leal, C.A.; Puente-Rarza, C.A.; Garcia-Lara, S. In vitro plant tissue culture: Means for production of biological active compounds. Planta 2008, 248, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fons, F.; Gargadennec, A.; Rapior, S. Culture of Plantago species as bioactive components resources: A 20-year review and recent applications. Acta Biol. Gall. 2008, 155, 277–300. [Google Scholar] [CrossRef]
- Budzianowska, A.; Skrzypczak, L.; Budzianowski, J. Phenylethanoid glucosides from in vitro propagated plants and callus cultures of Plantago lanceolata. Planta Med. 2004, 70, 834–840. [Google Scholar] [CrossRef]
- Budzianowska, A.; Kikowska, M.; Małkiewicz, M.; Karolak, I.; Budzianowski, J. Phenylethanoid glycosides in Plantago media L. organs obtained in in vitro cultures. Acta Biol. Crac. Ser. Bot. 2019, 61, 75–86. [Google Scholar]
- Budzianowska, A.; Kikowska, M.; Budzianowski, J. Adventitious root culture of Plantago ovata Forssk. As a source of phenylethanoid glycosides. Ind. Crops Prod. 2022, 180, 114773. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 [μM] | ||||||
|---|---|---|---|---|---|---|
| Acteoside | Plantamajoside | |||||
| Cell Line | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h |
| OVCAR-3 | 314.1 ± 3.52 d,F | 232.0 ± 5.08 d,D | 162.8 ± 3.41 b,B | 243.5 ± 2.61 a,E | 201.3 ± 5.29 a,C | 138.9 ± 4.85 a,A |
| MCF-7 | 219.1 ± 1.40 a,D | 154.2 ± 2.71 a,B | 113.1 ± 2.81 a,A | 312.9 ± 4.11 c,E | 243.8 ± 3.57 c,D | 170.8 ± 2.50 c,C |
| MDA-MB-231 | 312.2 ± 3.28 d,D | 244.9 ± 4.96 e,B | 200.2 ± 2.45 d,A | 366.7 ± 3.71 d,e,E | 311.7 ± 4.83 d,D | 263.1 ± 3.20 d,C |
| MCF-12A | 362.3 ± 1.86 e,D | 346.8 ± 3.67 f,C | 326.3 ± 3.89 f,B | 376.3 ± 7.91 e,E | 339.2 ± 4.31 e,C | 296.2 ± 4.90 e,A |
| U251-MG | 3152.7 ± 4.15 f,F | 2412.5 ± 7.97 h,E | 1165.3 ± 6.05 g,C | 2099.9 ± 5.61 g,D | 1117.3 ± 6.21 g,B | 985.6 ± 5.71 g,A |
| U138-MG | 274.3 ± 2.61 c,D | 201.9 ± 4.90 b,B | 156.6 ± 4.74 b,A | 359.5 ± 2.75 d,F | 342.1 ± 2.35 e,E | 266.7 ± 3.88 d,C |
| Caco-2 | 507.6 ± 4.05 e,F | 466.9 ± 8.71 g,E | 280.3 ± 6.13 e,A | 447.0 ± 6.16 f,D | 378.0 ± 5.23 f,C | 316.4 ± 4.27 f,B |
| HepG2 | 261.3 ± 3.52 b,D | 219.6 ± 2.65 c,C | 173.8 ± 1.59 c,B | 284.7 ± 5.04 b,E | 228.2 ± 1.95 b,C | 156.1 ± 2.90 b,A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budzianowska, A.; Totoń, E.; Romaniuk-Drapała, A.; Kikowska, M.; Budzianowski, J. Cytotoxic Effect of Phenylethanoid Glycosides Isolated from Plantago lanceolata L. Life 2023, 13, 556. https://doi.org/10.3390/life13020556
Budzianowska A, Totoń E, Romaniuk-Drapała A, Kikowska M, Budzianowski J. Cytotoxic Effect of Phenylethanoid Glycosides Isolated from Plantago lanceolata L. Life. 2023; 13(2):556. https://doi.org/10.3390/life13020556
Chicago/Turabian StyleBudzianowska, Anna, Ewa Totoń, Aleksandra Romaniuk-Drapała, Małgorzata Kikowska, and Jaromir Budzianowski. 2023. "Cytotoxic Effect of Phenylethanoid Glycosides Isolated from Plantago lanceolata L." Life 13, no. 2: 556. https://doi.org/10.3390/life13020556