

Allophylus africanus Stem Bark Extract Modulates the Mitochondrial Apoptotic Pathway in Human Stomach Cancer Cells
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Chemicals, Reagents and Equipment
2.2. Plant Material and Extraction
2.3. Cell-Based Assays
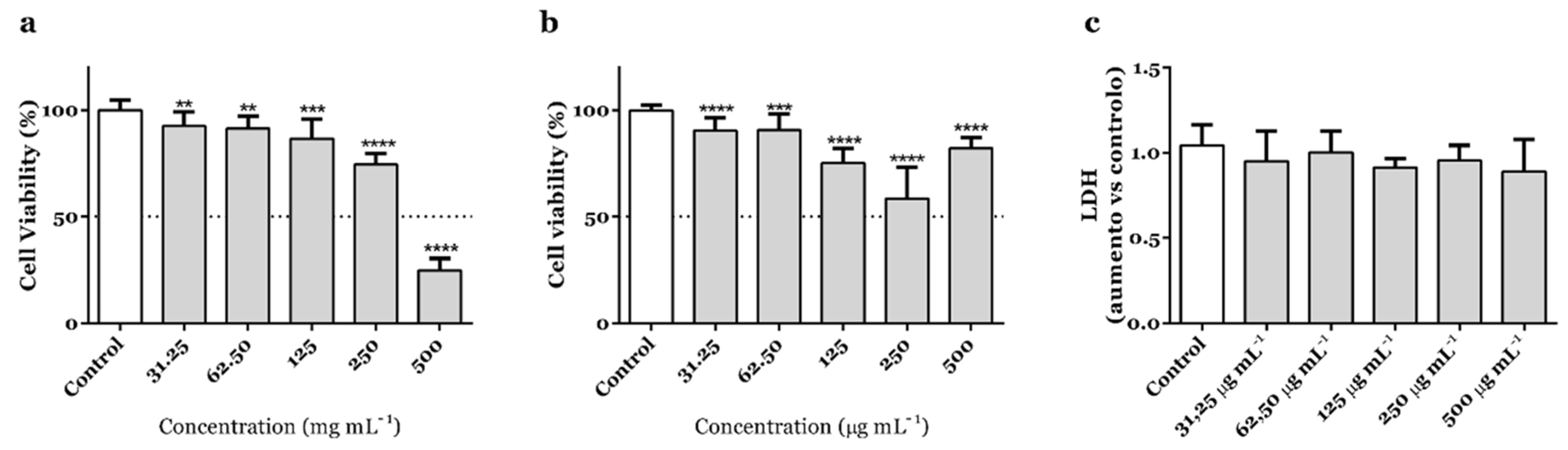
2.4. Cell Viability and Membrane Integrity Assays
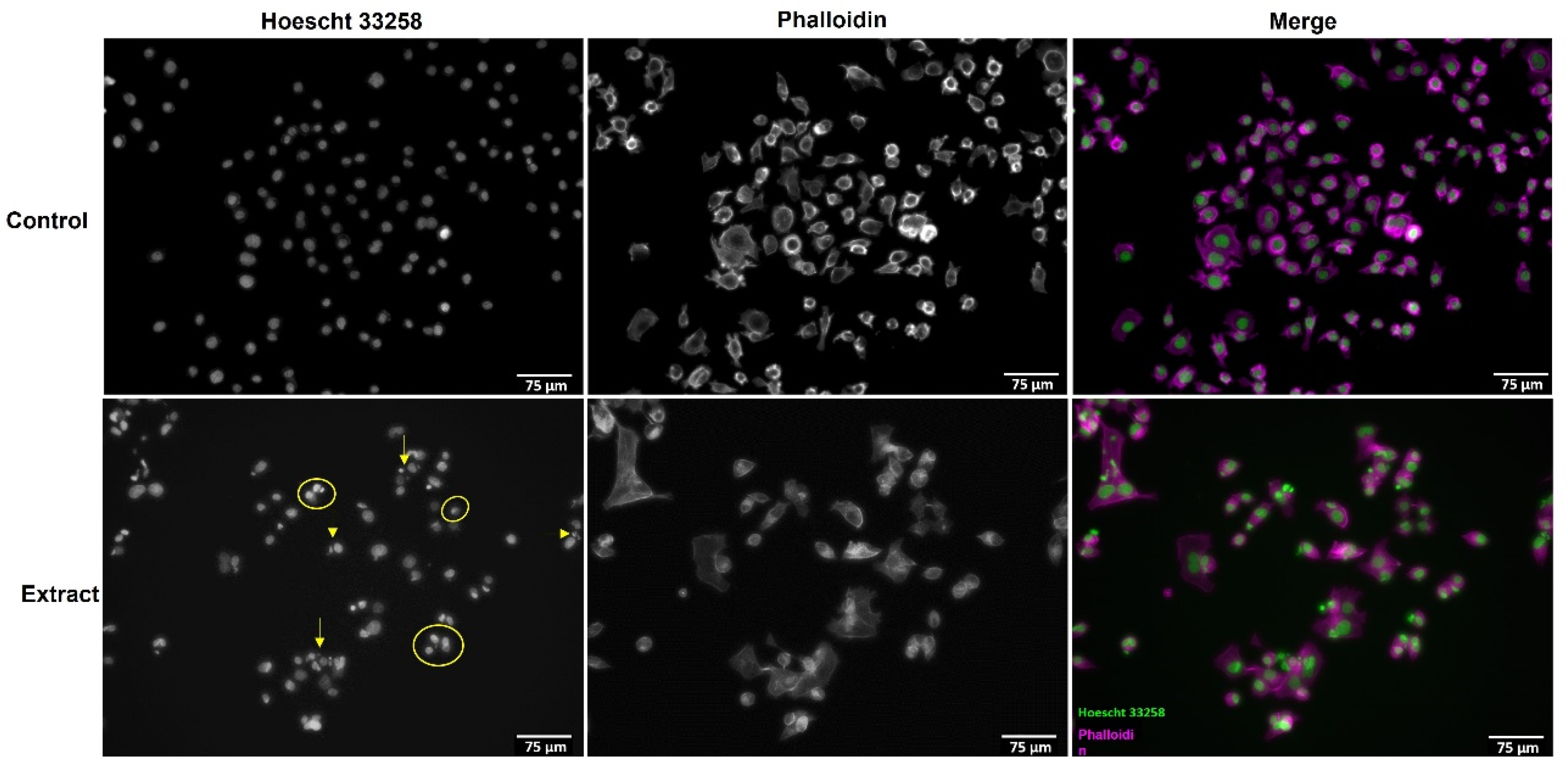
2.5. Morphological Image Analysis
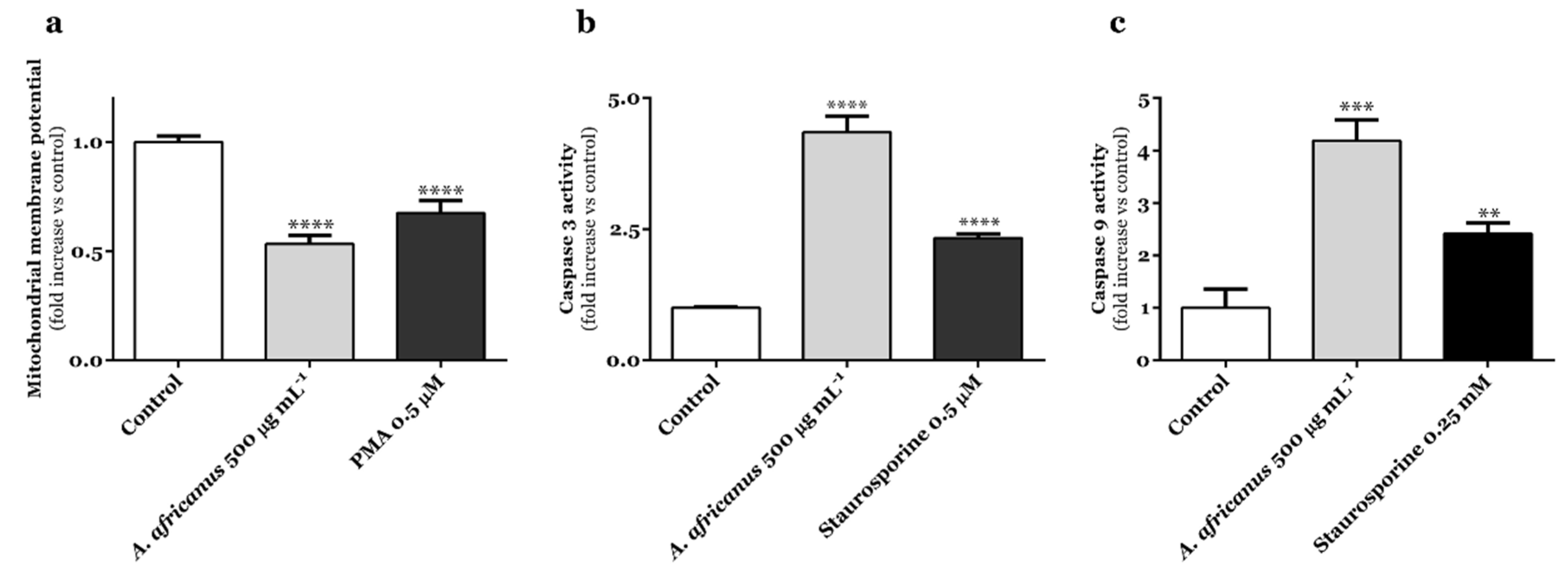
2.6. Caspase-3 and -9 Activity Assay
2.7. Mitochondrial Membrane Potential Assay
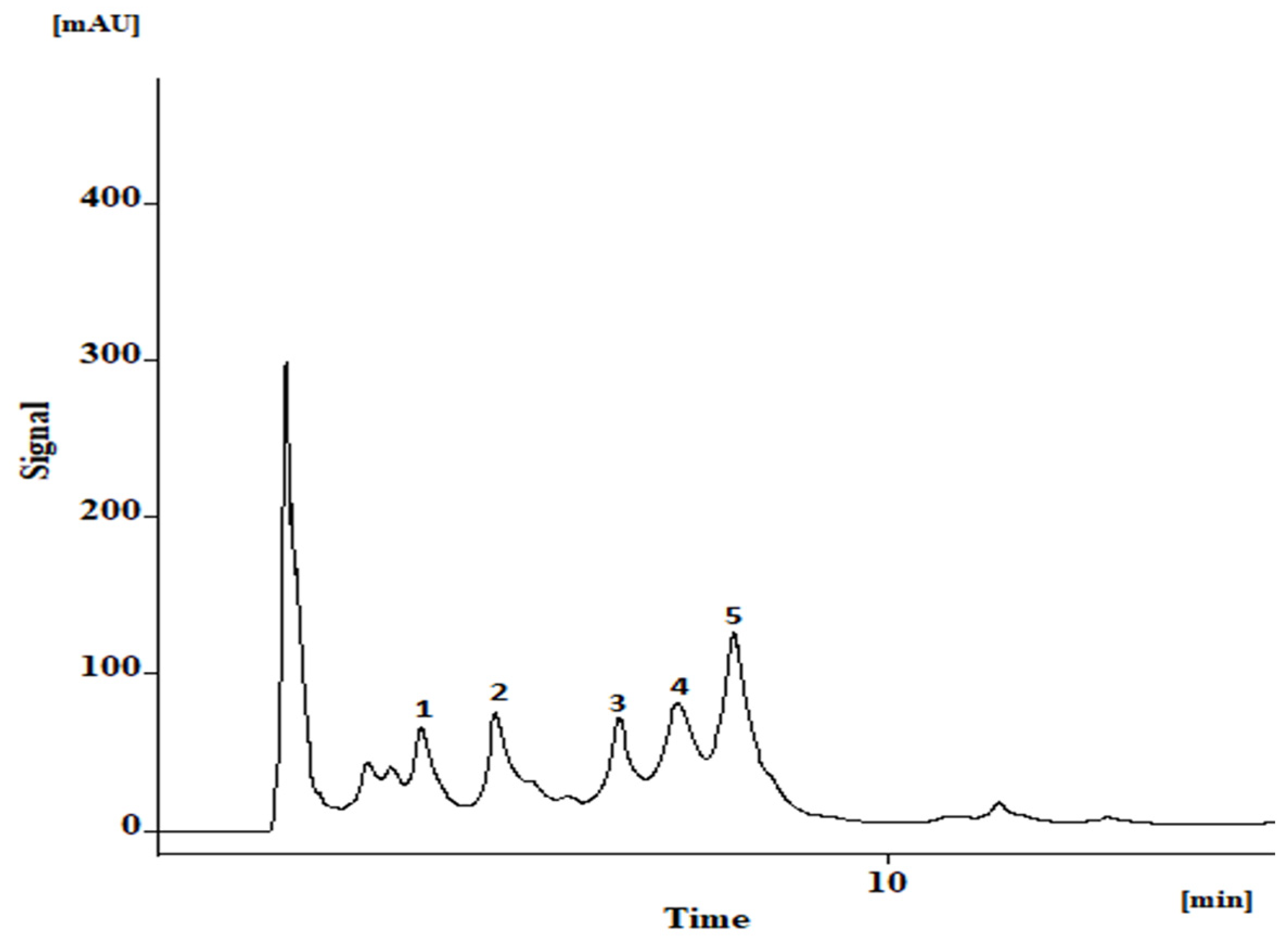
2.8. Phenolic Compounds Analysis by HPLC-DAD
2.9. Statistical Analysis
3. Results
3.1. A. africanus Stem Bark Shows Selective Cytotoxicity in Human Gastric Adenocarcinoma AGS Cells
3.2. A. africanus Stem Bark Causes Apoptotic Cell Death in AGS Cells
3.3. A. africanus Stem Bark Causes Caspase-Dependent Cell Death in AGS Cells via Activation of the Intrinsic Apoptosis Pathway
3.4. Bioactive Phenolics in A. africanus Stem Bark Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Apaf-1 | Apoptotic protease activating factor-1 |
| CHAPS | 3-[(3-cholamidopropyl)dimethylammonium]-1-propanesulfonate |
| DAD | Diode array detector |
| DMSO | Dimethyl sulfoxide |
| DTT | 1,4-dithiothreitol |
| EDTA | Ethylenediaminetetraacetic acid |
| FBS | Fetal bovine serum |
| HEPES | 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid |
| HPLC | High Performance Liquid Chromatography |
| IC50 | Inhibitory concentration of 50% |
| LDH | Lactate dehydrogenase |
| MMP | Mitochondrial membrane potential |
| MTT | 3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide |
| NCI-H23 | Human non-small cell lung cancer |
| OSCC | Oral squamous cell carcinoma |
| PARP | Polymerase Poly(ADP-Ribose) |
| PMA | Phorbol 12-myristate 13-acetate |
| ROS | Reactive oxygen species |
| Rt | Retention time |
References
- Chavan, R.; Gaikwad, D. The Ethnobotany, Phytochemistry and Biological Properties of Allophylus Species Used in Traditional Medicine: A Review. World J. Pharm. Pharm. Sci. 2016, 5, 664–682. [Google Scholar] [CrossRef]
- Trevizan, L.N.F.; Nascimento, K.F.D.; Santos, J.A.; Kassuya, C.A.L.; Cardoso, C.A.L.; Vieira, M.D.C.; Moreira, F.M.F.; Croda, J.; Formagio, A.S.N. Anti-Inflammatory, Antioxidant and Anti-Mycobacterium Tuberculosis Activity of Viridiflorol: The Major Constituent of Allophylus edulis (A. St.-Hil., A. Juss. & Cambess.) Radlk. J. Ethnopharmacol. 2016, 192, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.; González, A.; Castro-Gamboa, I.; Gonzalez, D.; Rossini, C. First Record of L-Quebrachitol in Allophylus edulis (Sapindaceae). Carbohydr. Res. 2008, 343, 2699–2700. [Google Scholar] [CrossRef] [PubMed]
- Arisawa, M.; Morinaga, Y.; Nishi, Y.; Ueno, H.; Suzuki, S.; Hayashi, T.; Shimizu, M.; Yoshizaki, M.; Morita, N.; Berganza, L.H. Chemical and Pharmaceutical Studies on Medicinal Plants in Paraguayi) Allophylus Edutis RADLK. Jpn. Soc. Pharmacogn. 1988, 43, 78–80. [Google Scholar]
- Yajía, M.E.; Martí, D.A.; Bidau, C.J.; Amat, A.G.; Riglos, A.G.; Silvestroni, A. Genotoxicity Evaluation of Allophylus edulis (Camb.) Radlk. (Sapindaceae) Aqueous Extract. Acta Hortic. 1999, 501, 31–35. [Google Scholar] [CrossRef]
- Hoffmann-Bohm, K.; Lotter, H.; Seligmann, O.; Wagner, H. Antihepatotoxic C-Glycosylflavones from the Leaves of Allophyllus edulis Var. edulis and gracilis. Planta Med. 1992, 58, 544–548. [Google Scholar] [CrossRef]
- Leenhouts, P.W. A Conspectus of the Genus Allophylus (Sapindaceae). The Problem of the Complex Species. Blumea Biodivers. Evol. Biogeogr. Plants 1967, 15, 301–358. [Google Scholar]
- Adeyemi, T.O.; Ogundipe, O.T.; Olowokudejo, J.D. A Review of the Taxonomy of African Sapindaceae Based on Quantitative and Qualitative Characters. Ife J. Sci. 2016, 15, 303–314. [Google Scholar] [CrossRef]
- Oladosu, I.A.; Balogun, S.O.; Ademowo, G.O. Phytochemical Screening, Antimalarial and Histopathological Studies of Allophylus africanus and Tragia benthamii. Chin. J. Nat. Med. 2013, 11, 371–376. [Google Scholar] [CrossRef]
- Ibrahim, F.S.; Mohammed, Z.; Nuhu, A.; Shehu, S.; Ilyas, N. Acute Toxicity and Anti-Inflammatory Activity of Hydromethanol Extract of Allophylus africanus Beauv in rats. J. Herbmed Pharmacol. 2018, 7, 119–123. [Google Scholar] [CrossRef]
- Lemordant, D. Contribution à l’ethnobotanique Éthiopienne (Fin). J. D’agriculture Tradit. Et De Bot. Appliquée 1971, 18, 142–179. [Google Scholar] [CrossRef]
- Oladosu, I.A.; Balogun, S.O.; Liu, Z.Q. Chemical Constituents of Allophylus africanus. Chin. J. Nat. Med. 2015, 13, 133–141. [Google Scholar] [CrossRef]
- Gathirwa, J.W.; Rukunga, G.M.; Mwitari, P.G.; Mwikwabe, N.M.; Kimani, C.W.; Muthaura, C.N.; Kiboi, D.M.; Nyangacha, R.M.; Omar, S.A. Traditional Herbal Antimalarial Therapy in Kilifi District, Kenya. J. Ethnopharmacol. 2011, 134, 434–442. [Google Scholar] [CrossRef]
- Chifundera, K. Contribution to the Inventory of Medicinal Plants from the Bushi Area, South Kivu Province, Democratic Republic of Congo. Fitoterapia 2001, 72, 351–368. [Google Scholar] [CrossRef]
- Catarino, L.; Havik, P.J.; Romeiras, M.M. Medicinal Plants of Guinea-Bissau: Therapeutic Applications, Ethnic Diversity and Knowledge Transfer. J. Ethnopharmacol. 2016, 183, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Balogun, O.S.; Oladosu, I.A.; Liu, Z. Fatty Acid Profile and in Vitro Biological Activities of Allophylus africanus (P. Beauv). J. Herbs Spices Med. Plants 2016, 22, 238–246. [Google Scholar] [CrossRef]
- Balogun, S.O.; Oladosu, I.A.; Liu, Z. Chemical Compositions and Antioxidant Potential of Essential Oils from Leaves and Flowers of Allophylus africanus. J. Essent. Oil Bear. Plants 2014, 17, 769–775. [Google Scholar] [CrossRef]
- Ferreres, F.; Gomes, N.G.M.; Valentão, P.; Pereira, D.M.; Gil-Izquierdo, A.; Araújo, L.; Silva, T.C.; Andrade, P.B. Leaves and Stem Bark from Allophylus africanus P. Beauv.: An Approach to Anti-Inflammatory Properties and Characterization of Their Flavonoid Profile. Food Chem. Toxicol. 2018, 118, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Iwu, M.M.; Anyanwu, B.N. Phytotherapeutic Profile of Nigerian Herbs I: Anti-Inflammatory and Anti-Arthritic Agents. J. Ethnopharmacol. 1982, 6, 263–274. [Google Scholar] [CrossRef]
- Biseko, E.Z.; Swai, H.S.; Wachuka Mbugua, R.; Ndung’u, J.W.; Chepng’etich, J.; Gathirwa, J.W. In Vitro Antiproliferative Potential of Annona senegalensis Pers. and Allophylus africanus P Beauv. Plant Extracts against Selected Cancer Cell Lines. J. Med. Plants Res. 2019, 13, 304–311. [Google Scholar] [CrossRef]
- Zeutsop, J.F.; Zébazé, J.N.; Nono, R.N.; Frese, M.; Chouna, J.R.; Lenta, B.N.; Nkeng-Efouet-Alango, P.; Sewald, N. Antioxidant and Cytotoxicity Activities of δ-Tocotrienol from the Seeds of Allophylus africanus. Nat. Prod. Res. 2021, 36, 4661–4671. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.; Ferreres, F.; Macedo, T.; Gil-Izquierdo, Á.; Oliveira, A.P.; Gomes, N.G.M.; Araújo, L.; Pereira, D.M.; Andrade, P.B.; Valentão, P. Activation of Caspase-3 in Gastric Adenocarcinoma AGS Cells by Xylopia aethiopica (Dunal) A. Rich. Fruit and Characterization of Its Phenolic Fingerprint by HPLC-DAD-ESI(Ion Trap)-MS n and UPLC-ESI-QTOF-MS 2. Food Res. Int. 2021, 141, 110121. [Google Scholar] [CrossRef]
- Ribeiro, V.; Ferreres, F.; Oliveira, A.P.; Gomes, N.G.M.; Gil-Izquierdo, Á.; Araújo, L.; Pereira, D.M.; Andrade, P.B.; Valentão, P. HPLC-DAD-ESI/MSn and UHPLC-ESI/QTOF/MSn Characterization of Polyphenols in the Leaves of Neocarya macrophylla (Sabine) Prance Ex F. White and Cytotoxicity to Gastric Carcinoma Cells. Food Res. Int. 2022, 155, 111082. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.; Andrade, P.B.; Valentão, P.; Pereira, D.M. Benzoquinones from Cyperus spp. Trigger IRE1α-Independent and PERK-Dependent ER Stress in Human Stomach Cancer Cells and Are Novel Proteasome Inhibitors. Phytomedicine 2019, 63, 153017. [Google Scholar] [CrossRef] [PubMed]
- Tonnus, W.; Meyer, C.; Paliege, A.; Belavgeni, A.; von Mässenhausen, A.; Bornstein, S.R.; Hugo, C.; Becker, J.U.; Linkermann, A. The Pathological Features of Regulated Necrosis. J. Pathol. 2019, 247, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Toné, S.; Sugimoto, K.; Tanda, K.; Suda, T.; Uehira, K.; Kanouchi, H.; Samejima, K.; Minatogawa, Y.; Earnshaw, W.C. Three Distinct Stages of Apoptotic Nuclear Condensation Revealed by Time-Lapse Imaging, Biochemical and Electron Microscopy Analysis of Cell-Free Apoptosis. Exp. Cell Res. 2007, 313, 3635–3644. [Google Scholar] [CrossRef] [PubMed]
- Hüttemann, M.; Pecina, P.; Rainbolt, M.; Sanderson, T.H.; Kagan, V.E.; Samavati, L.; Doan, J.W.; Lee, I. The Multiple Functions of Cytochrome c and Their Regulation in Life and Death Decisions of the Mammalian Cell: From Respiration to Apoptosis. Mitochondrion 2011, 11, 369–381. [Google Scholar] [CrossRef]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of Cytochrome c Release from Mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef]
- Bradacs, G.; Maes, L.; Heilmann, J. In Vitro Cytotoxic, Antiprotozoal and Antimicrobial Activities of Medicinal Plants from Vanuatu. Phytother. Res. 2010, 24, 800–809. [Google Scholar] [CrossRef]
- Ghagane, S.C.; Puranik, S.I.; Nerli, R.B.; Hiremath, M.B. Evaluation of in Vitro Antioxidant and Anticancer Activity of Allophylus cobbe Leaf Extracts on DU-145 and PC-3 Human Prostate Cancer Cell Lines. Cytotechnology 2017, 69, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Baruah, T.J.; Sharan, R.N.; Kma, L. Vicenin-2: A Potential Radiosensitizer of Non-Small Cell Lung Cancer Cells. Mol. Biol. Rep. 2018, 45, 1219–1225. [Google Scholar] [CrossRef]
- Sahoo, N.; Manchikanti, P.; Dey, S. Herbal Drugs: Standards and Regulation. Fitoterapia 2010, 81, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Albert, A.; Sareedenchai, V.; Heller, W.; Seidlitz, H.K.; Zidorn, C. Temperature Is the Key to Altitudinal Variation of Phenolics in Arnica montana L. Cv. ARBO. Oecologia 2009, 160, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sytar, O.; Zivcak, M.; Bruckova, K.; Brestic, M.; Hemmerich, I.; Rauh, C.; Simko, I. Shift in Accumulation of Flavonoids and Phenolic Acids in Lettuce Attributable to Changes in Ultraviolet Radiation and Temperature. Sci. Hortic. 2018, 239, 193–204. [Google Scholar] [CrossRef]
- Semaan, D.G.; Igoli, J.O.; Young, L.; Marrero, E.; Gray, A.I.; Rowan, E.G. In Vitro Anti-Diabetic Activity of Flavonoids and Pheophytins from Allophylus cominia Sw. on PTP1B, DPPIV, Alpha-Glucosidase and Alpha-Amylase Enzymes. J. Ethnopharmacol. 2017, 203, 39–46. [Google Scholar] [CrossRef]
- Semaan, D.G.; Igoli, J.O.; Young, L.; Gray, A.I.; Rowan, E.G.; Marrero, E. In Vitro Anti-Diabetic Effect of Flavonoids and Pheophytins from Allophylus cominia Sw. on the Glucose Uptake Assays by HepG2, L6, 3T3-L1 and Fat Accumulation in 3T3-L1 Adipocytes. J. Ethnopharmacol. 2018, 216, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Rawat, P.; Dixit, P.; Mishra, D.; Gautam, A.K.; Pandey, R.; Singh, D.; Chattopadhyay, N.; Maurya, R. Anti-Osteoporotic Constituents from Indian Medicinal Plants. Phytomedicine 2010, 17, 993–999. [Google Scholar] [CrossRef]
- David, J.P.; dos Santos, I.D.; David, J.M. A New Sesquiterpene from the Fruits of Allophylus laevigatus. Fitoterapia 2004, 75, 795–798. [Google Scholar] [CrossRef]
- Girish, T.K.; Kumar, K.A.; Prasada Rao, U.J.S. C-Glycosylated Flavonoids from Black Gram Husk: Protection against DNA and Erythrocytes from Oxidative Damage and Their Cytotoxic Effect on HeLa Cells. Toxicol. Rep. 2016, 3, 652–663. [Google Scholar] [CrossRef]
- Hanafi, M.M.M.; Afzan, A.; Yaakob, H.; Aziz, R.; Sarmidi, M.R.; Wolfender, J.L.; Prieto, J.M. In Vitro Pro-Apoptotic and Anti-Migratory Effects of Ficus deltoidea L. Plant Extracts on the Human Prostate Cancer Cell Lines PC3. Front. Pharmacol. 2017, 8, 895. [Google Scholar] [CrossRef] [PubMed]
- Czemplik, M.; Mierziak, J.; Szopa, J.; Kulma, A. Flavonoid C-Glucosides Derived from Flax Straw Extracts Reduce Human Breast Cancer Cell Growth in Vitro and Induce Apoptosis. Front. Pharmacol. 2016, 7, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivis, L.A.; Veloz, R.; María, D.C.L.; Perera, S. Plantas Cubanas Con Efecto Antiinflamatorio Cuban Plants with Anti-Inflammatory Effects. Rev. Cuba. de Farm. 2015, 49, 156–164. [Google Scholar]
- Perveen, S.; El-Shafae, A.M.; Al-Taweel, A.; Fawzy, G.A.; Malik, A.; Afza, N.; Latif, M.; Iqbal, L. Antioxidant and Urease Inhibitory C-Glycosylflavonoids from Celtis Africana. J. Asian Nat. Prod. Res. 2011, 13, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Sadek, E.S.; Makris, D.P.; Kefalas, P. Polyphenolic Composition and Antioxidant Characteristics of Kumquat (Fortunella margarita) Peel Fractions. Plant Foods Hum. Nutr. 2009, 64, 297–302. [Google Scholar] [CrossRef]
- Abdulai, I.L.; Kwofie, S.K.; Gbewonyo, W.S.; Boison, D.; Puplampu, J.B.; Adinortey, M.B. Multitargeted Effects of Vitexin and Isovitexin on Diabetes Mellitus and Its Complications. Sci. World J. 2021, 2021, 6641128. [Google Scholar] [CrossRef] [PubMed]
- Li, L.Z.; Gao, P.Y.; Song, S.J.; Yuan, Y.Q.; Liu, C.T.; Huang, X.X.; Liu, Q.B. Monoterpenes and Flavones from the Leaves of Crataegus pinnatifida with Anticoagulant Activities. J. Funct. Foods 2015, 12, 237–245. [Google Scholar] [CrossRef]
- Lv, S.X.; Qiao, X. Isovitexin (IV) Induces Apoptosis and Autophagy in Liver Cancer Cells through Endoplasmic Reticulum Stress. Biochem. Biophys. Res. Commun. 2018, 496, 1047–1054. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, Q.; Liu, H.; Luo, S. Vitexin Induces Apoptosis through Mitochondrial Pathway and PI3K/Akt/MTOR Signaling in Human Non-Small Cell Lung Cancer A549 Cells. Biol. Res. 2019, 52, 7. [Google Scholar] [CrossRef]
- Zhou, P.; Zheng, Z.H.; Wan, T.; Wu, J.; Liao, C.W.; Sun, X.J. Vitexin Inhibits Gastric Cancer Growth and Metastasis through HMGB1-Mediated Inactivation of the PI3K/AKT/MTOR/HIF-1α Signaling Pathway. J. Gastric Cancer 2021, 21, 439–456. [Google Scholar] [CrossRef]
- Gennari, L.; Felletti, M.; Blasa, M.; Angelino, D.; Celeghini, C.; Corallini, A.; Ninfali, P. Total Extract of Beta vulgaris Var. cicla Seeds versus Its Purified Phenolic Components: Antioxidant Activities and Antiproliferative Effects against Colon Cancer Cells. Phytochem. Anal. 2011, 22, 272–279. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, X.; Zhang, W.; Rengarajan, T. Vicenin-2 Inhibits Wnt/β-Catenin Signaling and Induces Apoptosis in HT-29 Human Colon Cancer Cell Line. Drug Des. Devel. Ther. 2018, 12, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Nagaprashantha, L.D.; Vatsyayan, R.; Singhal, J.; Fast, S.; Roby, R.; Awasthi, S.; Singhal, S.S. Anti-Cancer Effects of Novel Flavonoid Vicenin-2 as a Single Agent and in Synergistic Combination with Docetaxel in Prostate Cancer. Biochem. Pharmacol. 2011, 82, 1100–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zheng, Y.; Wang, H. Anticancer Activity of Vicenin-2 against 7,12 Dimethylbenz[a]Anthracene-Induced Buccal Pouch Carcinoma in Hamsters. J. Biochem. Mol. Toxicol. 2021, 35, e22673. [Google Scholar] [CrossRef]
- Singhal, S.S.; Jain, D.; Singhal, P.; Awasthi, S.; Singhal, J.; Horne, D. Targeting the Mercapturic Acid Pathway and Vicenin-2 for Prevention of Prostate Cancer. BBA—Rev. Cancer 2017, 1868, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | mg kg−1 Dry Extract | |
|---|---|---|
| 1 | Vicenin-2 | 266 ± 40 |
| 2 | Vicenin-2 derivative | 461 ± 14 |
| 3 | Vicenin-2 derivative | 410 ± 17 |
| 4 | 2″-O-Rhamnosyl vitexin | 523 ± 3 |
| 5 | 2″-O-Rhamnosyl vitexin derivative | 1081 ± 2 |
| Total | 2741 ± 76 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, V.; Ferreres, F.; Oliveira, A.; Gomes, N.G.M.; Gil-Izquierdo, Á.; Araújo, L.; Pereira, D.; Valentão, P. Allophylus africanus Stem Bark Extract Modulates the Mitochondrial Apoptotic Pathway in Human Stomach Cancer Cells. Life 2023, 13, 406. https://doi.org/10.3390/life13020406
Ribeiro V, Ferreres F, Oliveira A, Gomes NGM, Gil-Izquierdo Á, Araújo L, Pereira D, Valentão P. Allophylus africanus Stem Bark Extract Modulates the Mitochondrial Apoptotic Pathway in Human Stomach Cancer Cells. Life. 2023; 13(2):406. https://doi.org/10.3390/life13020406
Chicago/Turabian StyleRibeiro, Vera, Federico Ferreres, Andreia Oliveira, Nelson G. M. Gomes, Ángel Gil-Izquierdo, Luísa Araújo, David Pereira, and Patrícia Valentão. 2023. "Allophylus africanus Stem Bark Extract Modulates the Mitochondrial Apoptotic Pathway in Human Stomach Cancer Cells" Life 13, no. 2: 406. https://doi.org/10.3390/life13020406