
Antiobesity and Antioxidative Effect of Fermented Brown Rice Using In Vitro with In Vivo Caenorhabditis elegans Model

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Culture Media
2.2. Bacterial Strains Growth Conditions and Preparation
2.3. Preparation of Brown Rice and Fermentation
2.4. Pancreatic Lipase Inhibition Assay
2.5. Determination of Total Phenolic Content (TPC)
2.6. Antioxidant Activity
2.6.1. DPPH and ABTS Radical Scavenging Effect
2.6.2. Ferric Reducing Antioxidant Power (FRAP)
2.7. Ferulic Acid (FA) Detection in Raw and Fermented Samples Using HPLC
2.8. Maintenance, Synchronization, and Establishment of a Glucose Diet in Caenorhabditis elegans
2.8.1. Lifespan Assay for Detecting FBR Effects in the C. elegans Model
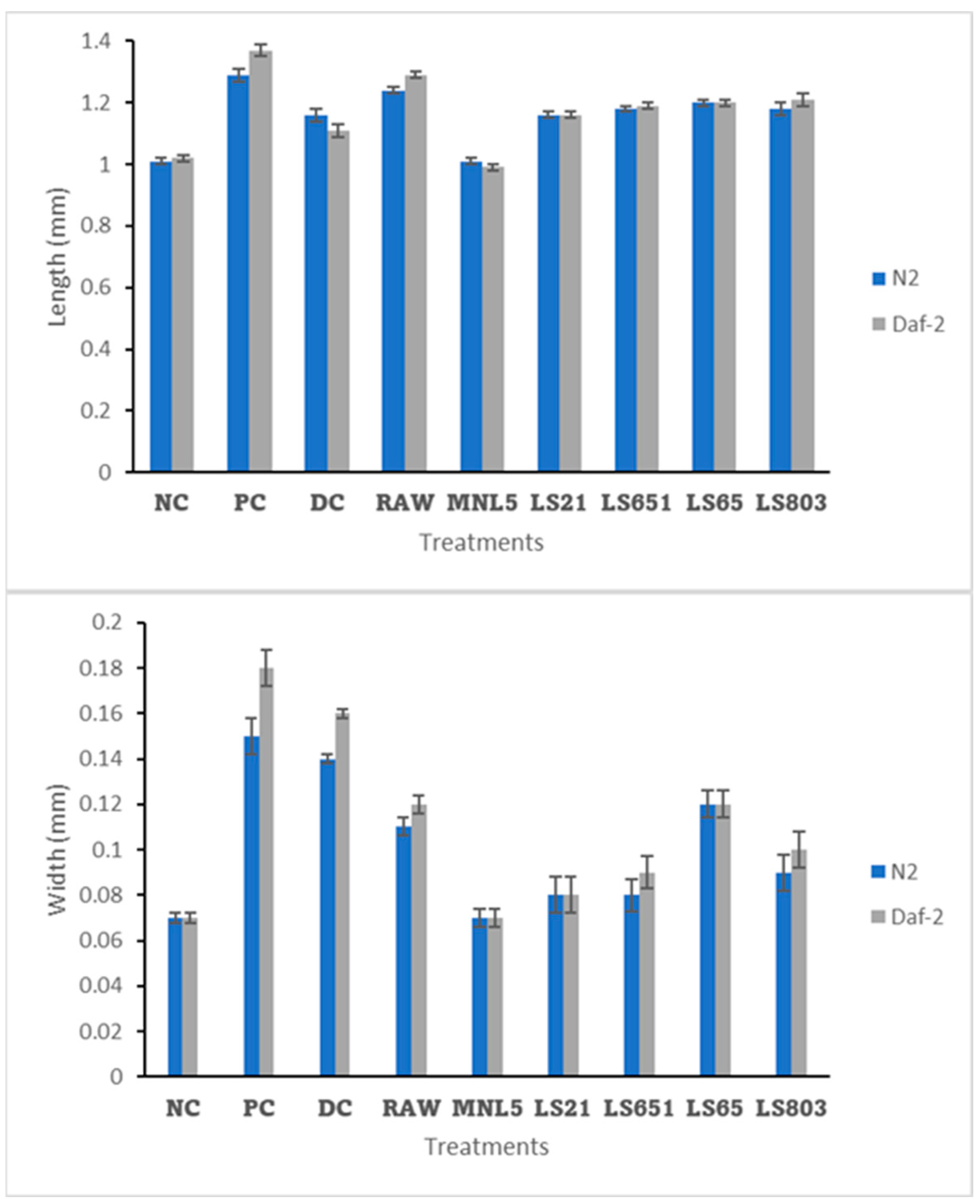
2.8.2. Impact of FBR C. elegans Body Size
2.8.3. Analysis of Lipid Deposition in C. elegans Using a Fluorescence Microscope
2.8.4. Fluorescence Quantification
2.8.5. Determining the Triglycerides
2.9. Statistical and Software Analysis
3. Results
3.1. Effect of Brown Rice Fermentation
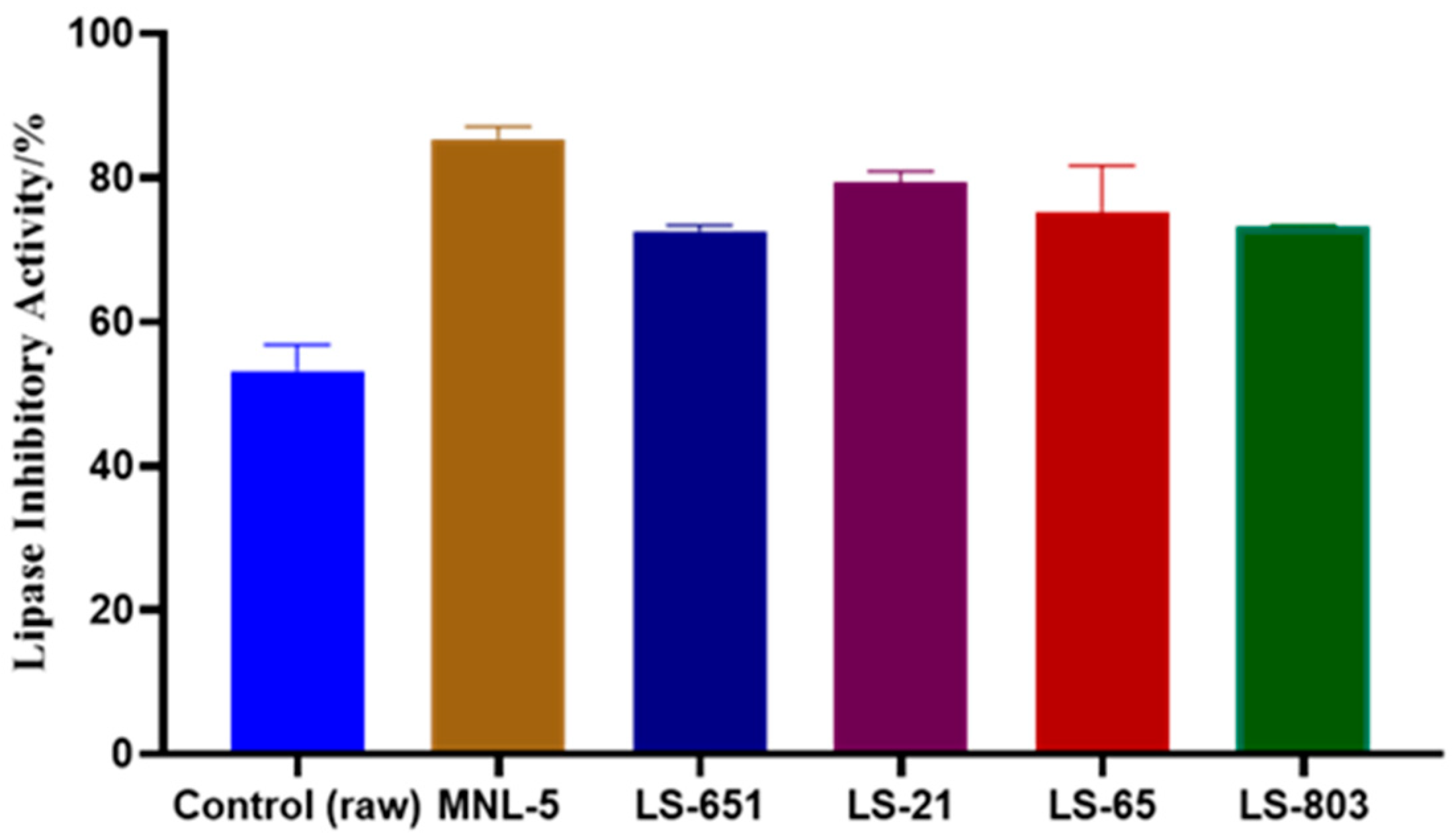
3.2. Impact of RBR and FBR Extracts on Lipase Inhibition
3.3. Total Phenolic Content of RBR and FBR Extracts
3.4. Antioxidant Assay (DPPH, ABTS, and FRAP) of RBR and FBR Extracts
The Effect of DPPH, ABTS, and FRAP Scavenging Activity
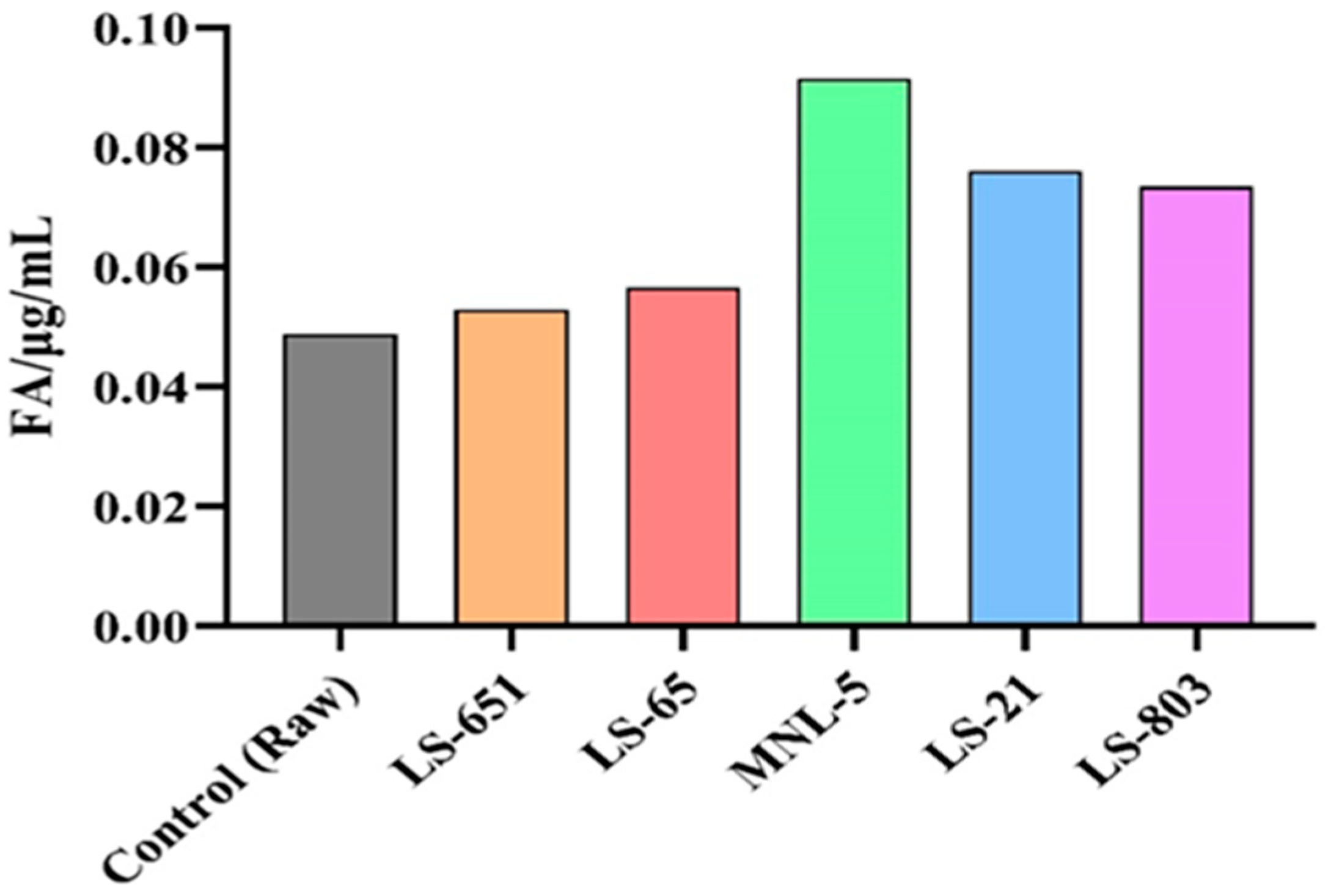
3.5. Optimization of the Detection of Ferulic Acid
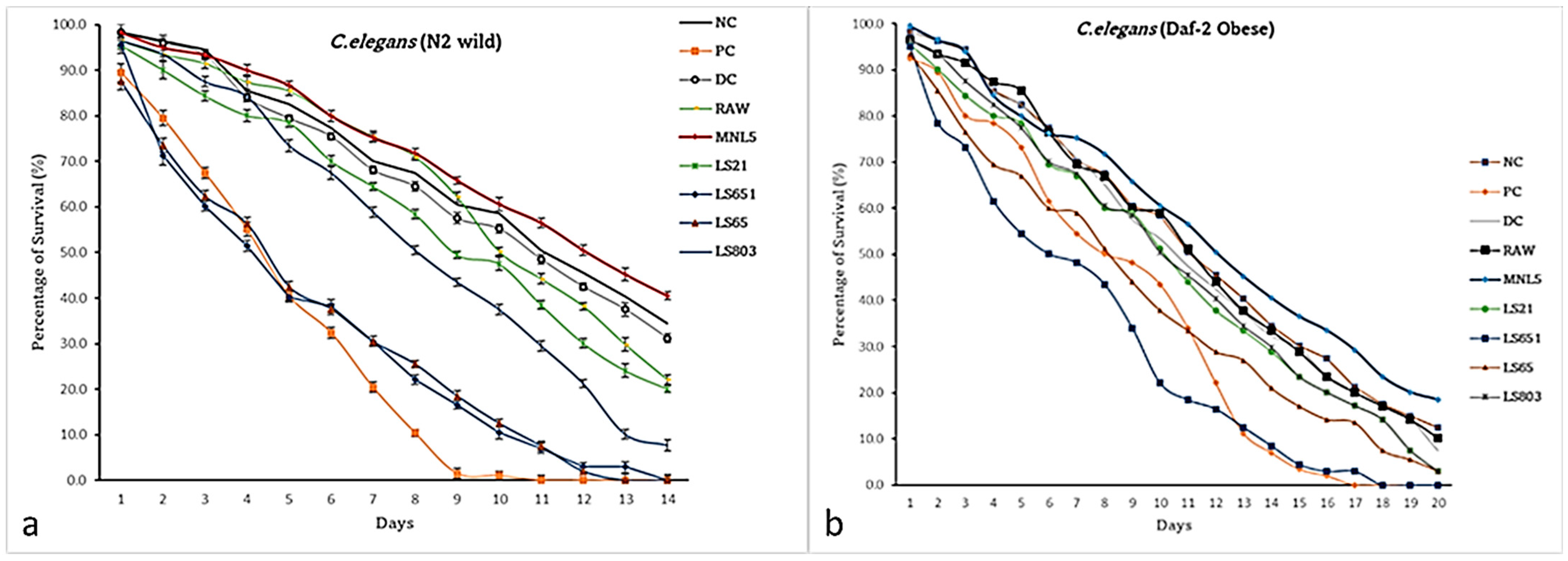
3.6. Effects of RBR and FBR Extracts on the Life Span of C. elegans
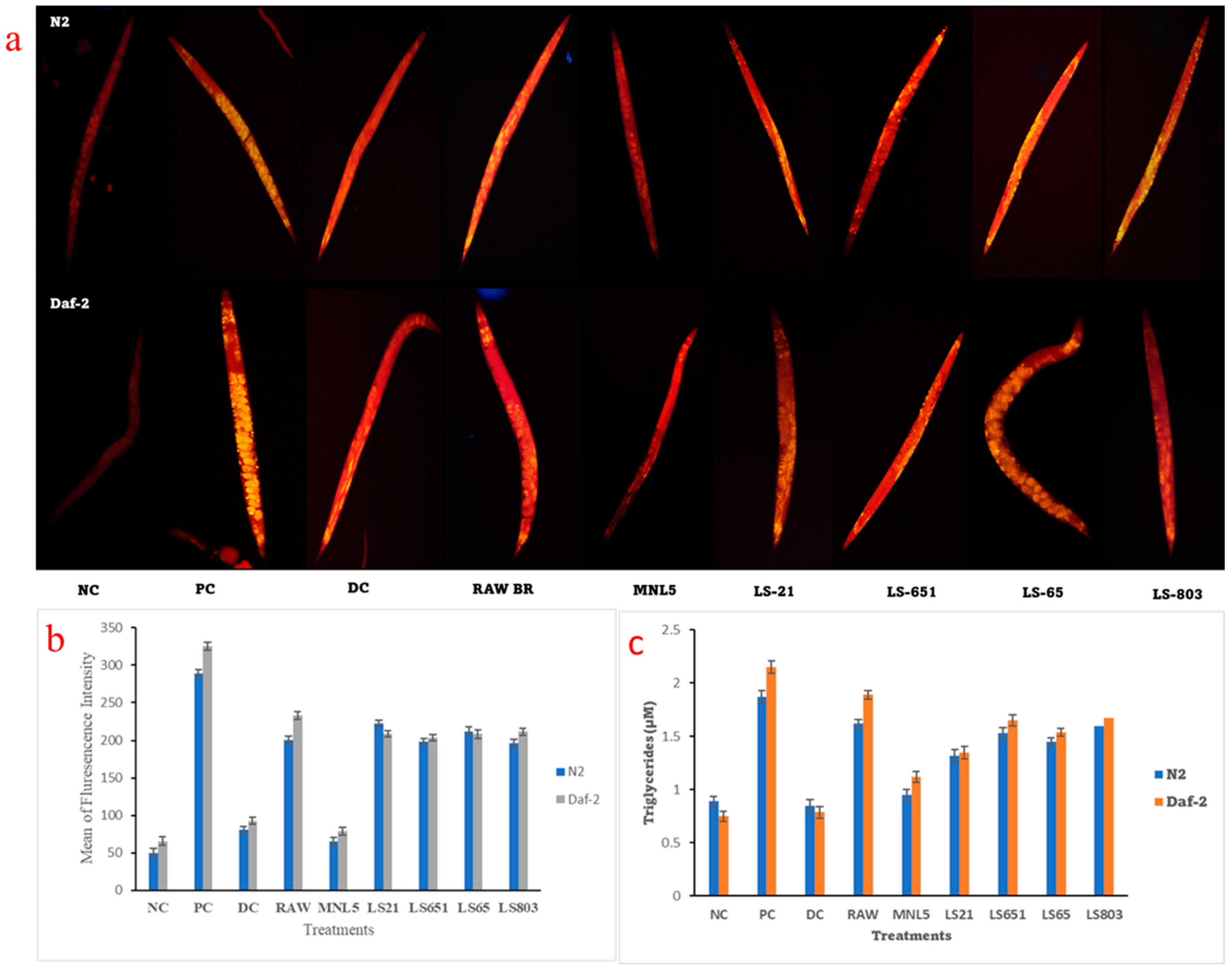
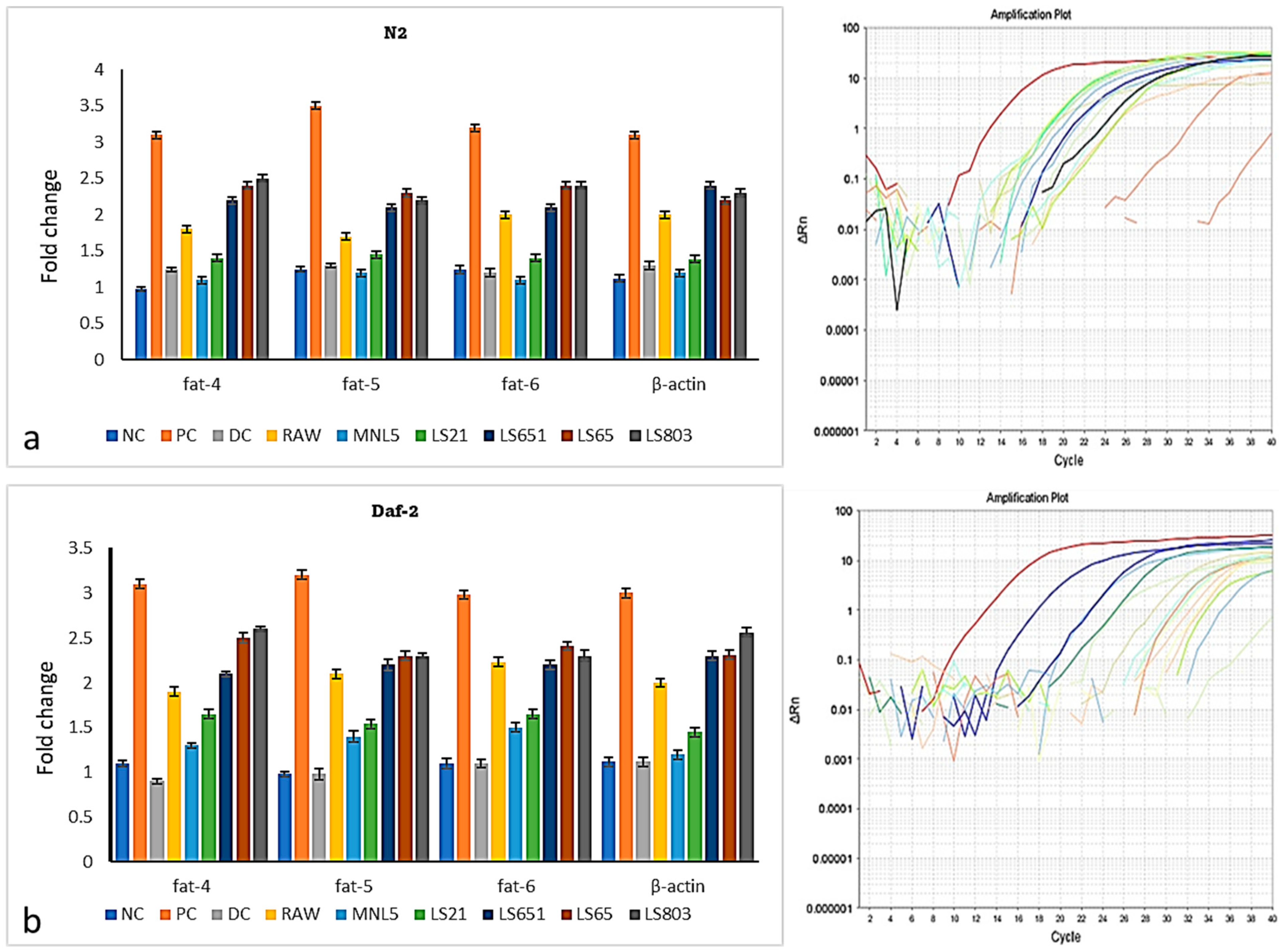
Effects of RBR and FBR Extract on Fat Deposition, Triglyceride (TG) Content, and Gene Expression Levels of C. elegans
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- World Health Organization. World Obesity Day 2022–Accelerating Action to Stop Obesity. 2022. Available online: https://www.who.int/news/item/04-03-2022-world-obesity-day-2022-accelerating-action-to-stop-obesity (accessed on 22 December 2022).
- Mohan, V.; Ruchi, V.; Gayathri, R.; Ramya Bai, M.; Shobana, S.; Anjana, R.; Unnikrishnan, R.; Sudha, V. Hurdles in brown rice consumption. In Brown Rice; Springer: Berlin/Heidelberg, Germany, 2017; pp. 255–269. [Google Scholar]
- Tyagi, A.; Shabbir, U.; Chelliah, R.; Daliri, E.B.-M.; Chen, X.; Oh, D.-H. Limosilactobacillus reuteri Fermented Brown Rice: A Product with Enhanced Bioactive Compounds and Antioxidant Potential. Antioxidants 2021, 10, 1077. [Google Scholar] [CrossRef] [PubMed]
- Ohsaki, Y.; Shirakawa, H.; Koseki, T.; Komai, M. Novel effects of a single administration of ferulic acid on the regulation of blood pressure and the hepatic lipid metabolic profile in stroke-prone spontaneously hypertensive rats. J. Agric. Food Chem. 2008, 56, 2825–2830. [Google Scholar]
- Tyagi, A.; Lim, M.-J.; Kim, N.-H.; Barathikannan, K.; Vijayalakshmi, S.; Elahi, F.; Ham, H.-J.; Oh, D.-H. Quantification of Amino Acids, Phenolic Compounds Profiling from Nine Rice Varieties and Their Antioxidant Potential. Antioxidants 2022, 11, 839. [Google Scholar] [CrossRef] [PubMed]
- Panneerselvam, P.; Binodh, A.K.; Kumar, U.; Sugitha, T.; Anandan, A. Microbial association in brown rice and their influence on human health. In Brown Rice; Springer: Berlin/Heidelberg, Germany, 2017; pp. 159–181. [Google Scholar]
- Gélinas, P.; McKinnon, C. Fermentation and microbiological processes in cereal foods. In Handbook of Cereal Science and Technology; CRC Press: Boca Raton, FL, USA, 2000; pp. 741–754. [Google Scholar]
- Yang, S.-T. Bioprocessing for Value-Added Products from Renewable Resources: New Technologies and Applications; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Jemil, I.; Mora, L.; Nasri, R.; Abdelhedi, O.; Aristoy, M.-C.; Hajji, M.; Nasri, M.; Toldrá, F. A peptidomic approach for the identification of antioxidant and ACE-inhibitory peptides in sardinelle protein hydrolysates fermented by Bacillus subtilis A26 and Bacillus amyloliquefaciens An6. Food Res. Int. 2016, 89, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Wilson, T.A.; Nicolosi, R.J.; Woolfrey, B.; Kritchevsky, D. Rice bran oil and oryzanol reduce plasma lipid and lipoprotein cholesterol concentrations and aortic cholesterol ester accumulation to a greater extent than ferulic acid in hypercholesterolemic hamsters. J. Nutr. Biochem. 2007, 18, 105–112. [Google Scholar] [CrossRef]
- Yi, C.; Xie, L.; Cao, Z.; Quan, K.; Zhu, H.; Yuan, J. Effects of rice bran fermented with Lactobacillus plantarum on palatability, volatile profiles, and antioxidant activity of brown rice noodles. Int. J. Food Sci. Technol. 2022, 57, 5048–5056. [Google Scholar] [CrossRef]
- Son, M.J.; Rico, C.W.; Nam, S.H.; Kang, M.Y. Effect of oryzanol and ferulic acid on the glucose metabolism of mice fed with a high-fat diet. J. Food Sci. 2011, 76, H7–H10. [Google Scholar] [CrossRef]
- Jones, K.T.; Ashrafi, K. Caenorhabditis elegans as an emerging model for studying the basic biology of obesity. Dis. Model. Mech. 2009, 2, 224–229. [Google Scholar] [CrossRef] [Green Version]
- Ashrafi, K.; Chang, F.Y.; Watts, J.L.; Fraser, A.G.; Kamath, R.S.; Ahringer, J.; Ruvkun, G. Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes. Nature 2003, 421, 268–272. [Google Scholar] [CrossRef]
- Li, H.; Black, P.N.; DiRusso, C.C. A live-cell high-throughput screening assay for identification of fatty acid uptake inhibitors. Anal. Biochem. 2005, 336, 11–19. [Google Scholar] [CrossRef]
- Aloo, S.O.; Ofosu, F.K.; Oh, D.H. Effect of Germination on Alfalfa and Buckwheat: Phytochemical Profiling by UHPLC-ESI-QTOF-MS/MS, Bioactive Compounds, and In-Vitro Studies of Their Diabetes and Obesity-Related Functions. Antioxidants 2021, 10, 1613. [Google Scholar] [CrossRef]
- Glorybai, L.; Kannan K, B.; Arasu, M.V.; Al-Dhabi, N.A.; Agastian, P. Some biological activities of Epaltes divaricata L.-an in vitro study. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, A.; Chelliah, R.; Banan-Mwine Daliri, E.; Sultan, G.; Madar, I.H.; Kim, N.; Shabbir, U.; Oh, D.-H. Antioxidant activities of novel peptides from Limosilactobacillus reuteri fermented brown rice: A combined in vitro and in silico study. Food Chem. 2023, 404, 134747. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Zhang, M.; Apea-Bah, F.B.; Beta, T. Hydroxycinnamic acid amide (HCAA) derivatives, flavonoid C-glycosides, phenolic acids and antioxidant properties of foxtail millet. Food Chem. 2019, 295, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Nadal, J.M.; Toledo, M.d.G.; Pupo, Y.M.; Padilha de Paula, J.; Farago, P.V.; Zanin, S.M.W. A stability-indicating hplc-dad method for determination of ferulic acid into microparticles: Development, validation, forced degradation, and encapsulation efficiency. J. Anal. Methods Chem. 2015, 2015, 286812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barathikannan, K.; Chelliah, R.; Elahi, F.; Tyagi, A.; Selvakumar, V.; Agastian, P.; Valan Arasu, M.; Oh, D.-H. Anti-Obesity Efficacy of Pediococcus acidilactici MNL5 in Canorhabditis elegans Gut Model. Int. J. Mol. Sci. 2022, 23, 1276. [Google Scholar] [CrossRef]
- Barathikannan, K.; Chelliah, R.; Yeon, S.-J.; Tyagi, A.; Elahi, F.; Vijayalakshmi, S.; Agastian, P.; Arockiasami, V.; Hawn Oh, D. Untargeted metabolomics of fermented onion (Allium cepa L) using UHPLC Q-TOF MS/MS reveals antiobesity metabolites and in vivo efficacy in Caenorhabditis elegans. Food Chem. 2023, 404, 134710. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.-H.; Jeong, J.; Kim, H. Synergistic effects of heat-killed kefir paraprobiotics and flavonoid-rich prebiotics on western diet-induced obesity. Nutrients 2020, 12, 2465. [Google Scholar] [CrossRef]
- Karamać, M.; Amarowicz, R. Inhibition of pancreatic Lipase by phenolic acids-examination in vitro. Z. Für Nat. C 1996, 51, 903–906. [Google Scholar] [CrossRef]
- Dai, T.; Chen, J.; McClements, D.J.; Hu, P.; Ye, X.; Liu, C.; Li, T. Protein–polyphenol interactions enhance the antioxidant capacity of phenolics: Analysis of rice glutelin–procyanidin dimer interactions. Food Funct. 2019, 10, 765–774. [Google Scholar] [CrossRef]
- Janarny, G.; Gunathilake, K. Changes in rice bran bioactives, their bioactivity, bioaccessibility and bioavailability with solid-state fermentation by Rhizopus oryzae. Biocatal. Agric. Biotechnol. 2020, 23, 101510. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.; Juraimi, A.S.; Tayebi-Meigooni, A. Comparative evaluation of different extraction techniques and solvents for the assay of phytochemicals and antioxidant activity of hashemi rice bran. Molecules 2015, 20, 10822–10838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilowefah, M.; Bakar, J.; Ghazali, H.M.; Muhammad, K. Enhancement of nutritional and antioxidant properties of brown rice flour through solid-state yeast fermentation. Cereal Chem. 2017, 94, 519–523. [Google Scholar] [CrossRef]
- Georgiev, V.G.; Weber, J.; Kneschke, E.-M.; Denev, P.N.; Bley, T.; Pavlov, A.I. Antioxidant activity and phenolic content of betalain extracts from intact plants and hairy root cultures of the red beetroot Beta vulgaris cv. Detroit dark red. Plant Foods Hum. Nutr. 2010, 65, 105–111. [Google Scholar] [CrossRef]
- De Melo, T.; Lima, P.; Carvalho, K.; Fontenele, T.; Solon, F.; Tomé, A.; De Lemos, T.; da Cruz Fonseca, S.; Santos, F.; Rao, V. Ferulic acid lowers body weight and visceral fat accumulation via modulation of enzymatic, hormonal and inflammatory changes in a mouse model of high-fat diet-induced obesity. Braz. J. Med. Biol. Res. 2017, 50, e5630. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sair, A.T.; Zhao, W.; Li, T.; Liu, R.H. Ferulic Acid Mediates Metabolic Syndrome via the Regulation of Hepatic Glucose and Lipid Metabolisms and the Insulin/IGF-1 Receptor/PI3K/AKT Pathway in Palmitate-Treated HepG2 Cells. J. Agric. Food Chem. 2022, 70, 14706–14717. [Google Scholar] [CrossRef]
- Naowaboot, J.; Piyabhan, P.; Munkong, N.; Parklak, W.; Pannangpetch, P. Ferulic acid improves lipid and glucose homeostasis in high-fat diet-induced obese mice. Clin. Exp. Pharmacol. Physiol. 2016, 43, 242–250. [Google Scholar] [CrossRef]
- Ye, L.; Hu, P.; Feng, L.P.; Huang, L.L.; Wang, Y.; Yan, X.; Xiong, J.; Xia, H.L. Protective Effects of Ferulic Acid on Metabolic Syndrome: A Comprehensive Review. Molecules 2023, 28, 281. [Google Scholar] [CrossRef]
- Moreno-Arriola, E.; Cárdenas-Rodríguez, N.; Coballase-Urrutia, E.; Pedraza-Chaverri, J.; Carmona-Aparicio, L.; Ortega-Cuellar, D. Caenorhabditis elegans: A useful model for studying metabolic disorders in which oxidative stress is a contributing factor. Oxidative Med. Cell. Longev. 2014, 2014, 705253. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Jiang, M.; Guo, Q.; Shi, W. Zymolytic Grain Extract (ZGE) Significantly Extends the Lifespan and Enhances the Environmental Stress Resistance of Caenorhabditis elegans. Int. J. Mol. Sci. 2019, 20, 3489. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, L.; Zhang, L.; Wang, W.; Wei, S.; Wang, J.; Che, H.; Zhang, Y. Effects of excess sugars and lipids on the growth and development of Caenorhabditis elegans. Genes Nutr. 2020, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Salgueiro, W.; Bianchini, M.; Veit, J.C.; Puntel, R.L.; Emanuelli, T.; Dernadin, C.C.; Ávila, D.S. Salvia hispanica L.(chia) seeds oil extracts reduce lipid accumulation and produce stress resistance in Caenorhabditis elegans. Nutr. Metab. 2018, 15, 83. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No | Raw and Fermented Brown Rice Strains | DPPH (mg Trolox Equiv./100 g, DW) | ABTS (mg Trolox Equiv./100 g, DW) | FRAP (mg Trolox Equiv./100 g, DW) | TPC (mg Gallic Acid Equiv./100 g, DW) |
|---|---|---|---|---|---|
| 1 | Control (Raw) | 55.65 ± 0.73 e | 49.54 ± 0.92 e | 58.02 ± 0.96 d | 288.83 ± 0.61 f |
| 2 | MNL-5 | 124.40 ± 0.87 a | 136.45 ± 1.0 a | 120.32 ± 0.64 a | 650.79 ± 0.84 a |
| 3 | LS-651 | 68.12 ± 1.01 d | 82.45 ± 0.71 d | 72.31 ± 0.59 c | 569.41 ± 0.73 b |
| 4 | LS-21 | 107.31 ± 0.94 b | 108.32 ± 0.83 b | 100.30 ± 1.0 b | 476.21 ± 1.03 e |
| 5 | LS-65 | 100.10 ± 0.59 b | 97.74 ± 0.74 b | 95.45 ± 0.88 b | 537.45 ± 0.89 c |
| 6 | LS-803 | 90.64 ± 0.99 c | 92.31 ± 0.55 c | 82.31 ± 0.97 c | 493.68 ± 0.51 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barathikannan, K.; Tyagi, A.; Shan, L.; Kim, N.-H.; Lee, D.-S.; Park, J.-S.; Chelliah, R.; Oh, D.-H. Antiobesity and Antioxidative Effect of Fermented Brown Rice Using In Vitro with In Vivo Caenorhabditis elegans Model. Life 2023, 13, 374. https://doi.org/10.3390/life13020374
Barathikannan K, Tyagi A, Shan L, Kim N-H, Lee D-S, Park J-S, Chelliah R, Oh D-H. Antiobesity and Antioxidative Effect of Fermented Brown Rice Using In Vitro with In Vivo Caenorhabditis elegans Model. Life. 2023; 13(2):374. https://doi.org/10.3390/life13020374
Chicago/Turabian StyleBarathikannan, Kaliyan, Akanksha Tyagi, Lingyue Shan, Nam-Hyeon Kim, Deuk-Sik Lee, Jong-Soon Park, Ramachandran Chelliah, and Deog-Hwan Oh. 2023. "Antiobesity and Antioxidative Effect of Fermented Brown Rice Using In Vitro with In Vivo Caenorhabditis elegans Model" Life 13, no. 2: 374. https://doi.org/10.3390/life13020374