

Using the Gut Microbiome to Assess Stocking Efforts of the Endangered Pallid Sturgeon, Scaphirhynchus albus
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Sequencing
2.2. Core Gut Microbiome
2.3. Comparison between Hatchery-Raised and Wild Pallid Sturgeon Gut Microbiomes
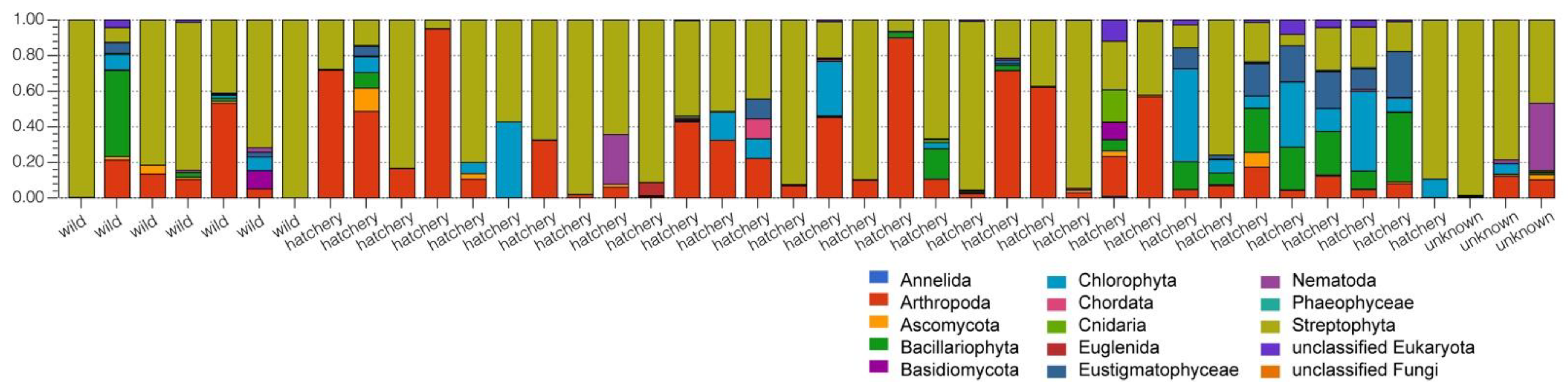
3. Results
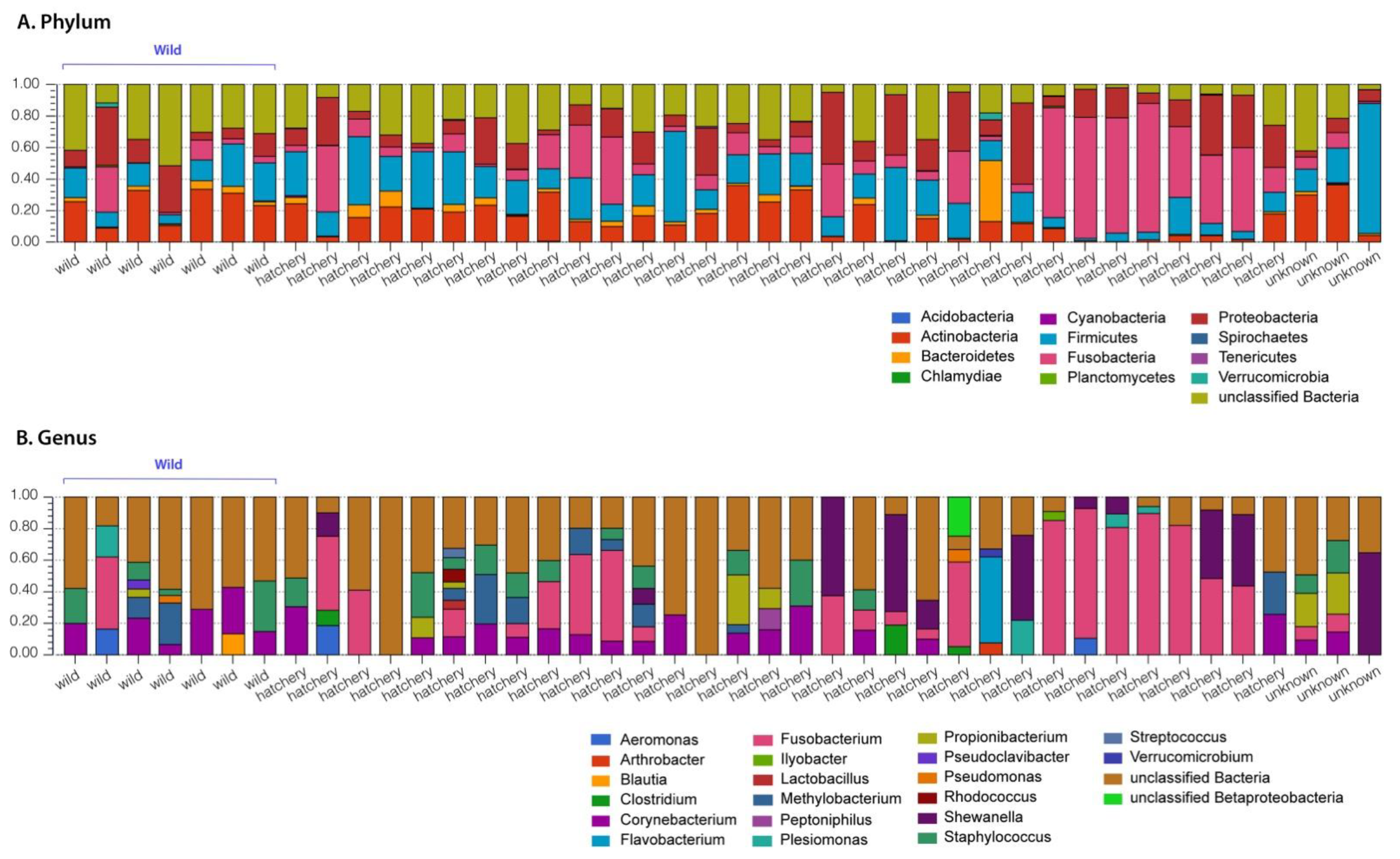
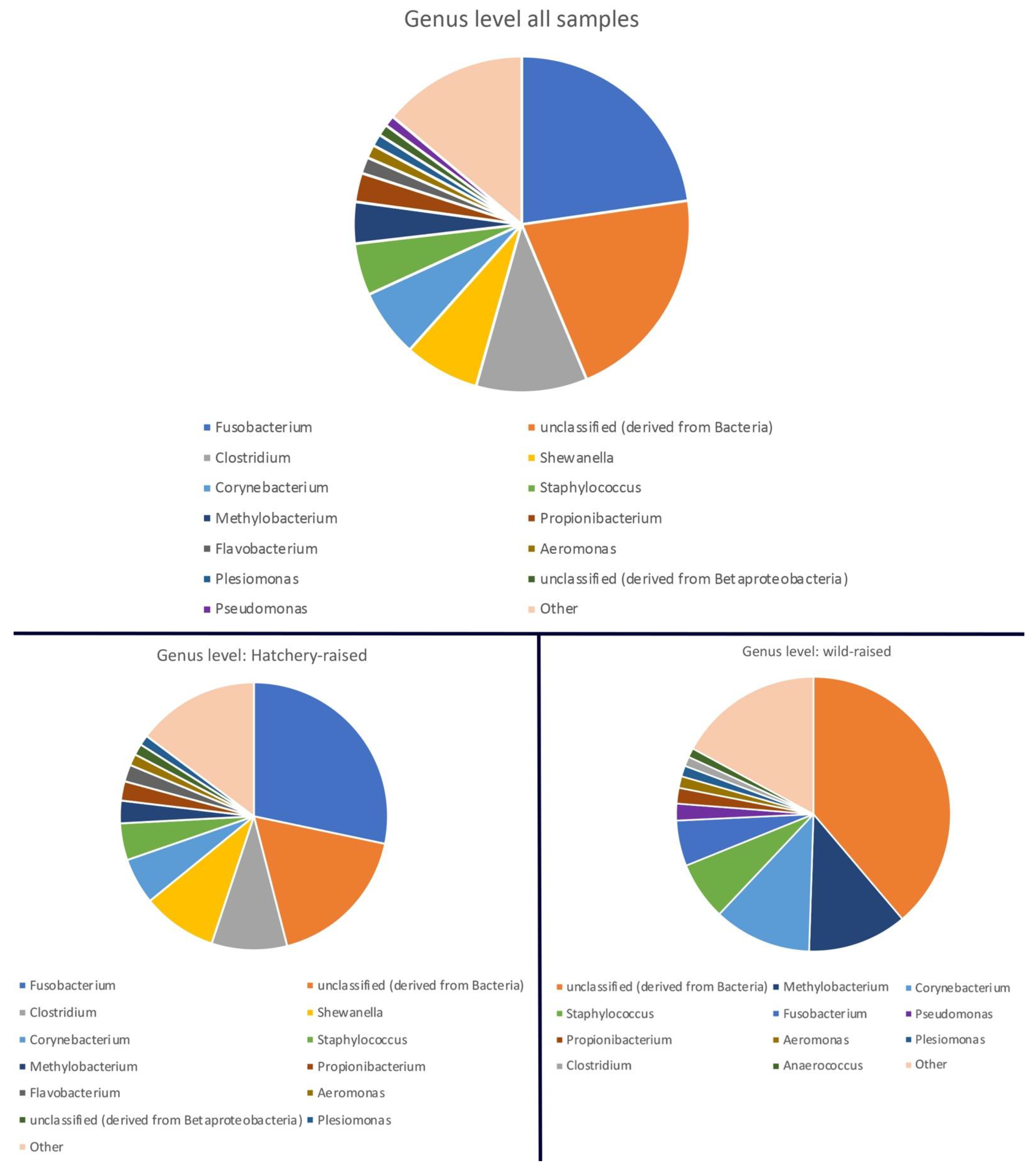
3.1. Pallid Sturgeon Gut Microbiome
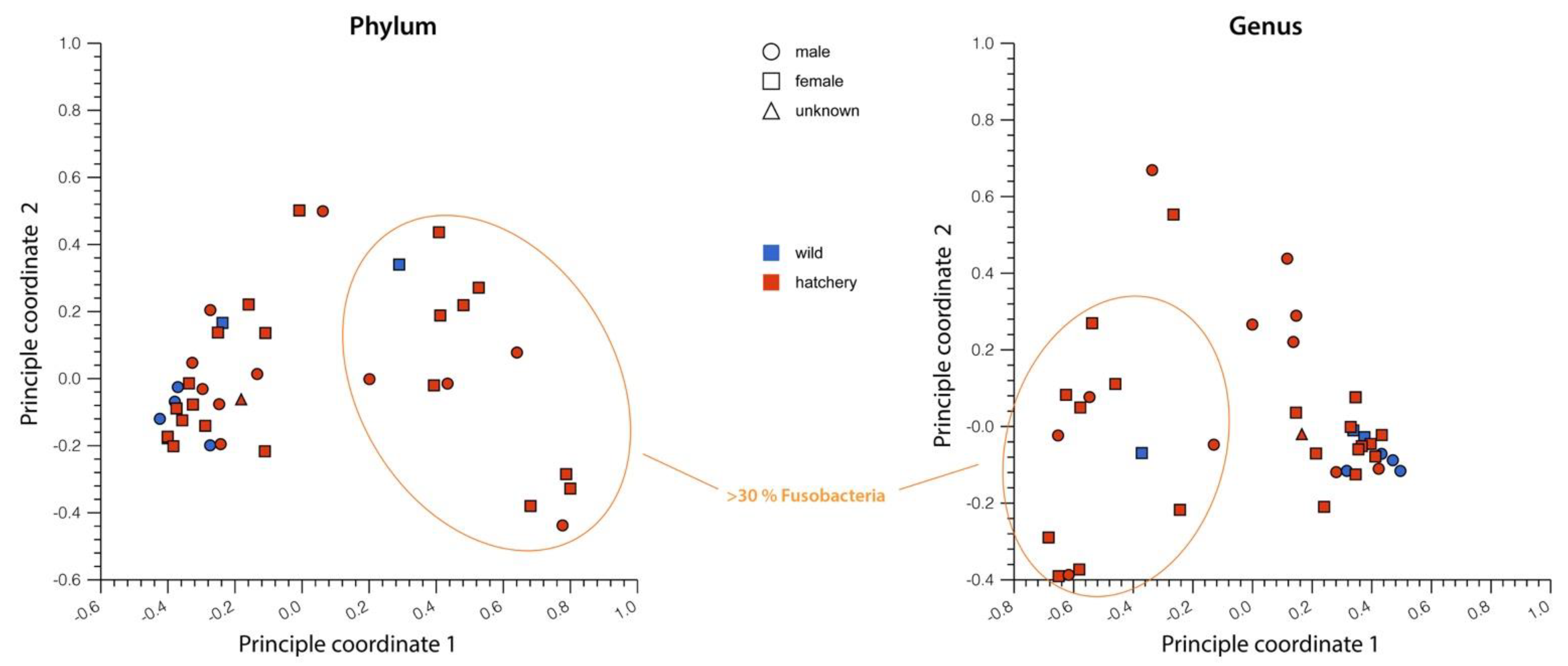
3.2. Comparison between Hatchery-Raised and Wild Pallid Sturgeon Gut Microbiomes
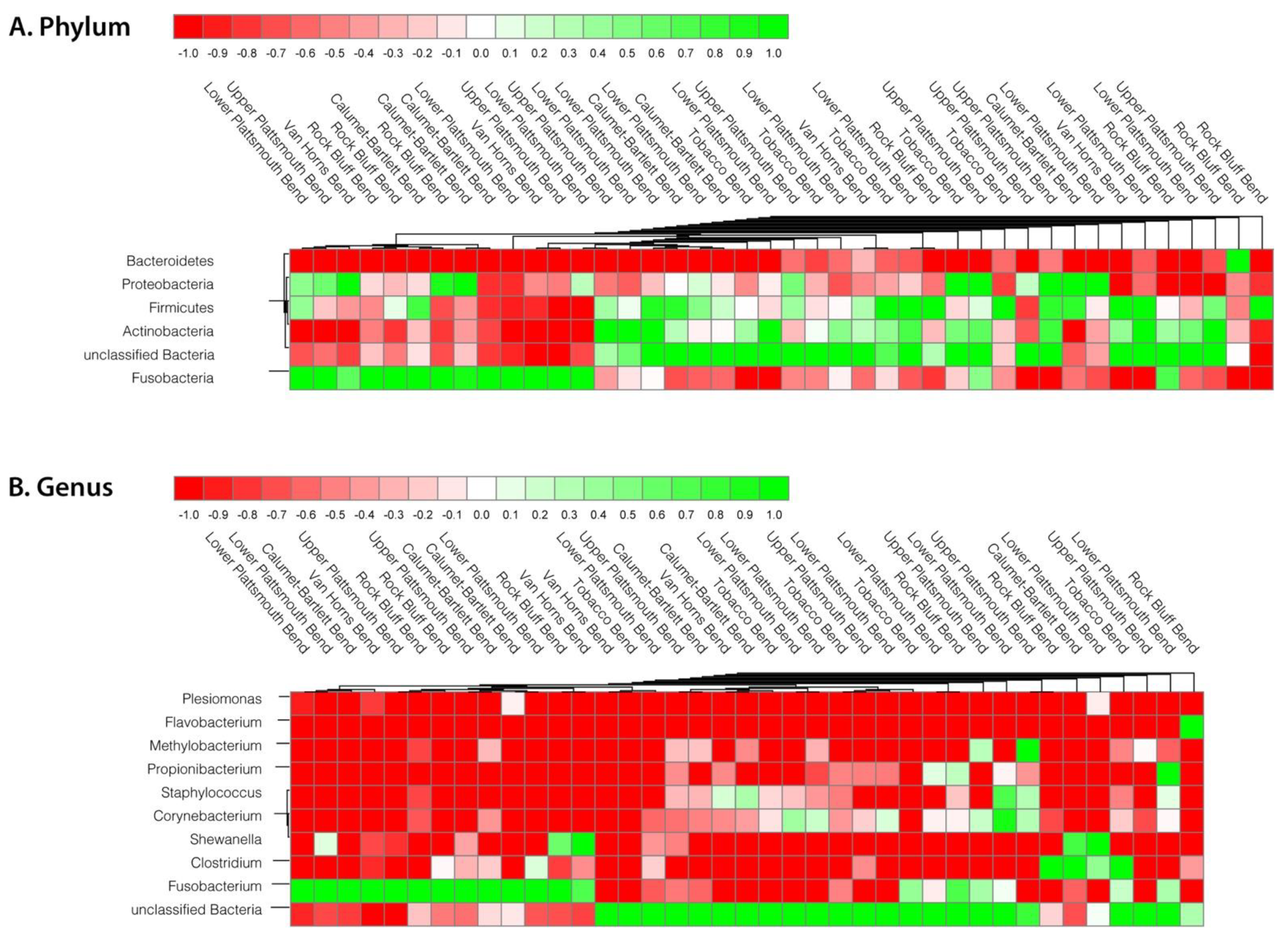
3.3. Other Factors That May Affect Microbiome Variation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, K.H.; Blitchington, R.B. Human colonic biota studied by ribosomal DNA sequence analysis. Appl. Environ. Microbiol. 1996, 62, 2273–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207. [Google Scholar]
- O’Hara, A.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, M.M. Bacterial flora of fishes: A review. Microb. Ecol. 1990, 19, 21–41. [Google Scholar] [CrossRef]
- Ringø, E.; Strøm, E.; Tabachek, J.A. Intestinal microflora of salmonids: A review. Aquac. Res. 1995, 26, 773–789. [Google Scholar] [CrossRef]
- Kim, P.S.; Shin, N.R.; Lee, J.B.; Kim, M.-S.; Whon, T.W.; Hyun, D.-W.; Yun, J.-H.; Jung, M.-J.; Kim, J.Y.; Bae, J.-W. Host habitat is the major determinant of the gut microbiome of fish. Microbiome 2021, 9, 166. [Google Scholar] [CrossRef]
- Sullam, K.E.; Essinger, S.D.; Lozupone, C.A.; O’Connor, M.P.; Rosen, G.L.; Knight, R.; Kilham, S.S.; Russell, J.A. Environmental and ecological factors that shape the gut bacterial communities of fish: A meta-analysis. Mol. Ecol. 2012, 21, 3363–3378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talwar, C.; Nagar, S.; Lal, R.; Negi, R.K. Fish gut microbiome: Current approaches and future perspectives. Ind. J. Microbiol. 2018, 58, 397–414. [Google Scholar] [CrossRef]
- Baldo, L.; Riera, J.L.; Tooming-Klunderud, A.; Albà, M.M.; Salzburger, W. Gut microbiota dynamics during dietary shift in eastern African cichlid fishes. PLoS ONE 2015, 10, e0127462. [Google Scholar] [CrossRef] [Green Version]
- Eichmiller, J.J.; Hamilton, M.J.; Staley, C.; Sadowsky, M.J.; Sorensen, P.W. Environment shapes the fecal microbiome of invasive carp species. Microbiome 2016, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.; Rawls, J.F. Intestinal microbiota composition in fishes is influenced by host ecology and environment. Mol. Ecol. 2012, 21, 3100–3102. [Google Scholar] [CrossRef] [PubMed]
- Taneja, V. Microbiome: Impact of gender on function & characteristics of gut microbiome. In Principles of Gender-Specific Medicine; Academic Press: Cambridge, MA, USA, 2017; pp. 569–583. [Google Scholar]
- Li, X.; Yan, Q.; Ringø, E.; Wu, X.; He, Y.; Yang, D. The influence of weight and gender on intestinal bacterial community of wild largemouth bronze gudgeon (Coreius guichenoti, 1874). BMC Microbiol. 2016, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dryer, M.; Sandvol, A. Recovery Plan for the Pallid Sturgeon (Scaphirhynchus albus); U.S. Fish and Wildlife Service: Washington, DC, USA, 1993. [Google Scholar]
- Forbes, S.A.; Richardson, R.E. On a new Shovelnose Sturgeon from the Mississippi River. Ill. Nat. Hist. Surv. Bull. 1905, 7, 1–10. [Google Scholar] [CrossRef]
- Brown, C.; Day, R.L. The future of stock enhancements: Lessons for hatchery practice from conservation biology. Fish Fish. 2002, 3, 79–94. [Google Scholar] [CrossRef] [Green Version]
- Ruskamp, R.L. Central Lowlands and Interior Highlands Pallid Sturgeon Spawning and Stocking Summary; Nebraska Game and Parks Commission Internal Document; Nebraska Game and Parks Commission: Lincoln, NE, USA, 2021. [Google Scholar]
- Steffensen, K.D.; Chojnacki, K.A.; Kalie, J.A.; Barton, M.L.; Heist, E.J.; Winders, K.R.; Loecher, N.C.; Doyle, W.J.; Welker, T.L. Evidence of Limited Recruitment of Pallid Sturgeon in the Lower Missouri River. J. Fish Wildl. Manag. 2019, 10, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Hoover, J.J.; George, S.G.; Killgore, K.J. Diet of shovelnose sturgeon and pallid sturgeon in the free-flowing Mississippi River. J. Appl. Ichthyol. 2007, 23, 494–499. [Google Scholar] [CrossRef]
- Braaten, P.J.; Fuller, D.B.; Lott, R.D.; Haddix, T.M.; Holte, L.D.; Wilson, R.H.; Bartron, M.L.; Kalie, J.A.; DeHaan, W.R.; Ardren, R.J.; et al. Natural growth and diet of known-age pallid sturgeon (Scaphirhynchus albus) early life stages in the upper Missouri River basin, Montana and North Dakota. J. Appl. Ichthyol. 2012, 28, 496–504. [Google Scholar] [CrossRef]
- Holley, C.; Braaten, P.; Poulton, B.; Heist, E.; Umland, L.; Haddix, T. Diet composition and overlap of larval pallid sturgeon and shovelnose sturgeon from the upper Missouri River, USA. Endanger. Species Res. 2022, 49, 103–114. [Google Scholar] [CrossRef]
- Wong, S.; Waldrop, T.; Summerfelt, S.; Davidson, J.; Barrows, F.; Kenney, P.B.; Welch, T.; Wiens, G.D.; Snekvik, K.; Rawls, J.F.; et al. Aquacultured Rainbow Trout (Oncorhynchus mykiss) possess a large core intestinal microbiota that is resistant to variation in diet and rearing density. Appl. Environ. Microbiol. 2013, 79, 4974–4984. [Google Scholar] [CrossRef] [Green Version]
- Neu, A.T.; Allen, E.E.; Roy, K. Defining and quantifying the core microbiome: Challenges and prospects. Proc. Natl. Acad. Sci. 2021, 118, e2104429118. [Google Scholar] [CrossRef]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, P.; Magne, F.; Mardones, P.; Riveros, M.; Opazo, R.; Suau, A.; Pochart, P.; Romero, J. Molecular analysis of intestinal microbiota of rainbow trout (Oncorhynchus mykiss). FEMS Microbiol Ecol. 2010, 71, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Hovda, M.B.; Lunestad, B.T.; Fontanillas, R.; Rosnes, J.T. Molecular characterization of the intestinal microbiota of farmed Atlantic salmon (Salmo salar L.). Aquaculture 2007, 272, 581–588. [Google Scholar] [CrossRef]
- Wilson, B.; Danilowicz, B.S.; Meijer, W.G. The diversity of bacterial communities associated with Atlantic cod Gadus morhua. Microb. Ecol. 2008, 55, 425–434. [Google Scholar] [CrossRef]
- Geraylou, Z.; Souffreau, C.; Rurangwa, E.; D’Hondt, S.; Callewaert, L.; Courtin, C.M.; Delcour, J.A.; Buyse, J.; Ollevier, F. Effects of arabinoxylan-oligosaccharides (AXOS) on juvenile Siberian sturgeon (Acipenser baerii) performance, immune responses and gastrointestinal microbial community. Fish Shellfish Immunol. 2012, 33, 718–724. [Google Scholar] [CrossRef]
- Ye, L.; Amberg, J.; Chapman, D.; Gaikowski, M.; Liu, W.-T. Fish gut microbiota analysis differentiates physiology and behavior of invasive Asian carp and indigenous American fish. ISME J. 2013, 8, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulski, T.; Kozłowski, K.; Ciesielski, S. Habitat and seasonality shape the structure of tench (Tinca tinca L.) gut microbiome. Sci. Rep. 2020, 10, 4460. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Glenn, T.C.; Pierson, T.W.; Bayona-Vásquez, N.J.; Kieran, T.J.; Hoffberg, S.L.; Thomas, J.C.; Lefever, D.E.; Finger, J.W.; Gao, B.; Bian, X.; et al. Adapterama II: Universal amplicon sequencing on Illumina platforms (TaggiMatrix). PeerJ 2019. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. GENEIOUS Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Timmerman, H.M.; Rutten, N.; Boekhorst, J.; Saulnier, D.M.; Kortman, G.; Contractor, N.; Kullen, M. Intestinal colonisation patterns in breastfed and formula-fed infants during the first 12 weeks of life reveal sequential microbiota signatures. Sci. Rep. 2017, 7, 8327. [Google Scholar] [CrossRef] [PubMed]
- Deering, K.E.; Devine, A.; O’Sullivan, T.A.; Lo, J.; Boyce, M.C.; Christophersen, C.T. Characterizing the composition of the pediatric gut microbiome: A systematic review. Nutrients 2020, 12, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Diserud, O.H.; Odegaard, F. A multiple-site similarity measure. Biol. Lett. 2007, 3, 20–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baselga, A. Partitioning abundance-based multiple-site dissimilarity into components: Balanced variation in abundance and abundance gradients. Methods Ecol. Evol. 2017, 8, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.M.; Kubal, M.; Paczan, T.; Rodriguez, A.; Stevens, R.; Wolke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Env. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Chai, B.; Marsh, T.L.; Farris, R.J.; Wang, Q.; Kulam, S.A.; Chandra, S.; McGarrell, D.M.; Schmidt, T.M.; Garrity, G.M.; et al. Ribosomal Database Project. The Ribosomal Database Project (RDP-II): Previewing a new autoaligner that allows regular updates and the new prokaryotic taxonomy. Nucleic Acids Res. 2003, 31, 442–443. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.R.; Curtis, J.T. An ordination of upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A. Environmental and physiological factors shape the gut microbiota of Atlantic salmon parr (Salmo salar L.). Aquaculture 2017, 467, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.K.; Mazmanian, S.K. Has the microbiota played a critical role in the evolution of the adaptive immune system? Science 2010, 330, 1768–1773. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.; Hamady, M.; Lozupone, C.; Turnbaugh, P.; Ramey, R. Evolution of mammals and their gut microbes. Science 2008, 320, 1647. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.; Willemsen, D.; Popkes, M.; Metge, F.; Gandiwa, E.; Reichard, M.; Valenzano, D.R. Regulation of life span by the gut microbiota in the short-lived African turquoise killifish. eLife 2017, 6, e27014. [Google Scholar] [CrossRef]
- Rawls, J.F.; Samuel, B.S.; Gordon, J.I. Gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota. Proc. Natl. Acad. Sci. USA. 2004, 101, 4596–4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawls, J.F.; Mahowald, M.A.; Ley, R.E.; Gordon, J.I. Reciprocal gut microbiota transplants from zebrafish and mice to germ-free recipients reveal host habitat selection. Cell 2006, 127, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, J.M.; Mittge, E.; Kuhlman, J.; Baden, K.N.; Cheesman, S.E.; Guillemin, K. Distinct signals from the microbiota promote different aspects of zebrafish gut differentiation. Dev. Biol. 2006, 297, 374–386. [Google Scholar] [CrossRef]
- Brugman, S.; Liu, K.Y.; Lindenbergh-Kortleve, D.; Samsom, J.N.; Furuta, G.T.; Renshaw, S.A.; Willemsen, R.; Nieuwenhuis, E.E. Oxazolone-induced enterocolitis in zebrafish depends on the composition of the intestinal microbiota. Gastroenterology 2009, 137, 1757–1767.e1. [Google Scholar] [CrossRef]
- Redford, K.H.; Segre, J.A.; Salafsky, N.; del Rio, C.M.; McAloose, D. Conservation and the microbiome. Conserv. Biol. 2012, 26, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.; Jeffery, I.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusak, S.; Harris, H.M.B.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Givens, C.E.; Ransom, B.; Bano, N.; Hollibaugh, J.T. Comparison of the gut microbiomes of 12 bony fish and 3 shark species. Mar. Ecol. Prog. Ser. 2015, 518, 209–223. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ni, J.; Li, J.; Wang, C.; Li, X.; Wu, S.; Zhang, T.; Yu, Y.; Yan, Q. Comparative study on gastrointestinal microbiota of eight fish species with different feeding habits. J. Appl. Microbiol. 2014, 117, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Riordan, T. Human infection with Fusobacterium necrophorum (Necrobacillosis), with a focus on Lemierre’s syndrome. Clin. Microbiol. Rev. 2007, 20, 622–659. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, M.; Kato, S.; Mawatari, H.; Kirikoshi, H.; Imajo, K.; Fujita, K.; Endo, H.; Takahashi, H.; Inamori, M.; Kobayashi, N.; et al. Liver abscess caused by periodontal bacterial infection with Fusobacterium necrophorum. Hepatol. Res. 2011, 41, 194–196. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Aliyu, S.H.; Marriott, R.K.; Curran, M.D.; Parmar, S.; Bentley, N.; Brown, N.M.; Brazier, J.S.; Ludlam, H. Real-time PCR investigation into the importance of Fusobacterium necrophorum as a cause of acute pharyngitis in general practice. J. Med. Microbiol. 2004, 53, 1029–1035. [Google Scholar] [CrossRef]
- Duncan, S.H.; Hold, G.L.; Harmsen, H.J.M.; Stewart, C.S.; Flint, H.J. Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2141–2146. [Google Scholar] [CrossRef] [Green Version]
- Larsen, A.M.; Mohammed, H.H.; Arias, C.R. Characterization of the gut microbiota of three commercially valuable warmwater fish species. J. Appl. Microbiol. 2014, 116, 1396–1404. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Adams, C. Fish pathogens. In The Genus Aeromonas; Austin, B., Altwegg, M., Gosling, P.J., Joseph, S., Eds.; John Wiley & Sons Ltd.: West Sussex, UK, 1996; pp. 197–243. [Google Scholar]
- Joseph, S.W.; Carnahan, A. The isolation, identification, and systematics of the motile Aeromonas species. Annu. Rev. Fish Dis. 1994, 4, 315–343. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.K.; Ghosh, K.; Ringø, E. Enzyme-producing bacteria isolated from fish gut: A review. Aquac. Nutr. 2012, 18, 465–492. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The Gut Microbiota of Marine Fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef]
- Cámara-Ruiz, M.; Balebona, M.C.; Moriñigo, M.; Esteban, M.Á. Probiotic Shewanella putrefaciens (SpPdp11) as a Fish Health Modulator: A Review. Microorganisms 2020, 8, 1990. [Google Scholar] [CrossRef] [PubMed]
- Crab, R.; Defoirdt, T.; Bossier, P.; Verstraete, W. Biofloc technology in aquaculture: Beneficial effects and future challenges. Aquaculture 2012, 351–356. [Google Scholar] [CrossRef]
- Encarnação, P. Functional feed additives in aquaculture feeds. Aquafeed Formul. 2016, 217–237. [Google Scholar] [CrossRef]
- Guerreiro, I.; Oliva-Teles, A.; Enes, P. Prebiotics as functional ingredients: Focus on Mediterranean fish aquaculture. Rev. Aquac. 2018, 10, 800–832. [Google Scholar] [CrossRef]
- D’Silva, A.; Kyndt, J.A. Bacterial Diversity Greatly Affects Ammonia and Overall Nitrogen Levels in Aquabioponics Bioflocs Systems, Based on 16S rRNA Gene Amplicon Metagenomics. Appl. Microbiol. Open Access 2020, 6, 169. [Google Scholar] [CrossRef]
- Xu, Z.; Knight, R. Dietary effects on human gut microbiome diversity. Br. J. Nutr. 2014, 113, S1–S5. [Google Scholar] [CrossRef] [PubMed]
- Gerrity, P.C.; Guy, C.S.; Gardner, W.M. Juvenile Pallid Sturgeon are piscivorous: A call for conserving native cyprinids. Trans. Am. Fish. Soc. 2006, 135, 604–609. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who is eating what: Diet assessment using next generation sequencing. Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Gordon, J.I. Honor thy symbionts. Proc. Natl. Acad. Sci. USA 2003, 100, 10452–10459. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Sex | Capture Location | Date Collected | Tag Number | Year Class | Yrs Since Released from Hatchery to 2018 |
|---|---|---|---|---|---|---|
| Hatchery | Female | Calumet-Bartlett Bend: River km 937.8–931.5 | 2018/4/12 | 4627152F1A | 2002 | 15 |
| 2018/4/13 | 470A643317 | 2005 | 12 | |||
| Lower Plattsmouth Bend: River km 952.2–947.9 | 2018/4/4 | 4A466B2E78 | 2007 | 10 | ||
| 2018/4/5 | 434771011A | 2008 | 9 | |||
| 2018/4/6 | 4900607F63 | 2007 | 10 | |||
| 2018/4/7 | 434A03184E | 2002 | 15 | |||
| Rock Bluff Bend: River km 943.1–937.8 | 2018/4/5 | 434A5C7340 | 2009 | 8 | ||
| 2018/4/10 | 4627056D3B | 2004 | 13 | |||
| 2018/4/10 | 4627273324 | 2002 | 15 | |||
| 2018/4/13 | 471269730D | 2002 | 15 | |||
| Tobacco Bend: River km 947.9–943.1 | 2018/4/5 | 43693D7261 | 2006 | 11 | ||
| 2018/4/6 | 4627313872 | 2003 | 14 | |||
| 2018/4/11 | 47191F2B24 | 2006 | 11 | |||
| Upper Plattsmouth Bend: River km 956–952.2 | 2018/4/8 | 435F151E79 | 2002 | 15 | ||
| 2018/4/8 | 43615C157E | 2002 | 15 | |||
| 2018/4/8 | 4367560D5D | 2002 | 15 | |||
| 2018/4/8 | 4369627915 | 2002 | 15 | |||
| 2018/4/13 | 471E0C4B0E | 2002 | 15 | |||
| Van Horns Bend: River km 927.6–924.7 | 2018/4/10 | 44451B466D | 2002 | 15 | ||
| 2018/4/10 | 46264C5368 | 2003 | 14 | |||
| 2018/4/10 | 471979463C | 2007 | 10 | |||
| 2018/4/10 | 487F075D74 | 2009 | 8 | |||
| Hatchery | Male | Calumet-Bartlett Bend: River km 937.8–931.5 | 2018/4/10 | 46267F6129 | 2006 | 11 |
| 2018/4/12 | 4627702A4D | 2009 | 8 | |||
| Upper Plattsmouth Bend: River km 956–952.2 | 2018/4/6 | 47161C0357 | 2002 | 15 | ||
| 2018/4/6 | 47191A7D15 | 2008 | 10 | |||
| 2018/4/6 | 847F623E77 | 2007 | 10 | |||
| 2018/4/8 | 4A467F4F41 | 2007 | 10 | |||
| Rock Bluff Bend: River km 943.1–937.8 | 2018/4/5 | 434A41496C | 2010 | 7 | ||
| 2018/4/5 | 46280A267E | 2005 | 12 | |||
| 2018/4/10 | 46267A6226 | 2009 | 8 | |||
| Tobacco Bend: River km 947.9–943.1 | 2018/4/6 | 4349450B3E | 2004 | 13 | ||
| Upper Plattsmouth Bend: River km 956–952.2 | 2018/4/7 | 434A4F216C | 2008 | 9 | ||
| Hatchery | Unknown | Tobacco Bend: River km 947.9–43.1 | 2018/4/6 | 4257361E77 | 2002 | 15 |
| Unknown | Unknown | Lower Plattsmouth Bend: River km 952.2–947.9 | 2018/4/7 | 434A6B1F62 | ||
| Upper Plattsmouth Bend: River km 956–952.2 | 2018/4/7 | 46256D3718 | ||||
| Unknown | Male | Rock Bluff Bend: River km 943.1–937.8 | 2018/4/10 | 434969543E | ||
| Wild | Female | Lower Plattsmouth Bend: River km 952.2–947.9 | 2018/4/4 | 4B191E7809 | ||
| Upper Plattsmouth Bend: River km 956–952.2 | 2018/4/8 | 4627683B2B | ||||
| Calumet-Bartlett Bend: River km 937.8–931.5 | 2018/4/12 | 47134D0F2A | ||||
| Wild | Male | Calumet-Bartlett Bend: River km 937.8–931.5 | 2018/4/12 | 470B110F50 | ||
| Lower Plattsmouth Bend: River km 952.2–947.9 | 2018/4/7 | 462622242B | ||||
| Lower Plattsmouth Bend: River km 952.2–947.9 | 2018/4/6 | 43449D6C1E | ||||
| Tobacco Bend: River km 947.9–943.1 | 2018/4/4 | 4704510611 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaughan, S.; Kyndt, J.A.; Haas, J.D.; Steffensen, K.D.; Kočovský, P.M.; Pope, K.L. Using the Gut Microbiome to Assess Stocking Efforts of the Endangered Pallid Sturgeon, Scaphirhynchus albus. Life 2023, 13, 309. https://doi.org/10.3390/life13020309
Gaughan S, Kyndt JA, Haas JD, Steffensen KD, Kočovský PM, Pope KL. Using the Gut Microbiome to Assess Stocking Efforts of the Endangered Pallid Sturgeon, Scaphirhynchus albus. Life. 2023; 13(2):309. https://doi.org/10.3390/life13020309
Chicago/Turabian StyleGaughan, Sarah, John A. Kyndt, Justin D. Haas, Kirk D. Steffensen, Patrick M. Kočovský, and Kevin L. Pope. 2023. "Using the Gut Microbiome to Assess Stocking Efforts of the Endangered Pallid Sturgeon, Scaphirhynchus albus" Life 13, no. 2: 309. https://doi.org/10.3390/life13020309