
Detection of Circulating SARS-CoV-2 Variants of Concern (VOCs) Using a Multiallelic Spectral Genotyping Assay

 ,
,
Abstract
:1. Introduction


2. Materials and Methods
2.1. Molecular Beacon and Primer Design
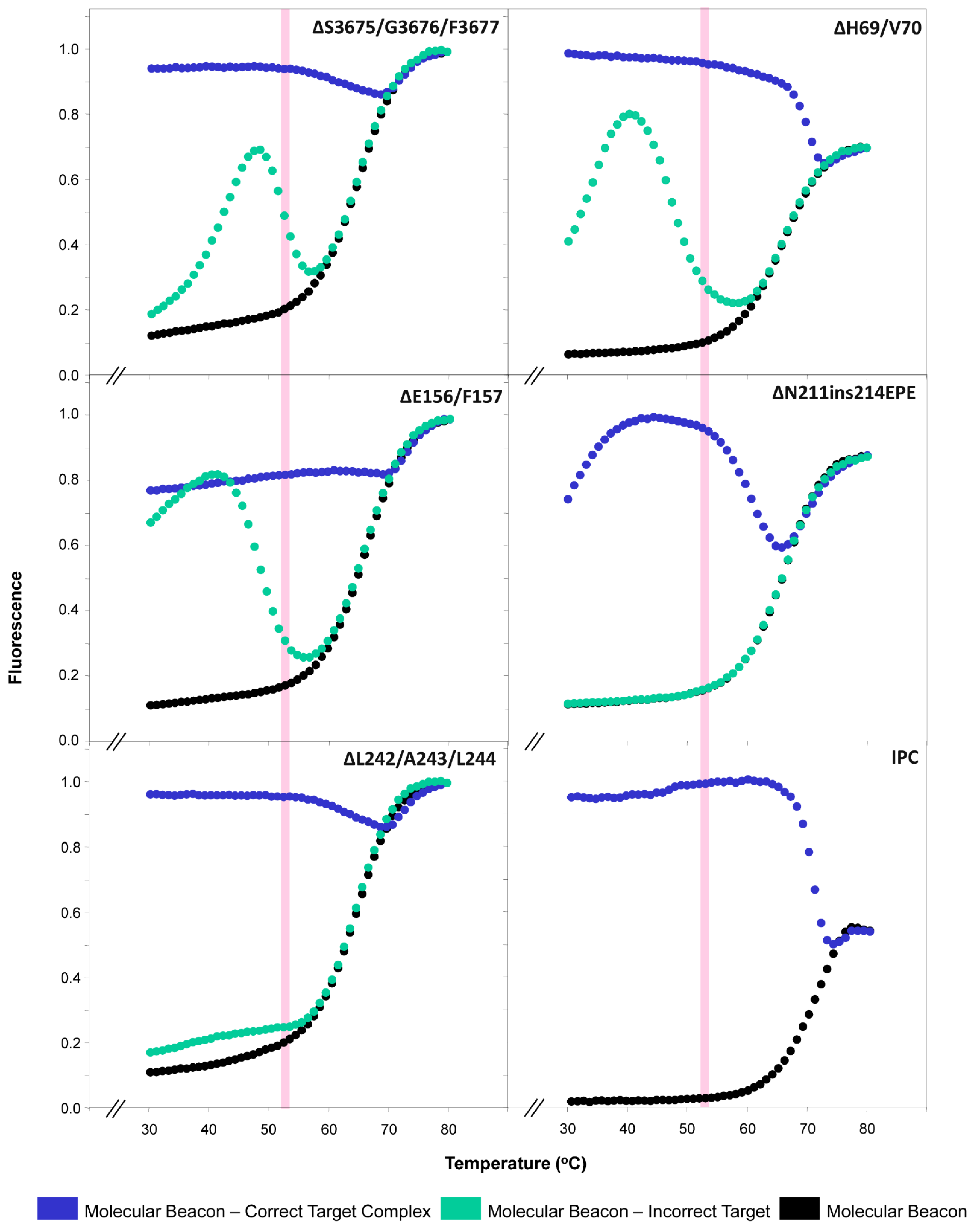
2.2. Thermal Profiles of Molecular Beacons
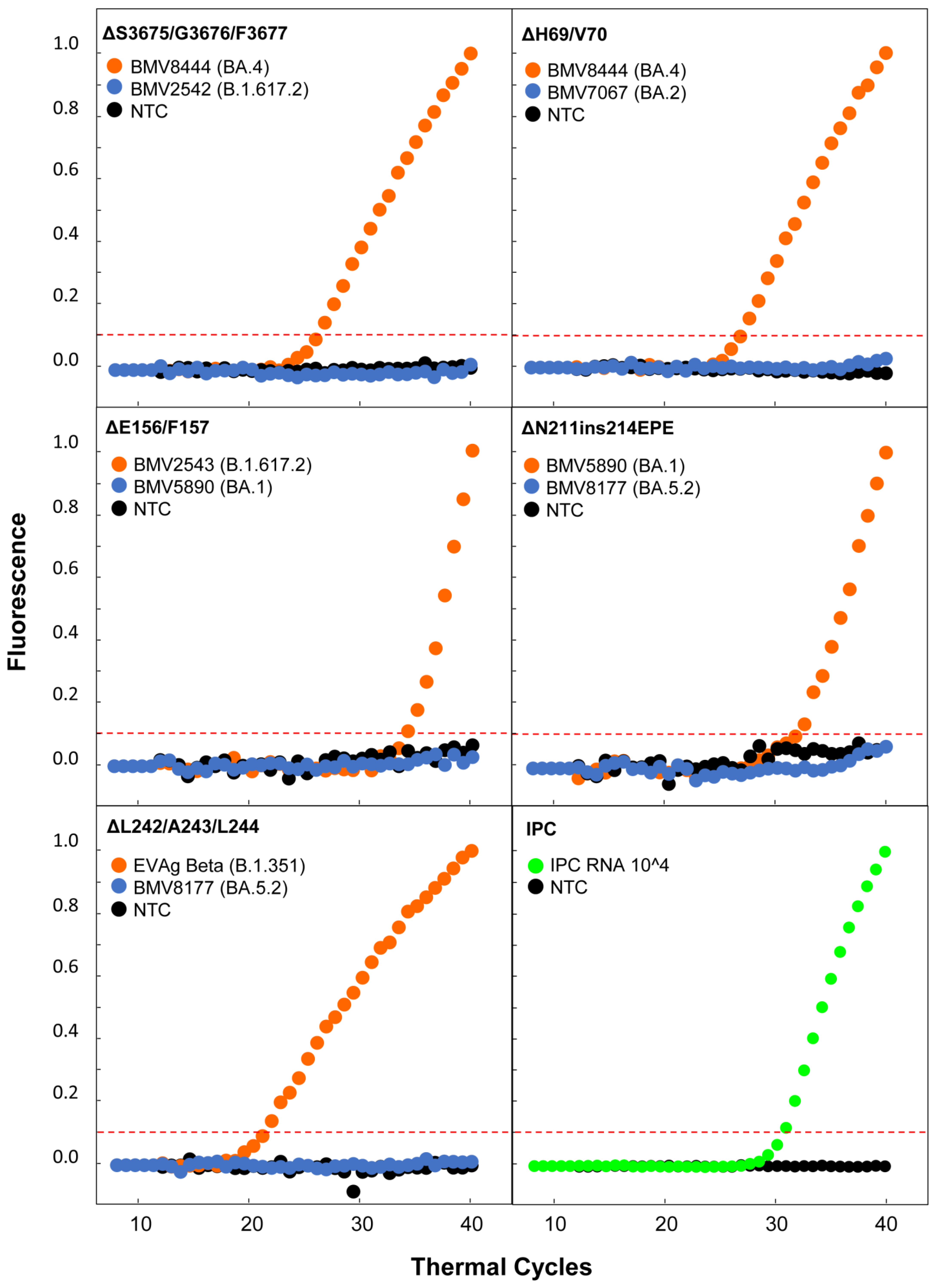
2.3. Real-Time RT-PCR
2.4. Reference and Clinical Samples Used to Test the Assay
3. Results
3.1. Molecular Beacon Thermal Denaturation Profiles
3.2. Real-Time RT-PCR Testing Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, J. SARS-CoV-2: An Emerging Coronavirus that Causes a Global Threat. Int. J. Biol. Sci. 2020, 16, 1678–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demoliner, M.; Gularte, J.S.; Girardi, V.; Almeida, P.R.; de Weber, M.N.; Eisen, A.K.A.; Fleck, J.D.; Spilki, F.R. SARS-CoV-2 and COVID-19: A perspective from environmental virology. Genet. Mol. Biol. 2021, 44. [Google Scholar] [CrossRef] [PubMed]
- CSSE JHU, C. for S.S. and E. (CSSE) at J.H.U. (JHU) COVID-19 Dashboard. Available online: https://gisanddata.maps.arcgis.com/apps/dashboards/bda7594740fd40299423467b48e9ecf6 (accessed on 14 December 2022).
- Dong, E.; Du, H.; Gardner, L. An interactive web-based dashboard to track COVID-19 in real time. Lancet Infect. Dis. 2020, 20, 533–534. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Apostolopoulos, V. COVID-19 vaccine design and vaccination strategy for emerging variants. Expert Rev. Vaccines 2022, 21, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- DeGrace, M.M.; Ghedin, E.; Frieman, M.B.; Krammer, F.; Grifoni, A.; Alisoltani, A.; Alter, G.; Amara, R.R.; Baric, R.S.; Barouch, D.H.; et al. Defining the risk of SARS-CoV-2 variants on immune protection. Nature 2022, 605, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Hanuma Kumar Ghali, E.N.; Yallapu, M.M.; Apostolopoulos, V. Therapeutics to tackle Omicron outbreak. Immunotherapy 2022, 14, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Patel, A.B.; Vaghasiya, D.D. SARS-CoV-2 variants and vulnerability at the global level. J. Med. Virol. 2022, 94, 2986–3005. [Google Scholar] [CrossRef]
- Chen, J.; Wang, R.; Wei, G.-W. Review of the mechanisms of SARS-CoV-2 evolution and transmission. arXiv 2021, arXiv:2109.08148. [Google Scholar]
- Singh, J.; Pandit, P.; McArthur, A.G.; Banerjee, A.; Mossman, K. Evolutionary trajectory of SARS-CoV-2 and emerging variants. Virol. J. 2021, 18, 166. [Google Scholar] [CrossRef]
- Tang, X.; Ying, R.; Yao, X.; Li, G.; Wu, C.; Tang, Y.; Li, Z.; Kuang, B.; Wu, F.; Chi, C.; et al. Evolutionary analysis and lineage designation of SARS-CoV-2 genomes. Sci. Bull. 2021, 66, 2297–2311. [Google Scholar] [CrossRef]
- Rambaut, A.; Holmes, E.C.; O’Toole, Á.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A dynamic nomenclature proposal for SARS-CoV-2 lineages to assist genomic epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organization (WHO). Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (accessed on 22 November 2022).
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; de Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The biological and clinical significance of emerging SARS-CoV-2 variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef] [PubMed]
- Flores-Vega, V.R.; Monroy-Molina, J.V.; Jiménez-Hernández, L.E.; Torres, A.G.; Santos-Preciado, J.I.; Rosales-Reyes, R. SARS-CoV-2: Evolution and Emergence of New Viral Variants. Viruses 2022, 14, 653. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Smith, D.M. SARS-CoV-2 variants of concern. Yonsei Med. J. 2021, 62, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, R.K.; Kuppili, S.; Kumar Suvvari, T.; Kandi, V.; Behera, A.; Verma, S.; Kudrat-E-Zahan; Biswal, S.K.; Al-Noor, T.H.; El-ajaily, M.M.; et al. SARS-CoV-2 and its variants of concern including Omicron: A never ending pandemic. Chem. Biol. Drug Des. 2022, 99, 769–788. [Google Scholar] [CrossRef]
- Migueres, M.; Lhomme, S.; Trémeaux, P.; Dimeglio, C.; Ranger, N.; Latour, J.; Dubois, M.; Nicot, F.; Miedouge, M.; Mansuy, J.M.; et al. Evaluation of two RT-PCR screening assays for identifying SARS-CoV-2 variants. J. Clin. Virol. 2021, 143, 104969. [Google Scholar] [CrossRef] [PubMed]
- Vega-Magaña, N.; Sánchez-Sánchez, R.; Hernández-Bello, J.; Venancio-Landeros, A.A.; Peña-Rodríguez, M.; Vega-Zepeda, R.A.; Galindo-Ornelas, B.; Díaz-Sánchez, M.; García-Chagollán, M.; Macedo-Ojeda, G.; et al. RT-qPCR Assays for Rapid Detection of the N501Y, 69-70del, K417N, and E484K SARS-CoV-2 Mutations: A Screening Strategy to Identify Variants With Clinical Impact. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Ting, Y.; Ye, X.; Rongrong, Z.; Xiaohong, Z.; Zehui, C.; Su, L.; Zihan, X.; Yiqun, L.; Yongyou, Z.; Qingge, L. Accessible and Adaptable Multiplexed Real-Time PCR Approaches to Identify SARS-CoV-2 Variants of Concern. Microbiol. Spectr. 2022, 10, e03222-22. [Google Scholar] [CrossRef]
- Bull, R.A.; Adikari, T.N.; Ferguson, J.M.; Hammond, J.M.; Stevanovski, I.; Beukers, A.G.; Naing, Z.; Yeang, M.; Verich, A.; Gamaarachchi, H.; et al. Analytical validity of nanopore sequencing for rapid SARS-CoV-2 genome analysis. Nat. Commun. 2020, 11, 6272. [Google Scholar] [CrossRef]
- John, G.; Sahajpal, N.S.; Mondal, A.K.; Ananth, S.; Williams, C.; Chaubey, A.; Rojiani, A.M.; Kolhe, R. Next-Generation Sequencing (NGS) in COVID-19: A Tool for SARS-CoV-2 Diagnosis, Monitoring New Strains and Phylodynamic Modeling in Molecular Epidemiology. Curr. Issues Mol. Biol. 2021, 43, 845–867. [Google Scholar] [CrossRef]
- Tan, W.; Wang, K.; Drake, T.J. Molecular beacons. Curr. Opin. Chem. Biol. 2004, 8, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Kramer, F.R. Molecular Beacons: Probes that Fluoresce upon Hybridization. Nat. Biotechnol. 1996, 14, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Bratu, D.P.; Kramer, F.R. Multicolor molecular beacons for allele discrimination. Nat. Biotechnol. 1998, 16, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Kostrikis, L.G.; Tyagi, S.; Mhlanga, M.M.; Ho, D.D.; Kramer, F.R. Spectral genotyping of human alleles. Science 1998, 279, 1228–1229. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.W.; Shin, A.; Park, S.S.; Lee, J.-M. Haplotype-specific insertion-deletion variations for allele-specific targeting in Huntington’s disease. Mol. Ther.-Methods Clin. Dev. 2022, 25, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Sherrill-Mix, S.; Van Duyne, G.D.; Bushman, F.D. Molecular Beacons Allow Specific RT-LAMP Detection of B.1.1.7 Variant SARS-CoV-2. J. Biomol. Tech. 2021, 32, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Sharma, K.; Singh, P.; Bhargava, A.; Negi, S.S.; Sharma, P.; Bhise, M.; Tripathi, M.K.; Jindal, A.; Nagarkar, N.M. Genomic characterization unravelling the causative role of SARS-CoV-2 Delta variant of lineage B.1.617.2 in 2nd wave of COVID-19 pandemic in Chhattisgarh, India. Microb. Pathog. 2022, 164, 105404. [Google Scholar] [CrossRef]
- McCallum, M.; Walls, A.C.; Sprouse, K.R.; Bowen, J.E.; Rosen, L.E.; Dang, H.V.; De Marco, A.; Franko, N.; Tilles, S.W.; Logue, J.; et al. Molecular basis of immune evasion by the Delta and Kappa SARS-CoV-2 variants. Science 2021, 374, 1621–1626. [Google Scholar] [CrossRef]
- Harari, S.; Tahor, M.; Rutsinsky, N.; Meijer, S.; Miller, D.; Henig, O.; Halutz, O.; Levytskyi, K.; Ben-Ami, R.; Adler, A.; et al. Drivers of adaptive evolution during chronic SARS-CoV-2 infections. Nat. Med. 2022, 28, 1501–1508. [Google Scholar] [CrossRef]
- Zhou, D.; Dejnirattisai, W.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.E.; Tuekprakhon, A.; Nutalai, R.; et al. Evidence of escape of SARS-CoV-2 variant B.1.351 from natural and vaccine-induced sera. Cell 2021, 184, 2348–2361.e6. [Google Scholar] [CrossRef]
- Pastorio, C.; Zech, F.; Noettger, S.; Jung, C.; Jacob, T.; Sanderson, T.; Sparrer, K.M.J.; Kirchhoff, F. Determinants of Spike infectivity, processing, and neutralization in SARS-CoV-2 Omicron subvariants BA.1 and BA.2. Cell Host Microbe 2022, 30, 1255–1268.e5. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Kemp, S.A.; Papa, G.; Datir, R.; Ferreira, I.A.T.M.; Marelli, S.; Harvey, W.T.; Lytras, S.; Mohamed, A.; Gallo, G.; et al. Recurrent emergence of SARS-CoV-2 spike deletion H69/V70 and its role in the Alpha variant B.1.1.7. Cell Rep. 2021, 35, 109292. [Google Scholar] [CrossRef] [PubMed]
- Kistler, K.E.; Huddleston, J.; Bedford, T. Rapid and parallel adaptive mutations in spike S1 drive clade success in SARS-CoV-2. Cell Host Microbe 2022, 30, 545–555.e4. [Google Scholar] [CrossRef] [PubMed]
- Chrysostomou, A.C.; Hezka Rodosthenous, J.; Topcu, C.; Papa, C.; Aristokleous, A.; Stathi, G.; Christodoulou, C.; Eleftheriou, C.; Stylianou, D.C.; Kostrikis, L.G. A Multiallelic Molecular Beacon-Based Real-Time RT-PCR Assay for the Detection of SARS-CoV-2. Life 2021, 11, 1146. [Google Scholar] [CrossRef] [PubMed]
- Hadjinicolaou, A.V.; Farcas, G.A.; Demetriou, V.L.; Mazzulli, T.; Poutanen, S.M.; Willey, B.M.; Low, D.E.; Butany, J.; Asa, S.L.; Kain, K.C.; et al. Development of a molecular-beacon-based multi-allelic real-time RT-PCR assay for the detection of human coronavirus causing severe acute respiratory syndrome (SARS-CoV): A general methodology for detecting rapidly mutating viruses. Arch. Virol. 2011, 156, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Dikdan, R.J.; Marras, S.A.E.; Field, A.P.; Brownlee, A.; Cironi, A.; Hill, D.A.; Tyagi, S. Multiplex PCR Assays for Identifying all Major Severe Acute Respiratory Syndrome Coronavirus 2 Variants. J. Mol. Diagnostics 2022, 24, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Manjunath, R.; Gaonkar, S.L.; Saleh, E.A.M.; Husain, K. A comprehensive review on Covid-19 Omicron (B.1.1.529) variant. Saudi J. Biol. Sci. 2022, 29, 103372. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). SARS-CoV-2 Variants of Concern as of 1 December 2022. Available online: https://www.ecdc.europa.eu/en/covid-19/variants-concern (accessed on 1 December 2022).
- Bateman, A.; Martin, M.J.; O’Donovan, C.; Magrane, M.; Alpi, E.; Antunes, R.; Bely, B.; Bingley, M.; Bonilla, C.; Britto, R.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.W.; Sahi, V.; Figueroa, A.; et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef]
- Ma, J.; Acevedo, A.C.; Wang, Q. High-Potency Polypeptide-based Interference for Coronavirus Spike Glycoproteins. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zhu, C.; He, G.; Yin, Q.; Zeng, L.; Ye, X.; Shi, Y.; Xu, W. Molecular biology of the SARs-CoV-2 spike protein: A review of current knowledge. J. Med. Virol. 2021, 93, 5729–5741. [Google Scholar] [CrossRef] [PubMed]
- Khelashvili, G.; Plante, A.; Doktorova, M.; Weinstein, H. Ca(2+)-dependent mechanism of membrane insertion and destabilization by the SARS-CoV-2 fusion peptide. Biophys. J. 2021, 120, 1105–1119. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Nair, M.S.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.D.; et al. Antibody Resistance of SARS-CoV-2 Variants B.1.351 and B.1.1.7. Nature 2021, 593, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, J.H.; Lee, S.; Shim, S.; Nguyen, T.T.; Hwang, J.; Kim, H.; Choi, Y.O.; Hong, J.; Bae, S.; et al. The progression of sars coronavirus 2 (Sars-cov2): Mutation in the receptor binding domain of spike gene. Immune Netw. 2020, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Manjunath, K.; Ranjan, R.K.; Kaushik, S.; Kumar, S.; Verma, V. COVID-19 pandemic: Insights into structure, function, and hACE2 receptor recognition by SARS-CoV-2. PLoS Pathog. 2020, 16, e1008762. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Xia, X. Domains and Functions of Spike Protein in Sars-Cov-2 in the Context of Vaccine Design. Viruses 2021, 13, 109. [Google Scholar] [CrossRef]
- Gobeil, S.M.C.; Janowska, K.; McDowell, S.; Mansouri, K.; Parks, R.; Manne, K.; Stalls, V.; Kopp, M.F.; Henderson, R.; Edwards, R.J.; et al. D614G Mutation Alters SARS-CoV-2 Spike Conformation and Enhances Protease Cleavage at the S1/S2 Junction. Cell Rep. 2021, 34, 108630. [Google Scholar] [CrossRef]
- Sasaki, M.; Uemura, K.; Sato, A.; Toba, S.; Sanaki, T.; Maenaka, K.; Hall, W.W.; Orba, Y.; Sawa, H. SARS-CoV-2 variants with mutations at the S1/ S2 cleavage site are generated in vitro during propagation in TMPRSS2-deficient cells. PLoS Pathog. 2021, 17, e1009233. [Google Scholar] [CrossRef]
- Chen, C.; Nadeau, S.; Yared, M.; Voinov, P.; Xie, N.; Roemer, C.; Stadler, T. CoV-Spectrum: Analysis of globally shared SARS-CoV-2 data to identify and characterize new variants. Bioinformatics 2022, 38, 1735–1737. [Google Scholar] [CrossRef]
- Johnson, R.; Mangwana, N.; Sharma, J.R.; Muller, C.J.F.; Malemela, K.; Mashau, F.; Dias, S.; Ramharack, P.; Kinnear, C.; Glanzmann, B.; et al. Delineating the Spread and Prevalence of SARS-CoV-2 Omicron Sublineages (BA.1–BA.5) and Deltacron Using Wastewater in the Western Cape, South Africa. J. Infect. Dis. 2022, 226, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Hodcroft, E.B. CoVariants: SARS-CoV-2 Mutations and Variants of Interest. Available online: https://covariants.org/ (accessed on 1 December 2022).
- Liu, J.; Liu, Y.; Xia, H.; Zou, J.; Weaver, S.C.; Swanson, K.A.; Cai, H.; Cutler, M.; Cooper, D.; Muik, A.; et al. BNT162b2-elicited neutralization of B.1.617 and other SARS-CoV-2 variants. Nature 2021, 596, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Shiakolas, A.R.; Kramer, K.J.; Johnson, N.V.; Wall, S.C.; Suryadevara, N.; Wrapp, D.; Periasamy, S.; Pilewski, K.A.; Raju, N.; Nargi, R.; et al. Efficient discovery of SARS-CoV-2-neutralizing antibodies via B cell receptor sequencing and ligand blocking. Nat. Biotechnol. 2022, 40, 1270–1275. [Google Scholar] [CrossRef] [PubMed]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Detection of a SARS-CoV-2 variant of concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.; Gurry, C.; Freitas, L.; Schultz, M.B.; Bach, G.; Diallo, A.; Akite, N.; Ho, J.; Lee, R.T.C.; Yeo, W. GISAID’s Role in Pandemic Response. China CDC Wkly. 2021, 3, 1049. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.F.W.; Siu, G.K.H.; Yuan, S.; Ip, J.D.; Cai, J.P.; Chu, A.W.H.; Chan, W.M.; Abdullah, S.M.U.; Luo, C.; Chan, B.P.C.; et al. Probable Animal-to-Human Transmission of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Delta Variant AY.127 Causing a Pet Shop-Related Coronavirus Disease 2019 (COVID-19) Outbreak in Hong Kong. Clin. Infect. Dis. 2022, 75, e76–e81. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 10, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Kuraku, S.; Zmasek, C.M.; Nishimura, O.; Katoh, K. aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity. Nucleic Acids Res. 2013, 41, W22–W28. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Chrysostomou, A.C.; Vrancken, B.; Haralambous, C.; Alexandrou, M.; Aristokleous, A.; Christodoulou, C.; Gregoriou, I.; Ioannides, M.; Kalakouta, O.; Karagiannis, C.; et al. Genomic Epidemiology of the SARS-CoV-2 Epidemic in Cyprus from November 2020 to October 2021: The Passage of Waves of Alpha and Delta Variants of Concern. Viruses 2023, 15, 108. [Google Scholar] [CrossRef]
- Chrysostomou, A.C.; Vrancken, B.; Koumbaris, G.; Themistokleous, G.; Aristokleous, A.; Masia, C.; Eleftheriou, C.; Iοannou, C.; Stylianou, D.C.; Ioannides, M.; et al. A Comprehensive Molecular Epidemiological Analysis of SARS-CoV-2 Infection in Cyprus from April 2020 to January 2021: Evidence of a Highly Polyphyletic and Evolving Epidemic. Viruses 2021, 13, 1098. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of epidemiological lineages in an emerging pandemic using the pangolin tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef] [PubMed]
- Aleem, A.; Akbar Samad, A.B.; Slenker, A.K. Emerging Variants of SARS-CoV-2 And Novel Therapeutics Against Coronavirus (COVID-19); StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Mariana, A.; AliReza, E.; Stephen, J.P.; George, B.; Antoine, C.; Kirby, C.; Nahuel, F.; Jessica, D.F.; Jennifer, L.G.; Julianne, V.K.; et al. Real-Time RT-PCR Allelic Discrimination Assay for Detection of N501Y Mutation in the Spike Protein of SARS-CoV-2 Associated with B.1.1.7 Variant of Concern. Microbiol. Spectr. 2022, 10, e00681-21. [Google Scholar] [CrossRef]
- Fan, Y.; Li, X.; Zhang, L.; Wan, S.; Zhang, L.; Zhou, F. SARS-CoV-2 Omicron variant: Recent progress and future perspectives. Signal Transduct. Target. Ther. 2022, 7, 141. [Google Scholar] [CrossRef] [PubMed]
- Padmapriya, B.; Raquel, G.; Sukalyani, B.; Abby, C.; Deanna, S.; Robert, J.; Soumitesh, C.; David, A. A Simple Reverse Transcriptase PCR Melting-Temperature Assay To Rapidly Screen for Widely Circulating SARS-CoV-2 Variants. J. Clin. Microbiol. 2021, 59, e00845-21. [Google Scholar] [CrossRef]
- Jiang, W.; Ji, W.; Zhang, Y.; Xie, Y.; Chen, S.; Jin, Y.; Duan, G. An Update on Detection Technologies for SARS-CoV-2 Variants of Concern. Viruses 2022, 14, 2324. [Google Scholar] [CrossRef]
- Etievant, S.; Bal, A.; Escuret, V.; Brengel-Pesce, K.; Bouscambert, M.; Cheynet, V.; Generenaz, L.; Oriol, G.; Destras, G.; Billaud, G.; et al. Performance Assessment of SARS-CoV-2 PCR Assays Developed by WHO Referral Laboratories. J. Clin. Med. 2020, 9, 1871. [Google Scholar] [CrossRef]





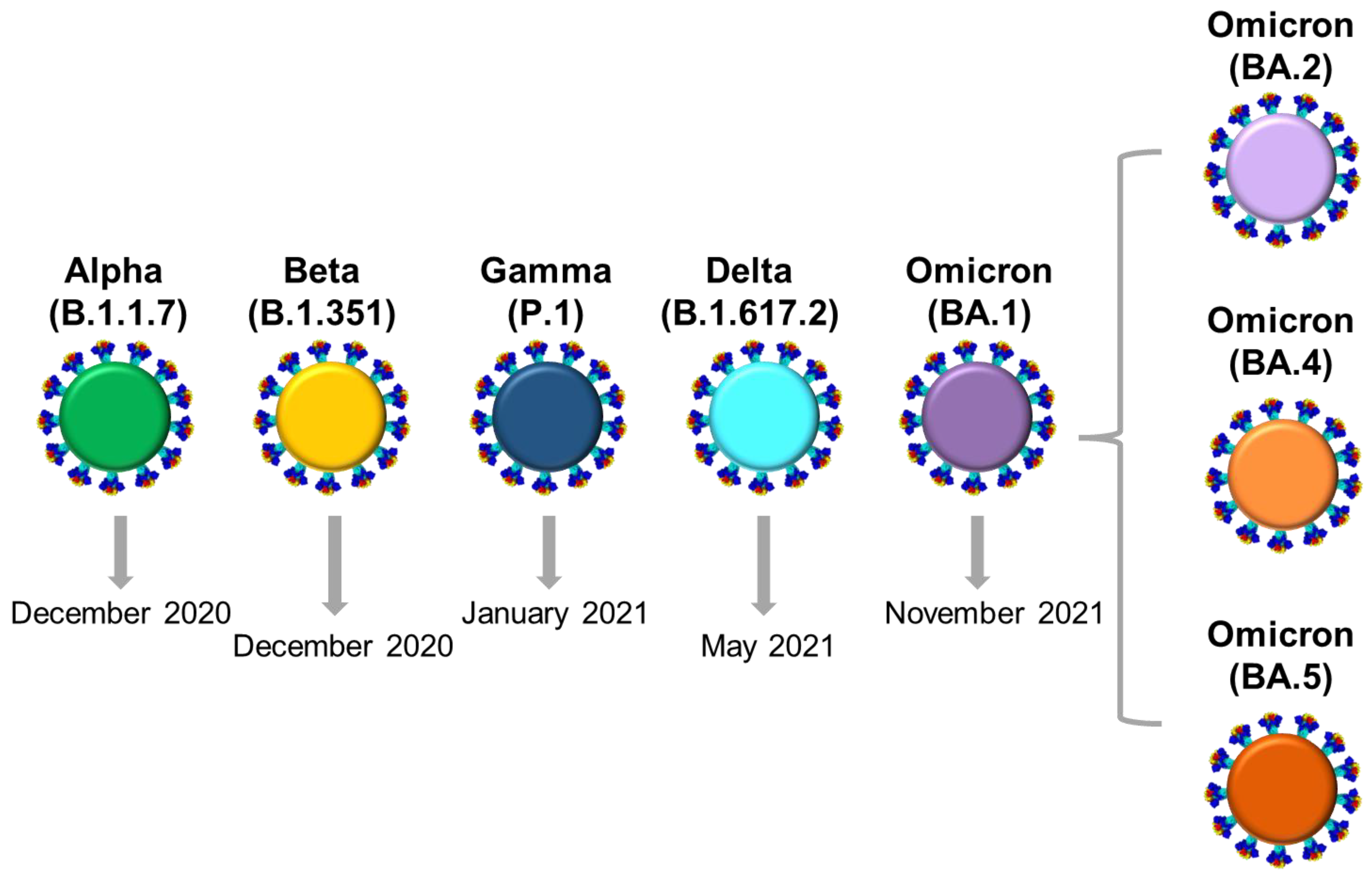{kind=link}
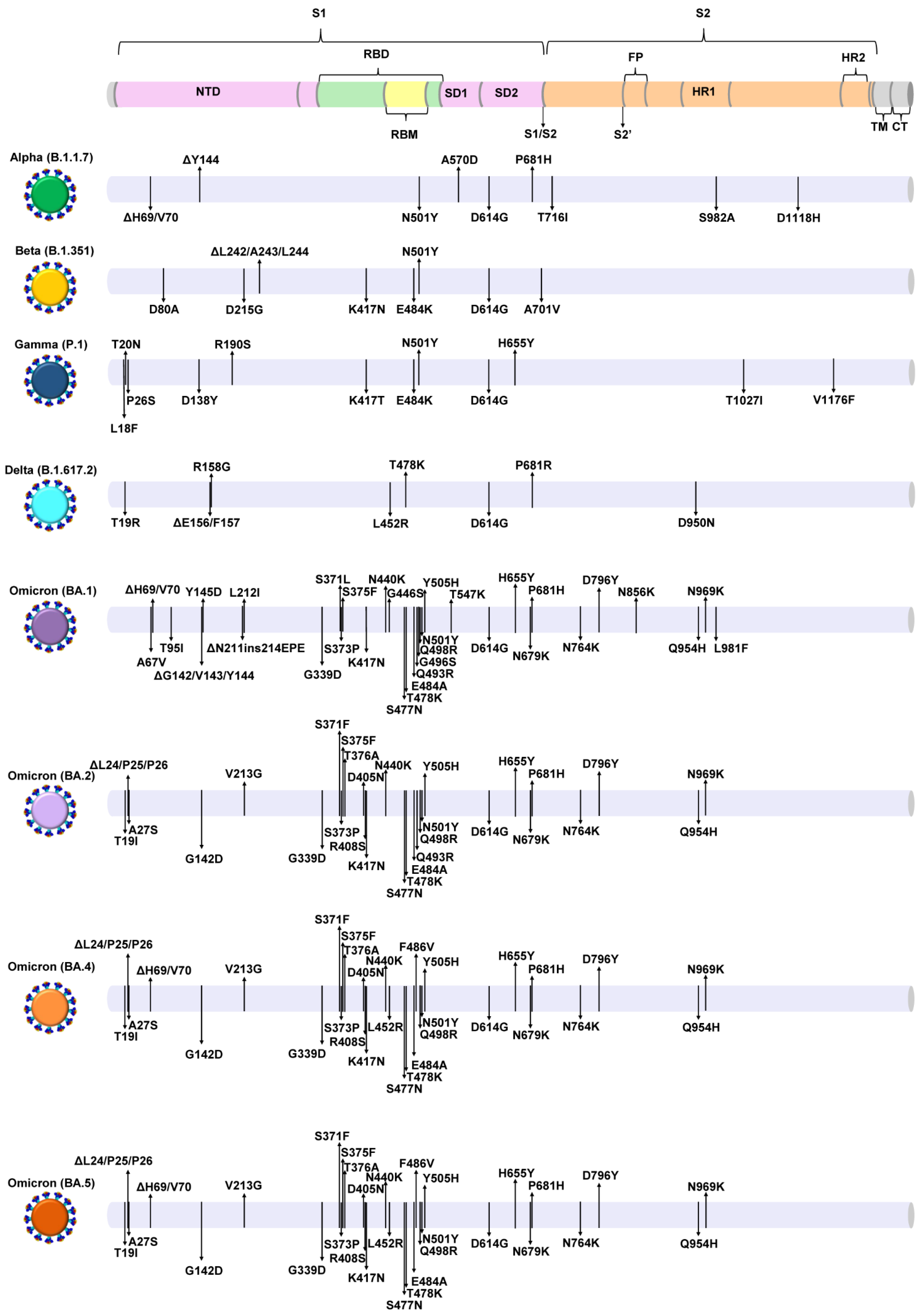{kind=link}
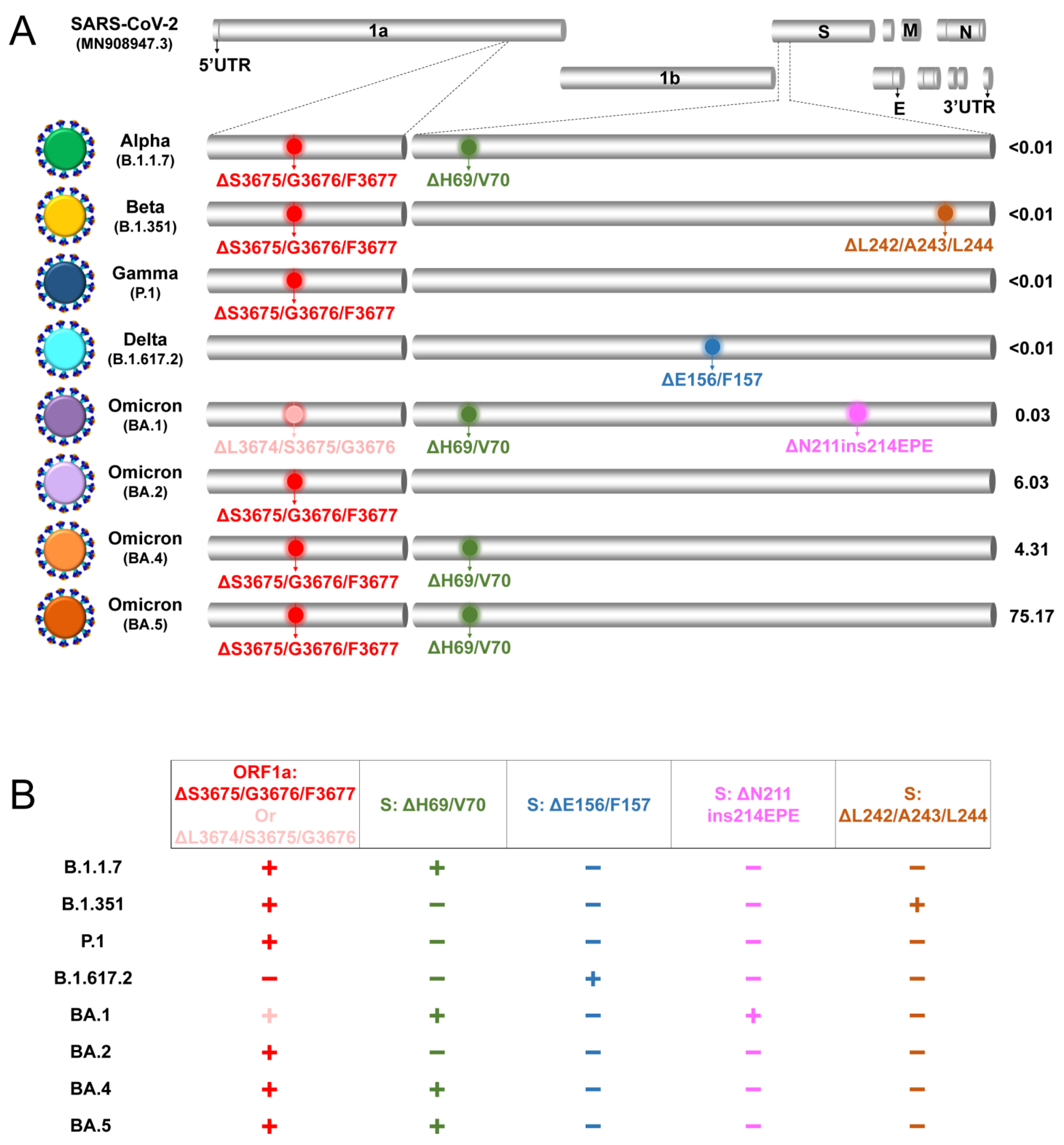{kind=link}
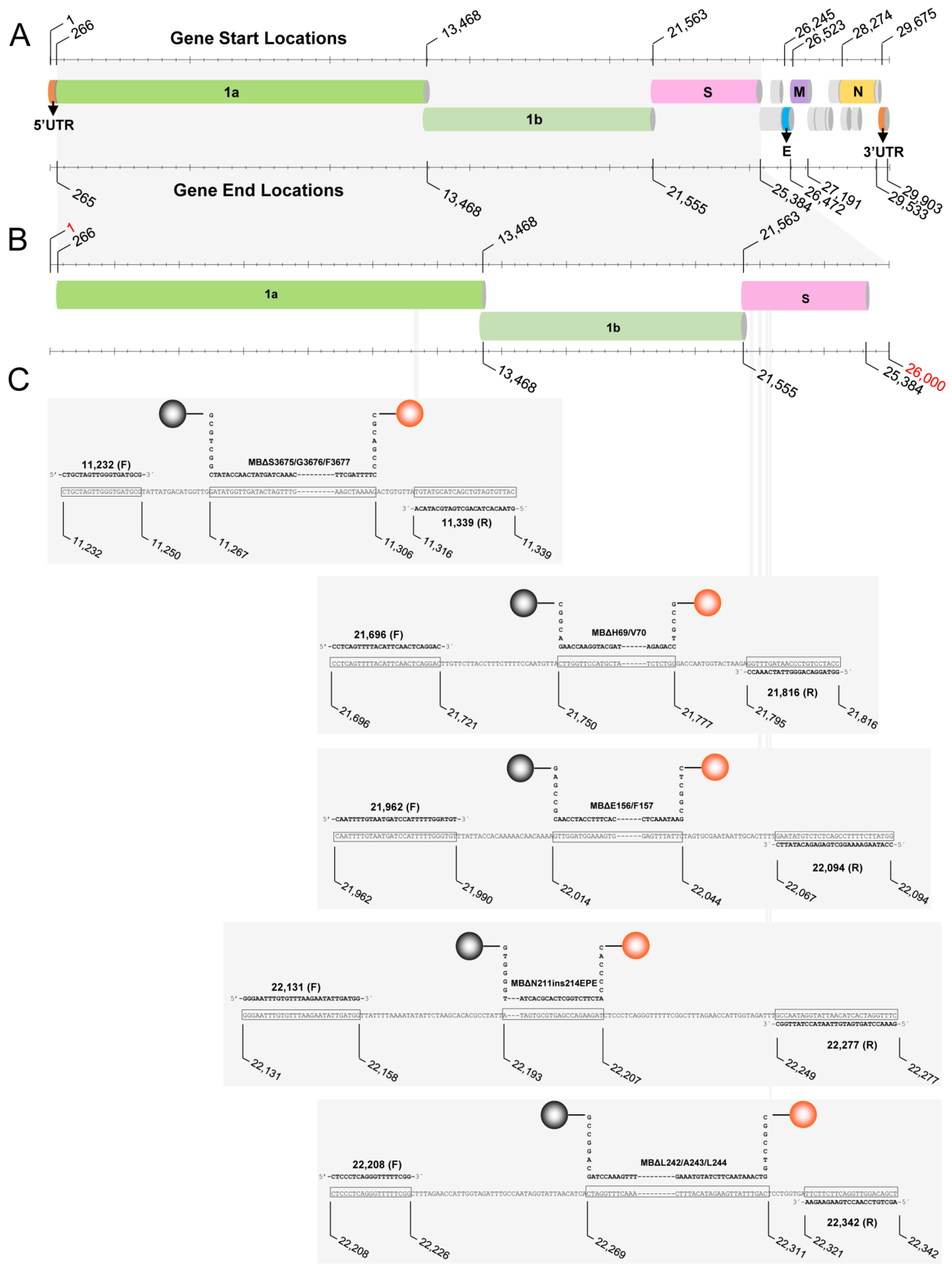{kind=link}
{kind=link}
{kind=link}
| Designation a | Target Gene | Sequence | Position b | Amplicon Length (nts) c | Gene Accession Number d | Reference e |
|---|---|---|---|---|---|---|
| PCR Primers | ||||||
| 11,232 (F) | ORF1a | CTGCTAGTTGGGTGATGCG | 11,232–11,250 | MN908947.3 | This study | |
| 11,339 (R) | ORF1a | GTAACACTACAGCTGATGCATACA | 11,316–11,339 | MN908947.3 | This study | |
| 21,696 (F) | S | CCTCAGTTTTACATTCAACTCAGGAC | 21,696–21,721 | MN908947.3 | This study | |
| 21,816 (R) | S | GGTAGGACAGGGTTATCAAACC | 21,795–21,816 | MN908947.3 | This study | |
| 21,962 (F) | S | CAATTTTGTAATGATCCATTTTTGGRTGT | 21,962–21,990 | MN908947.3 | This study | |
| 22,094 (R) | S | CCATAAGAAAAGGCTGAGAGACATATTC | 22,067–22,094 | MN908947.3 | This study | |
| 22,131 (F) | S | GGGAATTTGTGTTTAAGAATATTGATGG | 22,131–22,158 | MN908947.3 | [36] | |
| 22,208 (F) | S | CTCCCTCAGGGTTTTTCGG | 22,208–22,226 | MN908947.3 | This study | |
| 22,277 (R) | S | GAAACCTAGTGATGTTAATACCTATTGGC | 22,249–22,277 | MN908947.3 | [36] | |
| 22,342 (R) | S | AGCTGTCCAACCTGAAGAAGAA | 22,321–22,342 | MN908947.3 | This study | |
| 26,355 (R) | E | AAGCGCAGTAAGGATGGCTA | 26,336–26,355 | MN908947.3 | [36] | |
| Target Amplicons | ||||||
| TΔS3675/G3676/F3677 | ORF1a | CTGCTAGTTGGGTGATGCGTATTATGACATGGTTGGATATGGTTGATACTAGTTTGAAGCTAAAAGA | 11,232–11,339 | 99 | MN908947.3 | This study |
| CTGTGTTATGTATGCATCAGCTGTAGTGTTAC | ||||||
| TΔH69/V70 | S | CCTCAGTTTTACATTCAACTCAGGACTTGTTCTTACCTTTCTTTTCCAATGTTACTTGGTTCCATGCTA | 21,696–21,816 | 115 | MN908947.3 | This study |
| TCTCTGGGACCAATGGTACTAAGAGGTTTGATAACCCTGTCCTACC | ||||||
| TΔE156/F157 | S | CAATTTTGTAATGATCCATTTTTGGGTGTTTATTACCACAAAAACAACAAAAGTTGGATGGAAAGTG | 21,962–22,094 | 127 | MN908947.3 | This study |
| GAGTTTATTCTAGTGCGAATAATTGCACTTTTGAATATGTCTCTCAGCCTTTTCTTATGG | ||||||
| TΔΝ211ins214EPE | S | GGGAATTTGTGTTTAAGAATATTGATGGTTATTTTAAAATATATTCTAAGCACACGCCTATTATAGTG | 22,131–22,277 | 153 | MN908947.3 | This study |
| CGTGAGCCAGAAGATCTCCCTCAGGGTTTTTCGGCTTTAGAACCATTGGTAGATTTGCCAATAGGTAT | ||||||
| TAACATCACTAGGTTTC | ||||||
| TΔL242/A243/L244 | S | CTCCCTCAGGGTTTTTCGGCTTTAGAACCATTGGTAGATTTGCCAATAGGTATTAACATCACTAGGTT | 22,208–22,342 | 126 | MN908947.3 | This study |
| TCAAACTTTACATAGAAGTTATTTGACTCCTGGTGATTCTTCTTCAGGTTGGACAGCT | ||||||
| TIPC | N/A | GGGAATTTGTGTTTAAGAATATTGATGGTTAGCTGCTGTTTACAGTCCAAGATGGTAGTATTCTTGCT | N/A | 96 | N/A | [36] |
| AGTTACACTAGCCATCCTTACTGCGCTT | ||||||
| Molecular Beacons f | ||||||
| MBΔS3675/G3676/F3677 | ORF1a | HEX-CGCAGCCCTTTTAGCTTCAAACTAGTATCAACCATATCGGCTGCG-DABCYL | 11,267–11,306 | MN908947.3 | This study | |
| MBΔH69/V70 | S | HEX-GCCGTCCAGAGATAGCATGGAACCAAGACGGC-DABCYL | 21,750–21,777 | MN908947.3 | This study | |
| MBΔE156/F157 | S | HEX-CTCGGCGAATAAACTCCACTTTCCATCCAACGCCGAG-DABCYL | 22,014–22,044 | MN908947.3 | This study | |
| MBΔΝ211ins214EPE | S | HEX-CACCCCATCTTCTGGCTCACGCACTATGGGGTG-DABCYL | 22,193–22,207 | MN908947.3 | This study | |
| MBΔL242/A243/L244 | S | HEX-CGGCCTGGTCAAATAACTTCTATGTAAAGTTTGAAACCTAGCAGGCCG-DABCYL | 22,269–22,311 | MN908947.3 | This study | |
| MBIPC | N/A | FAM-GCCCACGTACCATCTTGGACTGTAAACAGCAGCCGTGGGC-DABCYL | N/A | N/A | [36] |
| Samples a | WHO VOC (Pango Lineage) d | Real-Time RT-PCR Result e | ||||
|---|---|---|---|---|---|---|
| ΔS3675/G3676/F3677 or ΔL3674/S3675/G3676 | ΔH69/V70 | ΔE156/F157 | ΔN211ins214EPE | ΔL242/A243/L244 | ||
| EVAg b | ||||||
| MT777677.1 | B.1 | − | − | − | − | − |
| BetaCoV/Germany/BavPat1/2020 p.1 | B.1 | − | − | − | − | − |
| EPI_ISL_918165 | Alpha (B.1.1.7) | + | + | − | − | − |
| EPI_ISL_1834082 | Beta (B.1.351) | + | − | − | − | + |
| EPI_ISL_877769 | Gamma (P.1) | + | − | − | − | − |
| EPI_ISL_2838050 | Delta (B.1.617.2) | − | − | + | − | − |
| Clinical Samplesc | ||||||
| BMV2539 | Delta (B.1.617.2) | − | − | + | − | − |
| BMV2543 | Delta (B.1.617.2) | − | − | + | − | − |
| BMV2542 | Delta (B.1.617.2) | − | − | + | − | − |
| BMV5679 | Omicron (BA.1) | + | + | − | + | − |
| BMV4354 | Omicron (BA.1.15) | + | + | − | + | − |
| BMV5890 | Omicron (BA.1) | + | + | − | + | − |
| BMV5872 | Omicron (BA.1.17) | + | + | − | + | − |
| BMV5687 | Omicron (BA.1) | + | + | − | + | − |
| BMV8083 | Omicron (BA.2) | + | − | − | − | − |
| BMV6824 | Omicron (BA.2) | + | − | − | − | − |
| BMV7517 | Omicron (BA.2) | + | − | − | − | − |
| BMV7015 | Omicron (BA.2) | + | − | − | − | − |
| BMV7067 | Omicron (BA.2) | + | − | − | − | − |
| BMV8468 | Omicron (BA.4) | + | + | − | − | − |
| BMV8444 | Omicron (BA.4) | + | + | − | − | − |
| BMV8436 | Omicron (BA.4) | + | + | − | − | − |
| BMV8054 | Omicron (BA.4) | + | + | − | − | − |
| BMV8072 | Omicron (BA.4) | + | + | − | − | − |
| BMV7629 | Omicron (BA.5.1) | + | + | − | − | − |
| BMV8220 | Omicron (BE.1.1) | + | + | − | − | − |
| BMV8173 | Omicron (BA.5.8) | + | + | − | − | − |
| BMV8056 | Omicron (BA.5.1) | + | + | − | − | − |
| BMV8177 | Omicron (BA.5.2) | + | + | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrysostomou, A.C.; Aristokleous, A.; Rodosthenous, J.H.; Christodoulou, C.; Stathi, G.; Kostrikis, L.G. Detection of Circulating SARS-CoV-2 Variants of Concern (VOCs) Using a Multiallelic Spectral Genotyping Assay. Life 2023, 13, 304. https://doi.org/10.3390/life13020304
Chrysostomou AC, Aristokleous A, Rodosthenous JH, Christodoulou C, Stathi G, Kostrikis LG. Detection of Circulating SARS-CoV-2 Variants of Concern (VOCs) Using a Multiallelic Spectral Genotyping Assay. Life. 2023; 13(2):304. https://doi.org/10.3390/life13020304
Chicago/Turabian StyleChrysostomou, Andreas C., Antonia Aristokleous, Johana Hezka Rodosthenous, Christina Christodoulou, Georgia Stathi, and Leondios G. Kostrikis. 2023. "Detection of Circulating SARS-CoV-2 Variants of Concern (VOCs) Using a Multiallelic Spectral Genotyping Assay" Life 13, no. 2: 304. https://doi.org/10.3390/life13020304