
Sugarcane Wax Metabolites and Their Toxicity to Silkworms

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Metabolome Analysis
2.2.2. Treatment of Feeding Experiment for Silkworm
16s rRNA Metabarcoding Analysis
3. Results
3.1. Sugarcane Wax Composition and the Major Metabolites
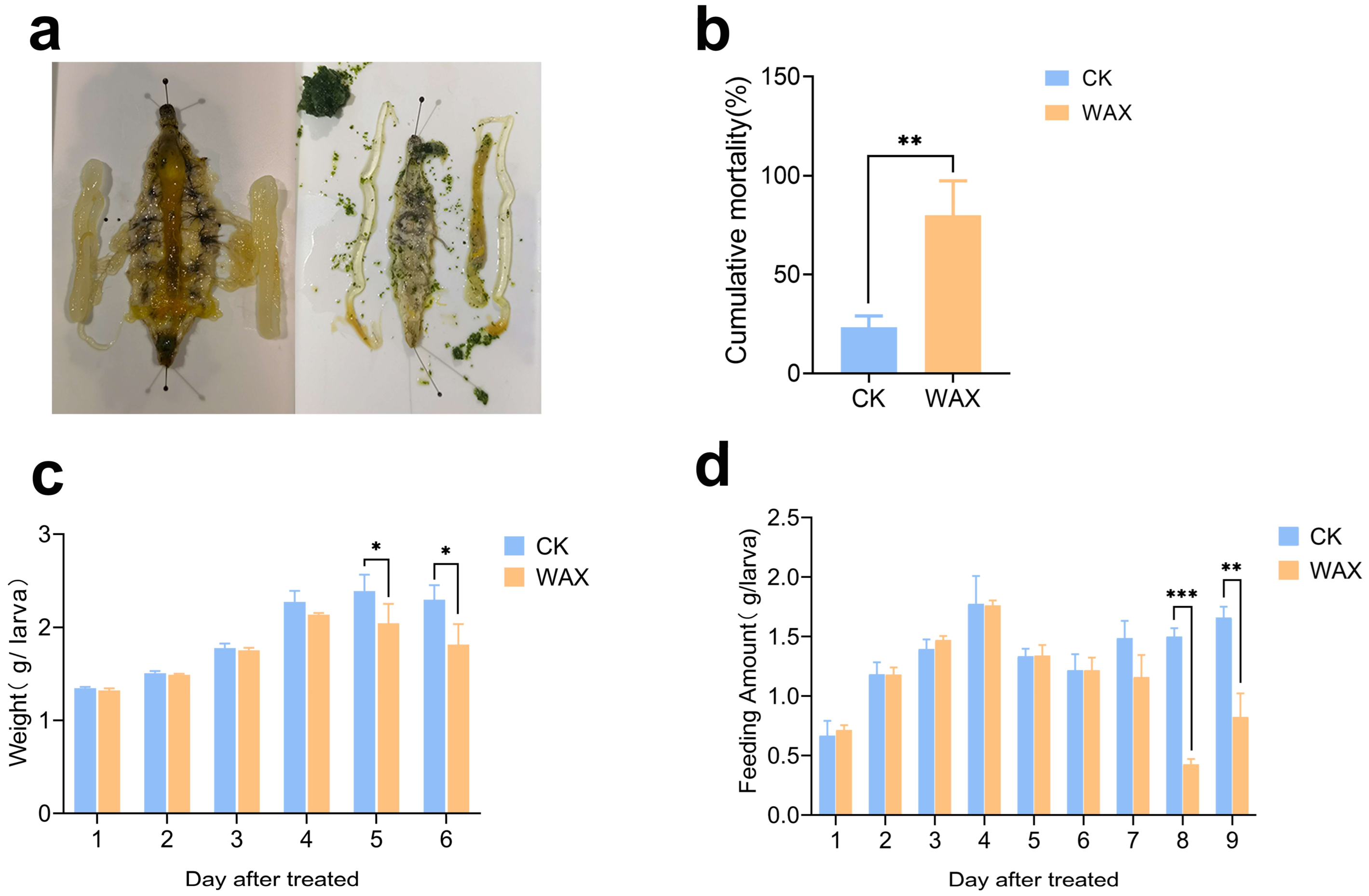
3.2. The Inhibited Appetite and Growth of the Silkworm Fed with Sugarcane Wax
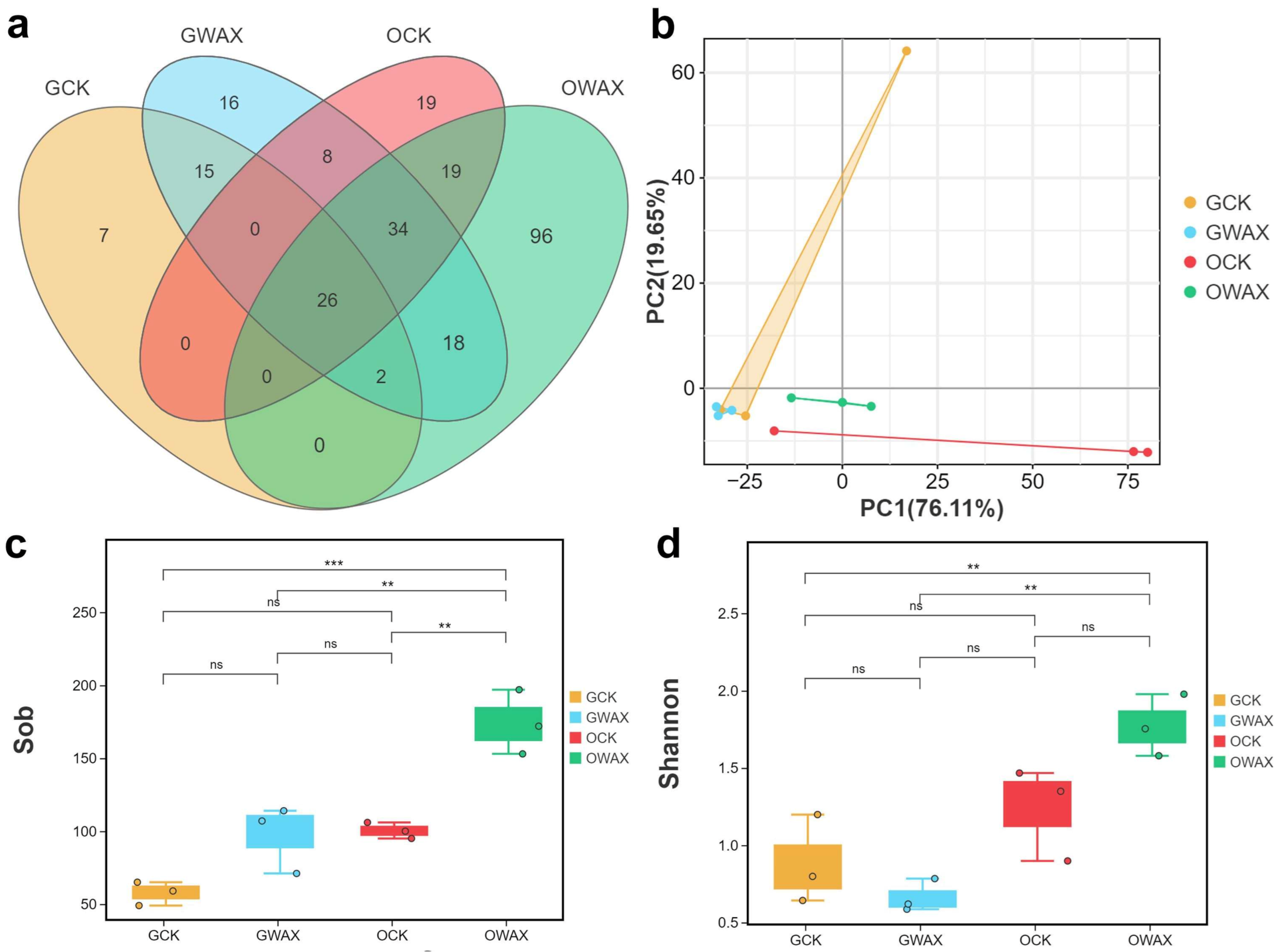
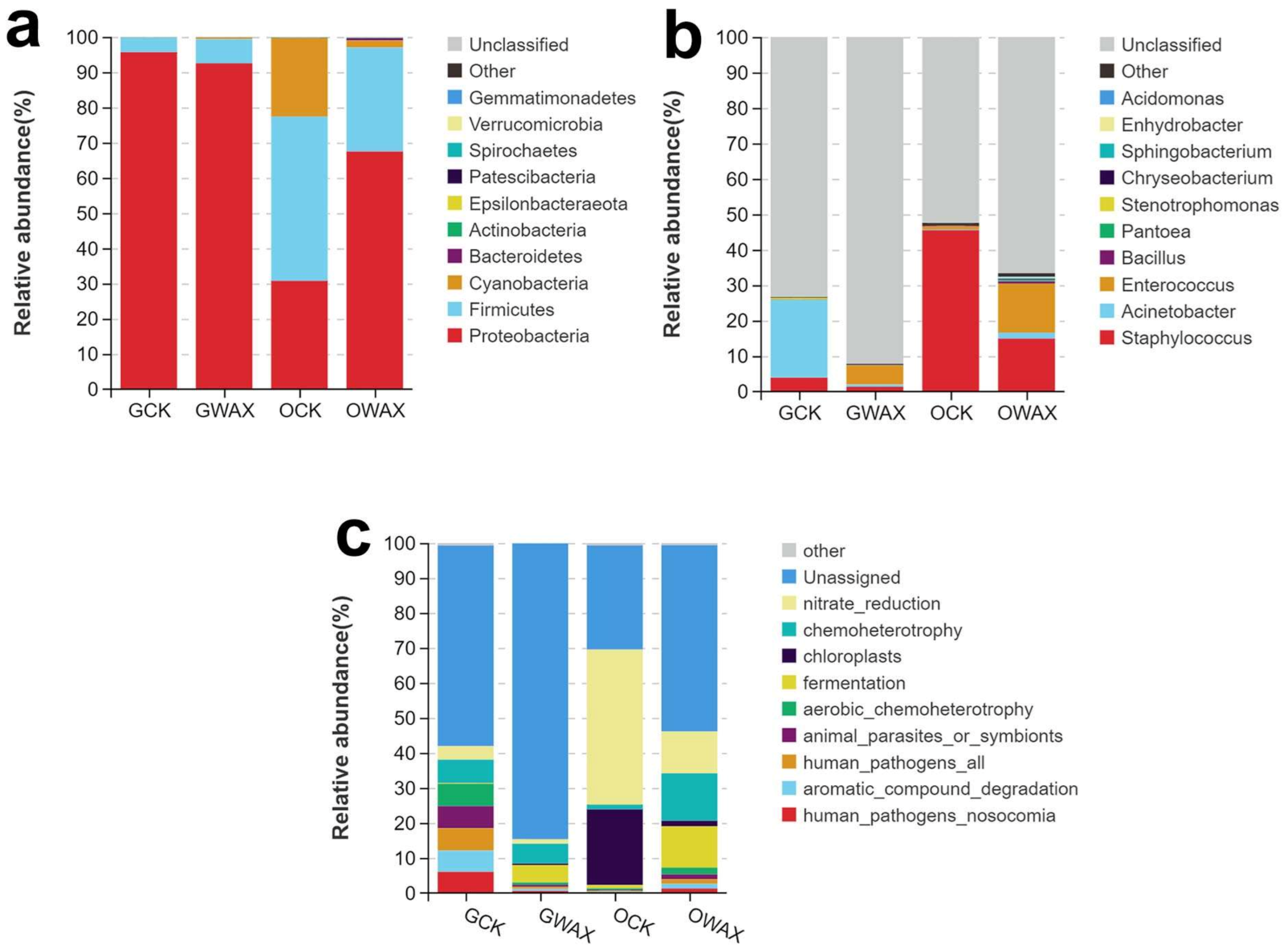
3.3. Bacterial Diversity Changed in the Gut of Silkworms Fed with Sugarcane Wax
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Oliveira Bordonal, R.; Carvalho, J.L.N.; Lal, R.; de Figueiredo, E.B.; de Oliveira, B.G.; La Scala, N. Sustainability of sugarcane production in Brazil. A review. Agron. Sustain. Dev. 2018, 38, 13. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Jackson, P.; Wei, X.; Ross, E.M.; Aitken, K.; Deomano, E.; Atkin, F.; Hayes, B.J.; Voss-Fels, K.P. Accelerating Genetic Gain in Sugarcane Breeding Using Genomic Selection. Agronomy 2020, 10, 585. [Google Scholar] [CrossRef] [Green Version]
- De Souza, A.P.; Grandis, A.; Leite, D.C.C.; Buckeridge, M.S. Sugarcane as a Bioenergy Source: History, Performance, and Perspectives for Second-Generation Bioethanol. BioEnergy Res. 2014, 7, 24–35. [Google Scholar] [CrossRef]
- Bhadha, J.H.; Xu, N.; Khatiwada, R.; Swanson, S.; LaBorde, C. Bagasse: A Potential Organic Soil Amendment Used in Sugarcane Production: SL477/SS690, 8/2020. EDIS 2020, 2020, 5. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Kheir, A.M.S. Saline soil properties, quality and productivity of wheat grown with bagasse ash and thiourea in different climatic zones. Chemosphere 2018, 193, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.J.; Duan, M.; Wang, J.; Han, S.; Ma, L.; Mo, X.; Li, M.; Hu, L.; Wang, L. Transcriptomic and Widely Targeted Metabolomic Approach Identified Diverse Group of Bioactive Compounds, Antiradical Activities, and Their Associated Genes in Six Sugarcane Varieties. Antioxidants 2022, 11, 1319. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Dermauw, W.; Mavridis, K.; Vontas, J. Significance and interpretation of molecular diagnostics for insecticide resistance management of agricultural pests. Curr. Opin. Insect Sci. 2020, 39, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Nuissier, G.; Bourgeois, P.; Grignon-Dubois, M.; Pardon, P.; Lescure, M.H. Composition of sugarcane waxes in rum factory wastes. Phytochemistry 2002, 61, 721–726. [Google Scholar] [CrossRef]
- Shanmugam, M.; Chithra, L.; Raman, N.; Venkatesan, M.S.; Ma, M. Sugarcane Wax—A Par Excellent by-Product of Sugar Industry—A Review. Agric. Rev. 2021, 42, 315–321. [Google Scholar] [CrossRef]
- Bezerra, J.I.M.; Molter, A.; Rafikov, M.; Frighetto, D.F. Biological control of the chaotic sugarcane borer-parasitoid agroecosystem. Ecol. Model. 2021, 450, 109564. [Google Scholar] [CrossRef]
- Rutherford, R.S. Mechanisms of Resistance to Pests and Pathogens in Sugarcane and Related Crop Species. In Sugarcane: Physiology, Biochemistry, and Functional Biology; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 435–482. [Google Scholar] [CrossRef]
- Rutherford, R.S.; van Staden, J. Towards a rapid near-infrared technique for prediction of resistance to sugarcane borerEldana saccharina walker (Lepidoptera: Pyralidae) using stalk surface wax. J. Chem. Ecol. 1996, 22, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Purcell, D.E.; O’Shea, M.G.; Kokot, S. Complex biopolymeric systems at stalk/epicuticular wax plant interfaces: A near infrared spectroscopy study of the sugarcane example. Biopolymers 2009, 91, 642–651. [Google Scholar] [CrossRef]
- Wartha, C.A.; de Aguiar Porto, N.; Tomaz, A.C.; Roque, J.V.; Diniz, M.B.T.; de Queiroz, M.E.L.R.; Teofilo, R.F.; Barbosa, M.H.P. Classification of sugarcane genotypes susceptible and resistant to the initial attack of sugarcane borer Diatraea saccharalis using epicuticular wax composition. Phytochemistry 2022, 199, 113175. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Syrenne, R.; Sun, J.-Z.; Yuan, J.S. Molecular approaches to study the insect gut symbiotic microbiota at the ‘omics’ age. Insect Sci. 2010, 17, 199–219. [Google Scholar] [CrossRef]
- Unban, K.; Klongklaew, A.; Kodchasee, P.; Pamueangmun, P.; Shetty, K.; Khanongnuch, C. Enterococci as Dominant Xylose Utilizing Lactic Acid Bacteria in Eri Silkworm Midgut and the Potential Use of Enterococcus hirae as Probiotic for Eri Culture. Insects 2022, 13, 136. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; He, J.; Zhang, N.; Muhammad, A.; Lu, X.; Shao, Y. Probiotic potentials of the silkworm gut symbiont Enterococcus casseliflavus ECB140, a promising L-tryptophan producer living inside the host. J. Appl. Microbiol. 2022, 133, 1620–1635. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Ishii, M.; Hasegawa, S.; Sekimizu, K. Enterococcus faecalis YM0831 suppresses sucrose-induced hyperglycemia in a silkworm model and in humans. Commun. Biol. 2019, 2, 157. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Sun, Z.; Cao, G.; Xue, R.; Hu, X.; Gong, C. Study of gut bacterial diversity of Bombyx mandarina and Bombyx mori through 16S rRNA gene sequencing. J. Asia-Pac. Entomol. 2019, 22, 522–530. [Google Scholar] [CrossRef]
- Dong, H.L.; Zhang, S.X.; Chen, Z.H.; Tao, H.; Li, X.; Qiu, J.F.; Cui, W.Z.; Sima, Y.H.; Cui, W.Z.; Xu, S.Q. Differences in gut microbiota between silkworms (Bombyx mori) reared on fresh mulberry (Morus alba var. multicaulis) leaves or an artificial diet. RSC Adv. 2018, 8, 26188–26200. [Google Scholar] [CrossRef]
- Chen, B.; Du, K.; Sun, C.; Vimalanathan, A.; Liang, X.; Li, Y.; Wang, B.; Lu, X.; Li, L.; Shao, Y. Gut bacterial and fungal communities of the domesticated silkworm (Bombyx mori) and wild mulberry-feeding relatives. ISME J. 2018, 12, 2252–2262. [Google Scholar] [CrossRef] [Green Version]
- Abdelli, N.; Peng, L.; Keping, C. Silkworm, Bombyx mori, as an alternative model organism in toxicological research. Environ. Sci. Pollut. Res. 2018, 25, 35048–35054. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.S.; Jordan, M.S.; Adams, S.M.; Suen, G.; Goodwin, L.A.; Davenport, K.W.; Currie, C.R.; Raffa, K.F. Cellulose-degrading bacteria associated with the invasive woodwasp Sirex noctilio. ISME J. 2011, 5, 1323–1331. [Google Scholar] [CrossRef]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senderovich, Y.; Halpern, M. The protective role of endogenous bacterial communities in chironomid egg masses and larvae. ISME J. 2013, 7, 2147–2158. [Google Scholar] [CrossRef] [Green Version]
- Mancini, M.V.; Damiani, C.; Accoti, A.; Tallarita, M.; Nunzi, E.; Cappelli, A.; Bozic, J.; Catanzani, R.; Rossi, P.; Valzano, M.; et al. Estimating bacteria diversity in different organs of nine species of mosquito by next generation sequencing. BMC Microbiol. 2018, 18, 126. [Google Scholar] [CrossRef] [Green Version]
- Duan, M.; Lu, M.; Lu, J.; Yang, W.; Li, B.; Ma, L.; Wang, L. Soil Chemical Properties, Metabolome, and Metabarcoding Give the New Insights into the Soil Transforming Process of Fairy Ring Fungi Leucocalocybe mongolica. J. Fungi 2022, 8, 680. [Google Scholar] [CrossRef]
- Guo, M.; Wu, F.; Hao, G.; Qi, Q.; Li, R.; Li, N.; Wei, L.; Chai, T. Bacillus subtilis Improves Immunity and Disease Resistance in Rabbits. Front. Immunol. 2017, 8, 354. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Inarkar, M.B.; Lele, S.S. Extraction and Characterization of Sugarcane Peel Wax. ISRN Agron. 2012, 2012, 340158. [Google Scholar] [CrossRef] [Green Version]
- Asikin, Y.; Takahashi, M.; Hirose, N.; Hou, D.-X.; Takara, K.; Wada, K. Wax, policosanol, and long-chain aldehydes of different sugarcane (Saccharum officinarum L.) cultivars. Eur. J. Lipid Sci. Technol. 2012, 114, 583–591. [Google Scholar] [CrossRef]
- Daisy, B.H.; Strobel, G.A.; Castillo, U.; Ezra, D.; Sears, J.; Weaver, D.K.; Runyon, J.B. Naphthalene, an insect repellent, is produced by Muscodor vitigenus, a novel endophytic fungus. Microbiology 2002, 148, 3737–3741. [Google Scholar] [CrossRef] [Green Version]
- Pajaro-Castro, N.; Caballero-Gallardo, K.; Olivero-Verbel, J. Toxicity of Naphthalene and Benzene on Tribollium castaneum Herbst. Int. J. Environ. Res. Public Health 2017, 14, 667. [Google Scholar] [CrossRef] [Green Version]
- Zong, S.; Liu, X.; Cao, C.; Luo, Y. Development of Semiochemical Attractants for Monitoring and Controlling Chlorophorus caragana. Z. Nat. C 2013, 68, 243–252. [Google Scholar] [CrossRef]
- Ingvarsdóttir, A.; Birkett, M.A.; Duce, I.; Genna, R.L.; Mordue, W.; Pickett, J.A.; Wadhams, L.J.; Mordue, A.J. Semiochemical strategies for sea louse control: Host location cues. Pest Manag. Sci. 2002, 58, 537–545. [Google Scholar] [CrossRef]
- Traas, P.C.; Boelens, H.; Takken, H.J. Two-step synthesis of a sex attractant of the male Boll Weevil from isophorone. Recl. Trav. Chim. Pays-Bas 1976, 95, 308–311. [Google Scholar] [CrossRef]
- Mamrutha, H.M.; Mogili, T.; Lakshmi, K.J.; Rama, N.; Kosma, D.; Kumar, M.U.; Jenks, M.A.; Nataraja, K.N. Leaf cuticular wax amount and crystal morphology regulate post-harvest water loss in mulberry (Morus species). Plant Physiol. Biochem. 2010, 48, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Yeruva, T.; Vankadara, S.; Ramasamy, S.; Lingaiah, K. Identification of Potential Probiotics in the Midgut of Mulberry Silkworm, Bombyx mori Through Metagenomic Approach. Probiotics Antimicrob. Proteins 2020, 12, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Lu, Y.; Zhang, H.; Kumar, D.; Liu, B.; Gong, Y.; Zhu, M.; Zhu, L.; Liang, Z.; Kuang, S.; et al. Effects of BmCPV Infection on Silkworm Bombyx mori Intestinal Bacteria. PLoS ONE 2016, 11, e0146313. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Compounds | Class | Formula | CAS | Peak Area |
|---|---|---|---|---|
| naphthalene | Aromatics | C10H8 | 91-20-3 | 3.90 × 106 |
| isophorone | Ketone | C9H14O | 78-59-1 | 3.03 × 106 |
| 2,4-nonadienal, (e,e)- | Aldehyde | C9H14O | 5910-87-2 | 1.70 × 106 |
| 6-methyl-3(2h)-pyridazinone | Heterocyclic compound | C5H6N2O | 13327-27-0 | 1.11 × 106 |
| hexanal | Aldehyde | C6H12O | 66-25-1 | 8.59 × 105 |
| 2-heptenal, (z)- | Aldehyde | C7H12O | 57266-86-1 | 8.20 × 105 |
| 3-methylbenzothiophene | Heterocyclic compound | C9H8S | 1455-18-1 | 7.78 × 105 |
| n-methylglycine | Acid | C3H7NO2 | 107-97-1 | 7.21 × 105 |
| cobalt, bis(.eta.-5-piperidinylcyclopentadienyl)- | Heterocyclic compound | C20H28CoN2 | 1000162-04-6 | 6.39 × 105 |
| benzene, pentamethyl- | Aromatics | C11H16 | 700-12-9 | 6.38 × 105 |
| benzeneacetaldehyde | Aldehyde | C8H8O | 122-78-1 | 6.37 × 105 |
| nonanal | Aldehyde | C9H18O | 124-19-6 | 5.69 × 105 |
| (5r,8ar)-5-propyloctahydroindolizine | Heterocyclic compound | C11H21N | 120057-35-4 | 4.90 × 105 |
| benzaldehyde | Aldehyde | C7H6O | 100-52-7 | 4.69 × 105 |
| 2,5-dihydroxy-4-isopropyl-2,4,6-cycloheptatrien-1-one | Ketone | C10H12O3 | 54755-56-5 | 4.34 × 105 |
| 2-acetonylcyclohexanone | Ketone | C9H14O2 | 6126-53-0 | 4.10 × 105 |
| nonanoic acid | Acid | C9H18O2 | 112-05-0 | 3.96 × 105 |
| benzene, 1,2,3,5-tetramethyl- | Aromatics | C10H14 | 527-53-7 | 3.87 × 105 |
| trans-4,5-epoxy-(e)-2-decenal | Aldehyde | C10H16O2 | 1000360-26-3 | 3.50 × 105 |
| naphthalene, 2-methyl- | Aromatics | C11H10 | 91-57-6 | 2.43 × 105 |
| SampleID | Tags | N90 (bp) | OTUs |
|---|---|---|---|
| GCK-1 | 116032 | 466 | 59 |
| GCK-2 | 126819 | 466 | 65 |
| GCK-3 | 120080 | 466 | 49 |
| GWAX-1 | 117556 | 466 | 107 |
| GWAX-2 | 127945 | 466 | 114 |
| GWAX-3 | 121674 | 466 | 71 |
| OCK-1 | 114870 | 443 | 106 |
| OCK-2 | 123914 | 443 | 95 |
| OCK-3 | 115539 | 466 | 100 |
| OWAX-1 | 98036 | 466 | 153 |
| OWAX-2 | 118634 | 466 | 197 |
| OWAX-3 | 123098 | 466 | 172 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Duan, M.; He, Z.; Zhang, Y.; Chen, Y.; Li, B.; Rao, M.J.; Hu, L.; Wang, L. Sugarcane Wax Metabolites and Their Toxicity to Silkworms. Life 2023, 13, 286. https://doi.org/10.3390/life13020286
Ma L, Duan M, He Z, Zhang Y, Chen Y, Li B, Rao MJ, Hu L, Wang L. Sugarcane Wax Metabolites and Their Toxicity to Silkworms. Life. 2023; 13(2):286. https://doi.org/10.3390/life13020286
Chicago/Turabian StyleMa, Li, Mingzheng Duan, Ziwei He, Yu Zhang, Yiting Chen, Bo Li, Muhammad Junaid Rao, Lihua Hu, and Lingqiang Wang. 2023. "Sugarcane Wax Metabolites and Their Toxicity to Silkworms" Life 13, no. 2: 286. https://doi.org/10.3390/life13020286